
Abstract
Alzheimer’s disease (AD) is one of the most common neurodegenerative diseases and is caused by accumulation of amyloid-β (Aβ) peptide and is associated with neurological abnormalities in learning and memory. The protective role of curcumin on nerve cells, along with a potent antioxidant and free radical scavenging activity, has been widely studied. However, its low bioavailability and limited transport ability across the blood-brain barrier are two major drawbacks of its application in the treatment of different neurodegenerative diseases. The present study was designed to improve the effectiveness of curcumin in the treatment of Aβ-induced cognitive deficiencies in a rat model of AD by loading it into nanostructured lipid carriers (NLCs). The accumulation rate of curcumin (505.76±38.4 ng/g–1 h) in rat brain, as well as its serum levels, were significantly increased by using curcumin-loaded NLCs. The effective role of NLCs for brain delivery of curcumin was confirmed by reduced oxidative stress parameters (ROS formation, lipid peroxidation, and ADP/ATP ratio) in the hippocampal tissue and improvement of spatial memory. Also, histopathological studies revealed the potential of Cur–NLCs in decreasing the hallmarks of Aβ in AD in the animal model. The result of studying the neuroprotective potential of Cur-NLC in both pre-treatment and treatment modes showed that loading curcumin in NLCs is an effective strategy for increasing curcumin delivery to the brain and reducing Aβ-induced neurological abnormalities and memory defects and that it can be the basis for further studies in the area of AD prevention and treatment.
Keywords
INTRODUCTION
Due to improvements in global health and life expectancy, the world’s older population will rise up to 22% by 2050. Dementia, a term that is used to describe the loss of a person’s cognitive functions and abilities is one of the major problems in aging. Approximately 47.4 million people are diagnosed with dementia worldwide, and this number is expected to triple by 2050 [1]. The most common form of dementia is Alzheimer’s disease (AD) [2]. The intracellular neurofibrillary tangles and the extracellular amyloid plaques are two most significant neuropathological AD symptoms [3]. Numerous studies have shown that inflammation and oxidative stress play important roles in the development of these lesions [4 –6]. AD is an advanced and complex neurological disorder that despite recent advances and medical science, has eluded a cure or an effective treatment [7]. The results from the intense studies carried out by many researchers in this area over the past years have led to the exploration of treatments and drugs that can relieve some of the patient’s symptoms and improve their performance, but unfortunately, none of them can stop the disease from progressing [7]. In addition, drug delivery to the brain is restricted, due to the presence of the blood-brain barrier and the tight junctions between endothelial cells [8]. As the number of people who are suffering from AD increases, the global demand for an effective solution to this problem increases as well. Various strategies for brain delivery of different drugs and medications have been evaluated, but this field still requires more research.
The use of nanoparticles is one of the successful employed strategies to overcome the BBB and increase the efficiency of drug delivery to the brain [9, 10]. Lipid-based nanoparticles are widely used by researchers in order to increase drug delivery to the brain, due to their unique advantages and quality [11]. This class of colloidal nanocarriers also has the advantages of both liposomal and polymeric nanoparticles; on the other hand, they do not have some restrictions on these carriers such as scale-up, cost of materials, production process, physical stability, and probability of toxicity. In particular, these lipid-based nanoparticles [solid lipid nanoparticles (SLNs) and nanostructured lipid carriers (NLCs)] are solid at room and body temperature and can control the drug release. Because of their lipidic nature, they are taken up readily by the brain [12, 13]. The role of the common surfactants in SLNs and NLCs production, such as Tween 80 and poloxamers, on the pattern of nanoparticles opsonization and protein corona is noticeable. The protein corona, due to the changes in the structural/conformational of proteins can determine the bio-fate of nanoparticles [14]. Apolipoprotein E (ApoE) is an example of this coating protein, since AopE receptors are predominantly expressed in the brain and binding ApoE with high affinity have pointed out the probability of this transporter mechanism across the BBB for SLNs and NLCs [15 –18]. Curcumin is the main active ingredient in peanut turmeric extract, with potent antioxidant, anti-inflammatory, and anti-Aβ accumulation effects [19, 20]. Beneficial effects of curcumin in different diseases, including neurogenic disorders like AD, has been reported previously [21, 22].
Curcumin alleviates Aβ-induced toxicity in cultured neurons cells [23], as well as protecting neuronal cells in a number of AD models by preventing the destruction of synapses [24]. Hoppe et al. showed a deleterious effect of Aβ on synapse activity in hippocampal fragments in the cell culture medium, as well as neutralizing these effects by curcumin [25]. On the other hand, curcumin plays a protective role against oxidative stress which is believed to be important in AD initiation and progression [26, 27]. Despite the mentioned beneficial effects, studies have shown that curcumin is not very stable under biological conditions, and due to the presence of hydroxyl groups in its structure, it is metabolized by the liver and kidney enzymes rapidly and shows poor absorption with low brain bioavailability [28, 29]. In addition, the BBB reduces its permeation to the brain even more [30].
In this study, curcumin loaded-NLCs (Cur–NLCs) are prepared and the uptake of curcumin by the brain and its biodistribution is investigated. Effects of the prepared formulation on disease progression and memory performance, as well as its effect on microscopic plaque formation in a rat model of AD, were also studied. It is worth mentioning that in most clinical trials, high amounts of curcumin (>1000 mg day-1) is used [31, 32]; however, by using the prepared nanocarrier system described in our present study, we were able to reduce the amount of curcumin dosage which can be considered as a great advantage.
MATERIALS AND METHODS
Curcumin (820354), cetyl palmitate (K44474819), and Tween®80 (8.22187) were purchased from Merck (Germany). Cholesterol (CD0122) was provided from Bio Basic (Canada). Amyloid-β Protein Fragment 1–42 (A9810) was purchased from Sigma (USA). A Milli-Q system (TKA, Smartt pure, and Germany) was used to prepare deionized water. Other chemical reagents were of analytical grade. Sprague-Dawley (SD) rats (200–220 g) 6–8 weeks old were obtained from the Centre of Laboratory Animals from Tehran University of Medical Sciences. All animal studies were conducted in compliance with the rules of the Ethics Committee of Tehran University of Medical Sciences.
Cur–NLCs preparation
Cur–NLCs were prepared by homogenization method [33]. Briefly, a mixture of cetyl palmitate, oleic acid, and cholesterol (3 : 1:1 weight ratio) was heated to 80±5°C while stirring. Then curcumin (drug to lipid ratio = 0.1 w/w) was dispersed into the melted lipid phase. The aqueous solution containing Tween 80 (0.7% w/v) was added to the lipid phase in a dropwise fashion and mixed by magnetic stirring (1000 rpm) for 30 min and then homogenized (10000 rpm) for 8 min (Heidolph, Germany). In order to obtain curcumin loaded–NLCs, the resultant lipidic dispersion was cooled down to room temperature.
In vitro characterizations of curcumin-loaded NLCs
An extensive in vitro optimization and characterization properties of Cur-NLCs prepared in this study has been reported in detail in our previous work [34]. Briefly, particle size and polydispersity index of the prepared nanodispersion was measured by photon correlation spectroscopy technique using a dynamic light scattering instrument (Malvern Zetasizer, UK). Samples were diluted with deionized water to achieve appropriate concentrations and measurements were done at 25°C, viscosity 0.8872 cP, and water refractive index of 1.33 under 90° at 633 nm wavelength. A scanning electron microscope (SEM, HITACHI S-4160, Netherlands) was used to determine the morphology of optimized nanoparticles. Thermal analysis of Cur-NLCs, and bulk materials was performed by differential scanning calorimetry (DSC, Mettler Toledo, UK). Thermograms of samples were recorded at a heating rate of 10°C min–1 under nitrogen gas at a temperature range of 30 to 250°C. The crystallinity of curcumin, cetyl palmitate, and dried Cur–NLCs were characterized using powder X-ray diffractometer (XRD, X’Pert PRO MPD, Netherlands) with a copper anode (Generator Settings: 40 mA, 40 kV). The samples were scanned and analyzed from 2θ (degree) of 3–80° with the step size of (°2θ): 0.0260 and the scan step time of (s): 37.9950. In vitro release studies of Cur–NLCs compared to free curcumin (unloaded curcumin) were investigated using the dialysis bag method [35, 36].
Pharmacokinetic and bio-distribution studies
Pharmacokinetic studies were done on male Sprague-Dawley rats (200–220 g at 6–8 weeks old, n = 30). The animals were kept in separate cages (54×33×20 cm) and maintained on a 12 h light/12 h dark cycle in a temperature-controlled environment (23±2°C) with free access to food and water. One day prior to the experiment, a polyethylene-silicone rubber cannula was inserted in the rat’s right jugular vein using a standard surgical protocol [37 –39]. The control sample (curcumin solution) was prepared by dissolving curcumin in dimethylacetamide, PEG400 and 5% dextrose solution (15 : 45 : 40 volume ratio), as reported in previous studies [40, 41]. On the day of the experiment, rats were divided into two groups (n = 15 for each group) which either received curcumin or Cur–NLCs (4 mg/kg–1 Cur). After drug administration, a 500 μl blood sample was obtained (from 3 rats each time) at the time intervals of 15, 30, 60, 120, and 240 min. The animals were anesthetized and decapitated immediately following each blood sampling and their brain and liver were collected, washed externally by saline, weighed and homogenized with an equal volume of saline to obtain homogenates. The serum plasma was also separated by centrifuging blood samples at 3000×g for 10 min. To determine the amount of curcumin, 200 μl acetonitrile was added to the samples with a ratio of 2 : 1, centrifuged (10000×g, 10 min) and supernatants were analyzed by HPLC (Knauer, EZ Chrom Elite) using a fluorescence detector (RF-10A×L) and a C18 column (Nucleodur®, 250 mm×4.6 mm, Germany) while the column temperature was kept at 25°C. A combination of citric acid buffer and acetonitrile (pH = 4) was used as the mobile phase with 50 μl injection volume and a flow rate of 1.4 ml/min–1. The detection wavelengths were set at 410 nm (λex) and 533 nm (λem). The detection limit of the assay was 10 ng/ml–1. The mean recovery rate of curcumin in plasma and brain was nearly 80% in the range of 10–1000 ng/ml–1.
Pharmacokinetic parameters were determined by non-compartmental analysis approach based on the area under the drug concentration versus time curve (AUC 0–t) according to standard procedures. To avoid confusion when determining the brain concentration of curcumin, the concentration of curcumin in brain blood samples were deducted from the total concentration of curcumin in the brain of the rat, while considering the brain blood volume (Vvas = 0.11 ml/g–1) in all our calculations according to previous studies [42].
Evaluation of neuroprotective effects of curcumin loaded-NLCs
The animal model of AD
Although the exact mechanism for the development of neurotoxicity caused by Aβ formation is not yet understood, existing evidence supports the major role of oxidative stress in this regard [4]. The hippocampus is a part of the brain with an important role in memory while Aβ accumulation initiates the process of neuronal destruction and apoptosis in this area of the brain [43]. Aβ was injected into hippocampus of the rat in order to create the AD model. The Aβ injections to the hippocampus induce neurodegeneration and reduce learning and memory [44, 45] which is accompanied by a decrease in synaptic transmission and neuronal dysfunction. Free radicals can initiate oxidative processes followed by neural degeneration and nerve damage [46].
Aβ aggregation preparation
Lyophilized Aβ was dissolved in PBS 0.1 mole lit–1 with a concentration of 200 ng μl–1 which was then aliquoted and stored at –20°C. Aβ solution was incubated for 7 days in 37°C, and was diluted to a final concentration of 100 ng μl–1 on the day of experiment [47].
Induction of AD in rats
Pharmacodynamics studies were conducted in 5 groups of male Sprague-Dawley rats (n = 30 for each group). The animals were kept in separate cages (54×33×20 cm) and maintained on a 12 h light/12 h dark cycle in a temperature-controlled environment (23±2°C) with free access to food and water. The Ethics Committee Guidelines of Tehran University of Medical Science was followed while working with the animals. On the day of the experiment, rats were anesthetized by i.p. injection of ketamine-xylazine, their hair was shaved and the scalp was dis-infected with alcohol 70% and the head was fixed in the stereotaxic device (Stoelting, Wood Dale, IL, USA). A longitudinal cut was made to the posterior part of the head and the skull became exposed. The skin was separated from the skull and 25% aluminum solution was used to stop bleeding. After determining the stereotaxic coordinates according to the Paxinos and Watson atlas [48] and conducting pilot stereotaxic coordinate studies for the lateral cerebellar ventricles as well, a double-hole drill was made into the skull. A Hamilton syringe was used to inject 2 μl of Aβ suspension (100 ng μl–1) on each side of the posterior hippocampus (the injection duration was 1 min) with the following coordinates: depth: –2.8, lateral: ±2 ‘anteroposterior: –3.5. Finally, the scalp was sutured. For prevention of infection, 40 mg of cefazolin was injected intraperitoneally for 4 days. The sham group was treated in the same way; however, instead of Aβ suspensions, they received an equal volume of saline solution. Morris water maze trials were performed from the 17th day of administration.
Drug administration and experimental design
The AD animal models were randomly divided into 5 groups (Table 1): Aβ (AD control), sham, BNLCs, CNLCs + Aβ, and Aβ+ CNLCs groups (for each group, n = 7). A schematic diagram of drug treatment schedule and protocol is demonstrated in (Fig. 3A).
Experimental groups and treatments
CNLCs: Cur-NLCs.
Brain homogenate preparation
Following the Morris water maze test, rats were anesthetized and their brain hippocampus tissue was extracted and kept at –80°C until the biochemical studies were carried out. On the day of the experiment, brain samples were thawed at 4°C, then homogenized in cold saline. The homogenates were centrifuged at 1000×g for 10 min and the supernatant was used for further biochemical studies.
Determination of malondialdehyde (MDA)
The MDA as an index of lipid peroxidation and level of oxidative stress was investigated using the method described by Zhang et al. [49]. 0.25 ml sulfuric acid (0.05 mole lit–1) and 0.3 ml thiobarbituric acid (0.2%) were added to 0.2 ml brain homogenate solution to perform the MDA reaction with thiobarbituric acid at 95–100°C. After 2 h of incubation, extraction was carried out using n-butanol. Then, the samples were centrifuged (3500×g) for 10 min. Formation of a pink complex was determined by spectrophotometer (PerkinElmer, lambda 365, USA) at 532 nm. The numerical value was reported in terms of μmol of MDA/mg of protein.
Reactive oxygen species (ROS) assessment
To determine the amount of ROS, 2, 7-dichlorofluorescein diacetate (DCF), an oxidation-sensitive fluorescent dye, was used. First, a buffer containing 5 moles of DCFH-DA in 1.25 mmol/lit–1 of methanol was prepared and incubated with brain homogenate solutions at 37°C for 30 min in the dark. DCFH-DA became attached to intracellular esterases and subsequently converted to DCFH and was then oxidized by ROS and converted to a DCF molecule with a strong fluorescence signal. The fluorescence was measured by ELISA F-2000 fluorescence spectrometer at 485 nm (λex) and 525 nm (λem).
Measurement of ADP/ATP ratios
In order to determine the ADP/ATP ratio, the hippocampus samples were thawed at 4°C, homogenized by adding 1 ml of 6% trichloroacetic acid and centrifuged at 4°C at 12000×g for 10 min. The supernatant was then neutralized using potassium hydroxide solution (4 mol/lit–1). An HPLC system (Waters, Milford, MA) was used for determining the ADP/ATP ratios, an ultraviolet-visible detector (Waters 486, USA) and a C18 column (SUPELCOSIL, 250 mm×4.6 mm, 5 μm), while the column temperature was kept at 25°C. A solution containing potassium phosphate buffer (0.1 M, pH 5.5), tetra-butyl ammonium hydrogen sulfate (4 mmole/lit–1) and methanol (85 : 15 v/v) was used as the mobile phase with 50 μl injection volume and flow rate of 1 ml/min–1. The detection wavelength was set at 254 nm. The levels of ATP and ADP were measured by the standard curve and then the ratio was evaluated [50].
Measurement of hippocampal total thiol groups
In a 10 ml test tube, the hippocampus homogenates were mixed with 0.6 ml of Tris base (0.25 mole/lit–1) and EDTA (20 mmole/lit–1) (pH 8.2). Then 40 ml of DTNB solution (10 mmole/lit–1 in methanol) was added to each test tube. The mixtures were diluted in a volume of 4 ml with methanol and centrifuged at 3000×g at room temperature for 15 min. The color was developed over 15–20 min. The absorbance of the supernatant was read at 412 nm using a spectrophotometer after 20 min [51].
Histopathology
Histopathological studies were performed on 3 rats in each group. Rat brain tissue was extracted, washed with saline, and kept in formalin until the day of the experiments. After performing the common tissue preparation processes, paraffin blocks were prepared and cut into 5 μm sections. Three sections were prepared for each brain sample. The prepared sections stained with Congo red and cresyl violet (Nissl staining). The histological slides were evaluated by an independent reviewer, using light microscopy (Olympus BX51; Olympus, Tokyo, Japan) to observe possible changes following Aβ injection due to inflammatory changes, degeneration of the neurons or formation of amyloid plaques. Magnification×400 was employed for counting the Nissl-positive cells and the calculation was repeated for three fields in each section, using computer software Image-Pro Plus®V.6 (Media Cybernetics, Inc., Silver Spring, USA). Finally, the average number of Nissl-positive cells in these fields was then recorded. The histological studies were done using an optical microscope.
The Morris water maze experiment
The Morris water maze method was used to determine if Aβ injection induced an AD phenotype in rats and to study the possible protective effect of curcumin on learning and memory and space learning [52]. One circular black pool (136 cm in diameter and 35 cm in depth) containing water with a temperature of 25±2°C was prepared and divided into four quadrants. The appropriate visual cues were mounted around the pool for the rats in order to be able to recognize their position. A Plexiglas black platform with a diameter of 10 cm in the northwest quadrant of the pool (goal quadrant) was immersed 2 cm below the water surface. The platform position remained constant throughout the training days. The procedure for the behavior assessment was done during 4-day training session and a probe trial test. The training included 4 sessions for 4 days, with each session consisting of 4 trials. In each trial test, the animals were placed at the starting point and allowed to swim for 90 s to find the platform. If the animal could not manage to find the platform within 90 s, it would be guided to the platform by hand. The rats were given 20 s to rest on the platform. This was done for each of the four starting points and the swimming direction of the rat was recorded with a camera mounted on top of the pool. In training sessions, the learning process was evaluated through the following three indicators: swimming speed, escape latency (the time that is taken to find the platform) and the traveled distance (the track distance traveled before finding the platform). The probe test was performed 24 h after the training session. In the probe test, the platform was removed and rats were allowed to swim for 90 s, then the period of time that each animal has spent in the target quadrant was recorded and analyzed. The behavioral data were recorded by using a computerized video tracking system and analyzed using Ethovision XT software (Version 6.0 Noldus Information Technology, Leesburg, VA). In order to assess the rat’s vision ability to follow the cues, a visual test was performed on all rats as well. A visible platform covered in foil, placed 1 cm higher than the water surface in the opposite quadrant. The rats were placed from a certain starting point in the pool and the time to reach the platform was recorded. All the trials were carried out at the same time of day and the environment conditions were kept consistent.
Statistical analysis
Quantitative data are reported as Mean±Standard Deviation of triplicate experiments. All measurements were analyzed statistically using ANOVA, independent t-test and paired t-test using SPSS (Version 16, SPSS Chicago, IL). For multiple comparisons, one-way ANOVAs were followed by Dunnett post hoc analysis when appropriate. Independent t-test and paired t-test were used for comparison between two groups. In all measurements, a p value < 0.05 considered significant.
RESULTS
Cur–NLCs in vitro characteristics
Optimized Cur–NLCs had a uniform spherical distribution when evaluated with SEM with the mean particle size of 117.36±1.36 nm and the polydispersity index of 0.188±0.020 (Fig. 1B). Crystalline curcumin endothermic peak during 173°C was disappeared in the Cur–NLCs thermogram due to changes in the physical state of curcumin from crystal to amorphous (Fig. 1C). In addition, a slight shift in cetyl palmitate peak (from 56 to 58°C) was probably due to the incorporation of curcumin into the lipid matrix or reduced particle size and increased surface area that led to a decrease in the melting temperature compared to the bulk material [53]. However, the characteristic curcumin peaks at 2°θ of 5 to 30 could not be detected in the XRD spectra of Cur–NLCs which indicated that curcumin was solubilized within the lipid matrix (Fig. 1D). In addition, the reduction of cetyl palpitate XRD spectral intensity in Cur–NLCs may be attributed to the intercalation of curcumin between the crystal lattice of the lipid, leading to a change in crystallinity which supports lower crystallinity of the lipid matrix in Cur–NLCs compared to bulk solid lipid (Fig. 1D). The in vitro release profiles of Cur–NLCs in Tween 80 phosphate buffer solution (1%, pH = 7.4) in sink conditions are shown in (Fig. 1E). During the study period, an initial burst curcumin release was observed that has also been reported previously in a number of studies on lipid-based nanoparticles [13 , 55]. However, at the second step, a steady and sustained release was observed. The results showed that approximately 65% of curcumin was released from NLCs over the test period whereas more than 80% of free curcumin was released in the early hours of the study.

The schematic diagram of Cur-NLC preparation method (A). The scanning electron microscope images demonstrated that the Cur-NLCs were uniform and spherical in shape (B). Crystalline curcumin demonstrated a sharp endothermic peak at 173°C corresponds to its melting point (Tm), which disappeared in the Cur-NLCs DSC thermogram (C). Free curcumin due to its crystalline structure has the characteristic peaks at 2θ (degree) of 5 to 30 by X-ray diffraction analysis. However, these peaks could not be detected in the XRD spectra of Cur-NLCs, which indicated that curcumin was solubilized within the lipid matrix (D). Up to 80% of free curcumin were released in the initial hours, over the test period. However, loaded curcumin in NLCs has a controlled slow rate of release (E).
Pharmacokinetic and biodistribution studies
To study the brain uptake of curcumin and the effect of using NLCs in maximizing BBB permeability, the pharmacokinetic parameters of the plasma, brain, and liver following the i.v. injection of curcumin solution and Cur–NLCs were evaluated (Fig. 2). It was observed that the area under the curve (AUC0–t), as well as curcumin’s half-life in plasma, was increased in Cur–NLCs (Fig. 2A). Increasing the MRT of curcumin in the plasma for Cur–NLCs indicated that the half-life of curcumin was increased (Table 2). This can be the result of decreased curcumin clearance due to its loading in NLCs. In addition, the brain accumulation of curcumin could not be detected at any of the sampling times. The brain concentration-time curve of curcumin initially showed an increasing trend, reaching its maximum in 60 min in Cur-NLCs. Curcumin concentration was detectable in the brain until the end of the sampling (Fig. 2B). Studying accumulation of curcumin in the liver at different times showed that curcumin concentrations in early time points (until 2 h) was lower in rats receiving Cur–NLCs compared to those who received Cur solution (Table 2 and Fig. 2B), which confirms lower reticuloendothelial system (RES) uptake of Cur-NLCs [56]. The AUC0–t of curcumin in brain (505.76±38.47 ng/g–1 h) was measured 2 h after dosing that produced 513±90.79 ng/g–1 h in the plasma [57] suggesting which by using low doses of curcumin (4 mg/kg–1), achievable curcumin brain levels may be close to curcumin blood levels.

Time-courses of plasma (A), brain (B), and liver (C) concentrations (ng/ml–1) of curcumin in male rats following a 4 mg/kg–1 i.v dose of curcumin solution and curcumin loaded by NLCs. The AUC0–t of curcumin in plasma has increased in Cur-NLCs (A). The brain accumulation of curcumin solution could not be detected at any of the sampling times. In which that the brain AUC0–t of curcumin initially showed an increasing trend reaching a maximum in 1 h in Cur-NLCs (B). Curcumin concentration was measurable in the brain until the end of the sampling. AUC0–t of liver showed that curcumin concentrations in early time points (until 1 h) were significantly lower in the case of rats receiving Cur-NLCs compared to those receiving curcumin solution, which confirmed the less RES uptake of Cur-NLCs. Statistical analysis was performed using an (ANOVA) test. Error bars represent standard deviation. Data are represented as mean±sd of 3 rats. ** p < 0.01 compared to rats who received Cur solution.
The plasma, brain and liver pharmacokinetic parameters of curcumin after i.v. administration of curcumin solution and Cur-NLC at a dose of 4 mg/kg in rat. aMean±S.D., n = 3
AUC: The area under the concentration – time curve, MRT: The mean residence time.
Neuroprotective effects of Cur–NLCs on AD animal model
Since high brain uptake of curcumin (505.76± 38.4 ng/g–1 h) was observed in Cur–NLCs, compared to curcumin solution (0.00±0.0 ng/g–1 h), the pharmacodynamics studies were designed to evaluate possible neuroprotective effects of curcumin in Aβ-induced AD by loading it into NLCs. In order to elucidate the mechanism of curcumin’s effect in an AD models, oxidative stress tests and histological examinations of the hippocampus were compared and the results were analyzed.
The effects of curcumin on the lipid peroxidation level
Effects of curcumin on the lipid peroxidation process were studied on the brain tissue samples of rats. The MDA levels in the hippocampus of rats pretreated (CNLCs + Aβ) and treated (Aβ+ CNLCs) with Cur-NLC are illustrated in (Fig. 3B). Analysis of variance showed that the MDA content in Aβ (AD control) hippocampus was significantly increased compared to sham group [58], while the MDA levels were significantly (p < 0.01) decreased in the treatment group. Previous studies have shown that curcumin not only acts as a direct radical scavenger but it also possesses powerful antioxidant properties and is able to significantly reduce lipid peroxidation [59, 60]. Our results also showed that Cur–NLCs effectively reduced the damage caused by oxidative stress in the treatment group. These results are in accordance with those of earlier reports that curcumin has protective effects against lipid peroxidation and decreases MDA [60].

A schematic diagram of drug treatment schedule and protocol design (A). The MDA content in Aβ (AD control) hippocampus was significantly increased compared to sham group, while the MDA levels significantly decreased in Aβ+ CNLCs (treatment) group (B). Injection of the Aβ into the hippocampus site caused a significant increase the production of ROS in AD control group compared to the sham group. In both of CNLCs + Aβ (prevention) and Aβ+ CNLCs (treatment) groups, a significant decrease in ROS production was observed, in comparison with the Aβ control group (C). The Cur-NLCs administration noticeably increased thiol level in comparison with the AD control group (p < 0.001). Moreover, this increased level of thiol in prevention group was more than in the treatment group (D), and the ADP/ATP brain ratio decreased in the groups receiving curcumin; however, this reduction was significant in the prevention group. Data are represented as mean±sd of 3 rats. * p < 0.1, ** p < 0.01, and *** p < 0.001compared to Aβ (AD control).
The significant effect of curcumin on ROS levels
The injection of the Aβ into the hippocampus site caused a significant increase in ROS production of AD control group (Fig. 3C) compared to the sham group (p < 0.05). The results showed a significant decrease of ROS in Cur-NLCs administration groups compared to the AD control group (p < 0.05) [61].
Curcumin reduced mitochondrial dysfunction
Curcumin has shown activities against mitochondrial dysfunction suggesting that there is a close relationship between the antioxidant properties of curcumin and the mitochondrial function [62]. Our results showed a significant difference in ADP/ATP level between the AD control groups compared to the sham group (p < 0.001). However, the ADP/ATP brain ratio showed a significant decrease in the prevention (p < 0.001) and treatment group (p < 0.003) compared to the AD control group (Fig. 3E).
Thiol groups in the hippocampus
Cur-NLCs administration significantly increased thiol levels in comparison with the AD control group (p < 0.001) (Fig. 3D). Moreover, this increase in thiol level in prevention group was significantly higher than the treatment group (p < 0.05).
Histopathological examination
Effects of Aβ administration on survival of hippocampal neural cells was examined using Cresyl violet staining of brain tissue sections (Fig. 4A–D). Morphological signs of necrosis were evident in the AD control group including cytoplasmic swelling of neurons, various degrees of vacuolization, and damaged cell bodies (Fig. 4B). In addition, the organization of this cellular layer (hippocampus CA1 neuronal layer) was completely disrupted. The evaluation of blood vessels in the brain hippocampus tissues sections showed amyloid angiopathy in AD control group (Fig. 4B, arrowheads). Moreover, all Congo red-stained sections from different experimental groups were evaluated histologically for any deposition of Aβ in the hippocampus tissue (Fig. 4E–H). The micrograph of saline-stereotaxic injected group is illustrated in Fig. 4E. No significant histopathological changes were observed in this group. The evaluation of AD control group has shown numerous Aβ deposits (plaque) in hippocampus area (Fig. 4F). Histopathological examination of the prevention and treatment groups showed that the number of Aβ plaques were decreased compared to the AD group (Fig. 4G and 4H, respectively); however, necrosis of CA1 pyramidal neurons and vacuolization were also evident in these groups (Fig. 4C, D). There was no Aβ deposition in the prevention group (Fig. 4G).

Identification of neuronal survival by Nissl-staining in rat hippocampi in the CA1 region in sham-, AD-, pre-treatment-, and treatment- groups, respectively (A–D). Representative images of Congo red staining of hippocampal region from AD model in sham, AD, pre-treatment-, and treatment- groups, respectively (E–H). The AD control group has shown morphological signs of necrosis (B). There are no significant histopathological changes in Congo red-stained sections of sham group. The evaluation of AD control group has shown numerous Aβ deposits in hippocampus area (F). Histopathological examination of the prevention and treatment groups showed that the numbers of Aβ plaques were decreased in comparison to the AD control group (G, H); however, necrosis of CA1 pyramidal neurons and vacuolization were also evident in this groups (C, D). Thick arrows: Aβ plaques, thin arrows: CA1 pyramidal neurons necrosis. Arrow heads: Aβ deposition in brain vessels.
Behavioral tests
In order to monitor the brain function improvement and learning process during training session, traveled distance, escape latency, and swimming speed were investigated. To examine spatial memory during the probe test day, the time that each animal had spent in the target quadrant was also studied. This parameter demonstrates the degree of learning and memory that has taken place after the training session. The time spent and distance travelled before finding the escape platform in four trials of each group are shown in (Fig. 5A and 5B), respectively. There was no significant difference between the first and fourth days in AD control group indicating memory loss and a delay in learning time. In sham group, there was significant decrease in both parameters (p < 0.002) that represents progressive learning ability following the training session [52]. The results showed that escape latency was significantly decreased between the first and fourth days in both prevention and treatment groups (p < 0.002). As shown in (Fig. 5B), Cur-NLCs treatment significantly decreased distance travelled to find the escape platform for both the treatment (p < 0.0012) and pre-treatment (p < 0.006) groups. Figure 5D represents the comparison of the swimming speed between the AD control, sham, and two Cur–NLCs receiving groups. There was no significant difference in the swimming speed between these 4 groups, which confirms the absence of any motor disturbances in animals [52]. Comparison of the time spent in the target quadrant on the probe test is represented in (Fig. 5C). The time spent in the target quadrant relative to the AD control group showed a significant increase in both the prevention (p < 0.003) and treatment groups (p < 0.001). The results of the probe test after removing the platform indicate that Cur-NLCs-receiving rats had greater retention of the position of the than did the AD control group. There was no significant difference in the time spent in target the quadrant between prevention and treatment groups. The result of visual test showed no significant difference in escape latency and traveled distance to find the visible platform, which corroborated the visual capability of animals in perceiving the visual cues. The results of this study showed that intracerebral injections of Aβ disrupted learning ability and spatial memory, as well as increasing the oxidative stress, compared to the control group [45, 50]. In several previous studies on Aβ, it was reported that the injection process alone did not affect the brain at all, but the observed damages that have occurred are caused by Aβ accumulation. Aβ aggregates cause cell death and initiate inflammatory processes through different mechanisms [63].
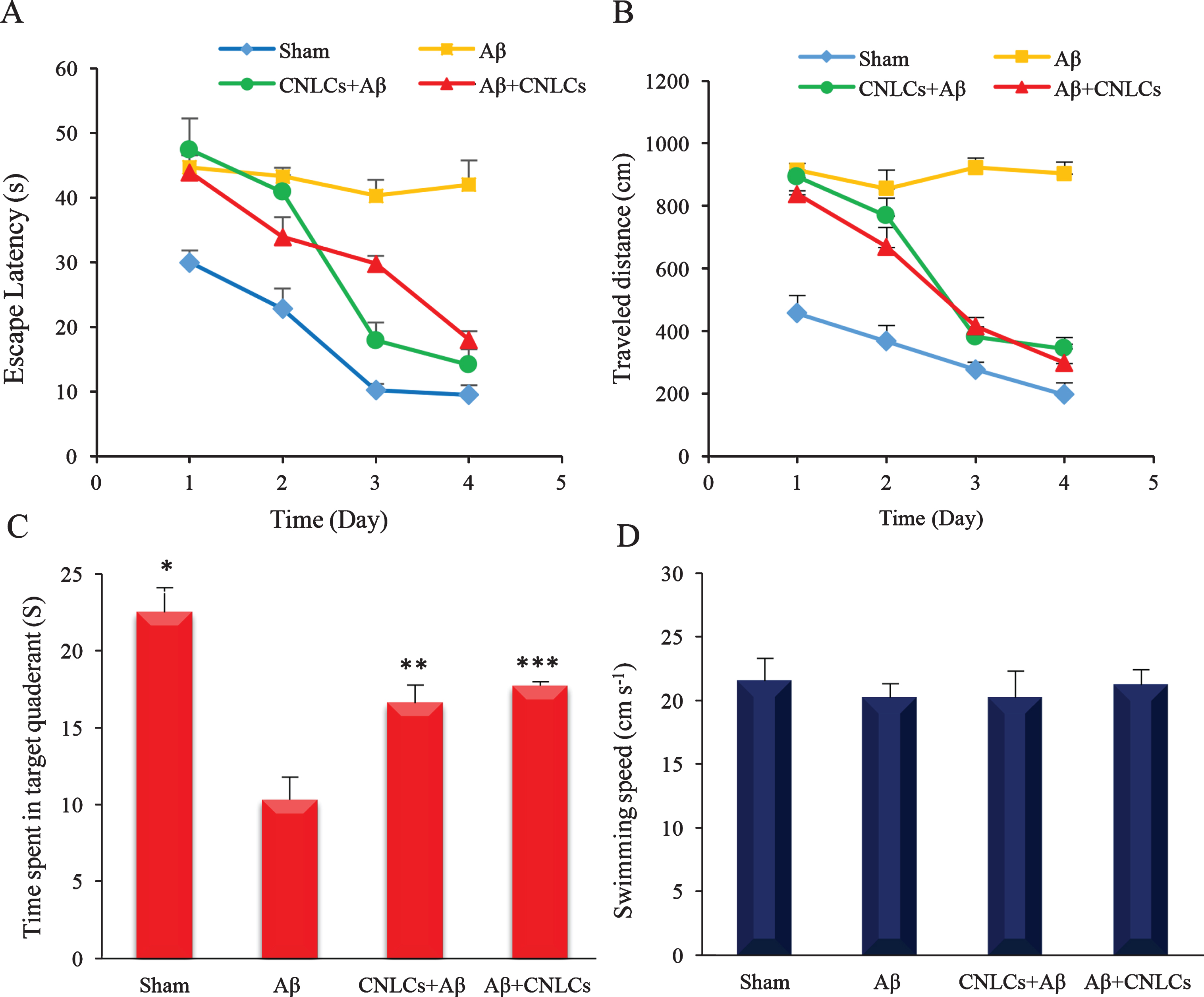
The effect of Cur-NLCs on (A) escape latency to platform zone and (B) distance travelled in MWM during the four trials of each training session in sham, Aβ (AD control), CNLCs + Aβ (prevention), and Aβ+ CNLCs (treatment) group. There was no significant difference between the first and fourth days in AD control group (A, B). In sham group there was significant decrease in both parameters (p < 0.001) that represent significant learning of task. The result of treated groups with Cur-NLCs showed a significant decrease in the escape latency and traveled distance compared to AD control group (A, B). Comparison of the time spent in the target quadrant on the probe test showed a significant increase in both Cur-NLCs receiving groups (C). There was no significant difference in the swimming speed between 4 groups, which indicated no motor disturbances in animals (D). Data are represented as mean±sd of 6 rats. * p < 0.1, ** p < 0.01, and *** p < 0.001compared to Aβ (AD control).
DISCUSSION
The therapeutic efficacy of curcumin for AD is prevented because of low brain penetration and bioavailability. In this study, we incorporated curcumin in a colloidal drug carrier system, nanostructured lipid carrier (NLCs). NLCs are suitable choices for targeted brain delivery because of good penetration from BBB and absorption by the brain due to their lipidic nature [64]. The results of pharmacokinetic studies indicate that, however, the brain accumulation of curcumin could not be detected in any sampling times but curcumin concentration was detectable in the brain until the end of the sampling (Fig. 2B). The longer retention time of curcumin could be due to the loading of curcumin in NLCs, as a reserve source for sustained release of curcumin in brain tissue. The results from the release studies confirm this controlled release. The accumulation of curcumin in the brain may be due to the nano sizes of the particles (about 120 nm) and the presence of Tween 80 on NLCs surface which may cause the uptake reduction of lipid carriers by RES [56]. In addition, the adsorption of circulating plasma ApoE on nanoparticles surface can improve uptake in to the brain through targeting LDL receptors on BBB endothelial cells [65, 66].
Many studies have pointed out the probability of this transport mechanism across the BBB for lipid-based nanoparticles [13 , 67]. However, many studies have also reported that Tween 80 can inhibit the brain’s P-gp and prevent drug efflux from the brain [68]. The nanoparticles size is one of the important parameters affecting the opsonization process and the nature of protein corona [69, 70]. In fact, the protein corona determines the bio-fate of nanoparticles. ApoE is an example of these coating proteins. Crossing the BBB also follows the same principle [15]. This phenomenon can cause the carriers to be less detectable and decrease their elimination by RES; however, more research needs to be done in order to determine the exact mechanism involved.
Increasing MDA, as a known product of nerve damage in oxidative stress conditions and free radical formation, is an important indicator of lipid peroxidation. Additionally, hippocampal MDA levels are often used as oxidative markers for AD models. Previous studies have shown that not only does curcumin act as a direct radical scavenger, but it also possesses powerful antioxidant properties and is able to significantly reduce lipid peroxidation [59]. Our results also showed that Cur–NLCs effectively reduced the damage caused by oxidative stress in the treatment group. These results are in accordance with those of earlier reports that curcumin has protective effects against lipid peroxidation and decreases MDA [60]. Oxidative stress has been introduced as not only an inducer, but as a sustaining factor in neurological diseases, including AD. Brain cells are highly affected by oxidative stress since they contain high contents of oxygen and unsaturated fatty acids that may cause nerve cell structural and functional damages. The results showed that using Cur-NLCs in both modes (prevention and treatment) can reduce ROS levels compared to the AD control group (p < 0.05). Our previous study has also shown that the preparation process of Cur-NLCs did not have any significant effect on the antioxidant activity of curcumin [34]. The antioxidant activity of curcumin, by inhibiting the formation and propagation of free radicals, has been reported previously [61]. Cur-NLCs treatment reduced mitochondrial dysfunction in AD animal model. Accordingly, a decrease in MDA level was observed accompanied by an increase in total thiol group levels in the hippocampus tissue especially in the prevention group. Protein-bound thiols and non-protein thiol groups are the major players of the defense system against oxidative stress [71]. Decreased Aβ aggregation in the presence of curcumin in histopathological examination of the prevention and treatment groups could be the result of NFκB inhibition and creation of an anti-inflammatory medium. Inhibition of NFκB leads to decreased formation of cytochemokines and Aβ [71]. Considerable reductions of Aβ deposition in the prevention group (Fig. 4F) can be attributed to the anti-Aβ accumulation effects of curcumin and inhibiting the assembly of Aβ plaques. Several studies have shown that curcumin can prevent Aβ aggregation and increase the dissociation of Aβ aggregates [72]. The highest improvements in terms of Aβ plaque deposition and number of surviving CA1 pyramidal neurons in the pretreatment group, suggests that Cur-NLCs can be used as an effective therapeutic agent after AD progression and development of neurological lesions. These results confirmed that the observed decrease in MDA and ROS levels as well as the increasing levels of thiol group concentrations in brain tissue samples of Cur-NLC-treated rats was accompanied with learning and memory improvements. The Morris water maze method was used to evaluate spatial learning and behavioral tests in different groups (AD control, sham, and two Cur–NLCs receiving groups). Decreased escape latency in Morris water maze in trials confirms the role of Cur-NLCs pretreatment in memory improvement. Cur-NLC administration decreased the time to reach the hidden platform in Morris water maze. The results of the probe test after removing the platform shows that Cur-NLCs-receiving rats had a higher ability in finding and remembering the platform’s position and spent more time in the target quadrant compared to the AD control group. In our study, the comparison of sham and AD groups was in accordance with the previous reports. Also, in all parts of our study (behavioral and biochemical tests), no significant difference was observed between the AD control group and the group which received blank NLCs (data not shown). This evidence confirmed that Cur–NLCs administration improved learning ability and spatial memory in rats with AD and was also able to reduce the levels of oxidative stress induced by Aβ in these animals. However, the lipid carrier alone did not show any effect on these parameters.
Conclusion
In this study, curcumin-loaded NLCs were prepared by using a modified homogenization protocol and the efficacy of the narrow size distribution (<120 nm) and high entrapment efficiency were investigated to improve neuroprotective effects of curcumin on an animal model of AD. Administration of Cur-NLCs in AD animal resulted in noticeable improvements in learning and memory capabilities in rats. The results of this study revealed that Cur-NLCs pre-treatment of AD rats might be able to reduce deficits caused by Aβ injection via providing a proper environment to maintain the functionality of neurons. Moreover, treatment of AD rats with Cur-NLCs improved the disease symptoms through reducing the number of amyloid plaques. Our results suggest that curcumin may even be beneficial after disease development. Due to low bioavailability and poor brain penetration of curcumin, its loading in the NLCs could increase curcumin accumulation, even at low doses. However, further studies are needed to optimize this treatment strategy.
Footnotes
ACKNOWLEDGMENTS
This study was financially supported by Tehran University of Medical Sciences (TUMS; Grant No: 93-04-33-27656), Zanjan University of Medical Sciences (ZUMS), and Iran Nanotechnology Initiative Council (INIC). The authors would like to especially thank Mehdi Gholami for his kind help in the animal testing part of the study and Parnian Banikarimi for her help with proofreading.