
Abstract
Circadian rhythm dysfunction is present in Alzheimer’s disease. Animal models of Alzheimer’s disease have been employed to investigate whether this dysfunction is a risk factor or symptom of the disease. The circadian phenotype in mouse models of Alzheimer’s disease is very disparate in terms of the degree and timing of the dysfunction. This is likely a result of some models elevating amyloid-β protein precursor instead of just the amyloid-β fragment present in human Alzheimer’s disease. We characterized activity rhythms in a novel knock-in mouse model (APPNL-G-F) of Alzheimer’s disease that elevates amyloid-β without overexpressing amyloid-β protein precursor. Despite increased rhythm amplitude, total activity, and a shortening of free-running period at 15 months of age, all other aspects of the activity rhythm were similar to controls from three to fifteen months of age. At two months of age, these mice were also able to entrain to a light-dark cycle with a period right on the edge of entrainment, which further suggests a healthy functioning circadian system. These data open the possibility that circadian rhythm disruptions in transgenic models of Alzheimer’s disease could be a result of these models having an artificial phenotype caused by overexpression of amyloid-β protein precursor.
INTRODUCTION
Circadian rhythms are endogenously generated cyclical processes that take approximately 24 hours per cycle. Sleep-wake cycle, locomotor activity, temperature regulation, water/food intake, metabolism, and hormone secretion are all physiological measurements of circadian rhythms. In humans and animal models, circadian rhythm disruption has been associated with many disease states and cognitive impairments [1, 2]. For example, there is circadian rhythm dysfunction in Alzheimer’s disease (AD). AD patients often sleep more during the day and less during the night-time (for review, see [3]). In fact, severity of dementia has been correlated with amount of circadian rhythm dysfunction (for review, see [3]).
While circadian rhythm disruption clearly accompanies AD, it is debated as to whether it is a cause or symptom of the disease. An intriguing hypothesis is that the circadian rhythm disruption is contributing to the cognitive impairments in AD [3–6]. The recent findings that circadian rhythm disruption in cognitively normal adults, as well as polymorphisms in the Clock gene increase the chances of developing AD, suggests that circadian rhythm disruption might precipitate AD development [7–11]. Similarly, the finding that environmental circadian rhythm disruption exacerbates memory in a pre-symptomatic mouse model of AD also suggests that circadian rhythm disruption might be a risk factor of AD [12].
Animal models of AD allow circadian rhythms and cognitive decline to be studied at a mechanistic level. Transgenic models of AD are useful because the hallmarks of AD pathology can be mimicked in a graded manner that mirrors human AD disease progression. Activity rhythms have been assessed in a variety of different transgenic mouse AD models. Several strains such as APP23, TgCRND8, and 3xTg display changes in diurnal activity patterns, with some of the changes occurring before AD pathology [3, 13–16]. There are also changes in free-running period in some strains, such as 3xTg and Tg2576 [16, 17].
Despite the strengths of transgenic mouse models of AD, they also possess faults. First, all AD mouse strains are not created equally, and this is seen in circadian rhythm applications. Despite the circadian rhythm changes mentioned above in mouse models of AD, often the circadian changes are very minor, as in APPxPS1 mice [18]. APPswe/PS1dE9 mice are a prime example with largely intact rhythms up to seven months of age [19]. Another fitting example of this is that the lengthening of period in Tg2576 mice is only seen in constant darkness and not in a light/dark cycle [17].
Perhaps more importantly, another problem is that transgenic AD mouse models are not representative of human AD pathology as they involve artificial phenotypes [20–24]. Thus, it is possible that circadian rhythm degradation in aging and AD might be mediated by a pathological mechanism completely independent from changes in amyloid-β (Aβ). This is a potential problem with AD mouse models that typically focus on one etiological mechanism.
There is a new AD mouse model that utilizes a novel knock in approach to elevate Aβ, which differs from the transgenic models that elevate Aβ by overexpressing amyloid-β protein precursor (AβPP) [20–22]. In this APPNL-G-F model, only Aβ42 is elevated and not all Aβ subtypes as in transgenic models. Aβ progression and deposition resembles that of humans and is in marked contrast to that of transgenic mice. APPNL-G-F mice develop cortical and hippocampal amyloid plaques at two and four months, respectively [20, 25]. These mice also have increased activity in the open field, deficits in behavioral flexibility, attention, and spatial learning around six months of age [20, 26]. However, in the Morris water task, the findings are mixed with some studies failing to find impairments at six [27] and 10-11 [28] months of age. We recently reported Morris water task impairments at six, nine, and 12 months of age [25, 26]. It is likely that differences in Morris water task protocol, and or facilitated learning and memory from repeated behavioral testing in the same animals are contributing to the disparate findings [26].
Much has yet to be discovered with these animals as there has not been any circadian rhythm characterization of any kind. In the present study, we assessed activity rhythms in APPNL-G-F mice. In one group of animals, activity rhythms were assessed continuously from three months of age to 15 months of age. Some manipulations such as stress can accelerate AD pathogenesis in APPNL-G-F mice [29]. With this in mind, in another group of animals, we exposed them to a non-24 hour light dark cycle at two months of age in order to potentially exacerbate the circadian phenotype. Similar to Levault and colleagues [12], we were assessing if environmental circadian rhythm disruption before the onset of AD symptomology could speed up pathogenesis. Despite this strain of AD mouse being very different from the previously studied transgenic models, we anticipated there to be dysfunction in activity rhythms.
MATERIALS AND METHODS
Subjects and materials
All experimental procedures were approved by the University of Lethbridge Animal Care Committee in compliance with the standards set out by the Canadian Council for Animal Care. The male and female breeding pairs of AD knock in mice carrying Swedish (NL), Arctic (G), and Beyreuther/Iberian (F) mutations were provided by RIKEN Brain Science Institute, Japan [20]. This strain has been established as a C57BL/6J congenic line [20]. Then, a colony of these mice was maintained at the Canadian Center for Behavioural Neuroscience in the University of Lethbridge. The strain was bred as heterozygote (APPNL-G-F+/NL-G-F-) × heterozygote (APP NL-G-F+/NL-G-F). Genotyping of all mice was done with polymerase chain reaction on ear notch tissue. For the AD animals, 20 male mice homozygous for the knock-in (APPNL-G-F+/NL-G-F+) were used and as in Saito and colleagues [20], eight wild type littermates (APP+/+) mice served as control animals.
The mice were assigned to groups based on their age and genotype. The AD mice were divided into two groups depending on their age. The first AD group (AD; n = 8) and the control group (Con; n = 8) was put into wheels at approximately three months of age (average age of 96 days old; range of 80–110 days old) and remained there for 12 months. These mice were maintained on a 12:12 hour light dark cycle with lights on at 0900 and lights off at 21:00. The other group of AD animals was put into wheels at approximately two months of age (average age of 57 days; range of 49–64 days of age) and remained there for eight months. An average age of two months was desirable for this group (Shift; n = 12) because they were going to be exposed to a non-24-hour light dark cycle to see if this accelerated circadian rhythm degradation. These mice were maintained on a 12:12 hour light dark cycle with lights on at 0730 and lights off at 1930.
Initially all of the mice were housed in groups of four mice per cage, until being placed in wheels, in which they were singly housed. The mice were randomly assigned to wheels. While housed in groups and singly in the wheels, enrichment was provided in the form of crinkle paper and cotton Nestlets. At all times, the mice were maintained in a controlled environment with a temperature of 22 to 25°C, and a humidity of 50%. The mice had free access to food (Purina standard laboratory chow; Woodstock, Ontario) and water for the duration of the experiment. The mice were checked daily by animal care staff to ensure they were of good health. The nails of the mice were periodically trimmed when needed to minimize the effects of scratching.
The AD and Con mice were housed in polycarbonate shoe-box cages that were 35 cm ×20.5 cm with a wheel of a diameter of 11 cm. The Shift mice were housed in polycarbonate shoe-box cages that were 45.5 cm×26 cm with a wheel diameter of 16.5 cm. On the outside of the cage, magnetic sensors were attached near a magnet affixed to the axle of the wheel. Clocklab data collection software (Coulbourn Instruments, Allentown, PA, USA) was used to collect data. The animals were checked daily at random times during the light phase of the cycle, to determine that they were healthy and had enough food and water. The cages were cleaned every two weeks at random times during the light phase of the cycle. The Clocklab data collection software was turned off for all animals at the beginning of cage cleaning and was turned on again when completed for all animals, which was typically approximately 1.5 h.
Procedures
Refer to Fig. 1 for flow charts of the experimental procedures for each group of animals. Day 1 refers to the day that the animals were added to wheels. Seven (AD & Con mice) and two months (Shift mice) into the experiment respectively, the mice received water task training during the light phase of the cycle. The training lasted nine days (distributed paradigm) for the AD & Con mice and three days (massed paradigm) for the Shift mice. The behavioral experiment was incomplete and the data were beyond the scope of the present study and thus were not included. Instead, readers are encouraged to consult our recent extensive characterization of the behavioral and brain changes associated with this AD model [26].

Experimental timeline for all groups of animals. The numbers refer to the day that an experimental event occurred in relation to the day that the animals were added to wheels. DD refers to conditions of constant darkness.
The AD and Con mice received two nine-day exposure of lights off (D12:D12; DD) at nine months into the experiment (∼12 months of age) and 12 months into the experiment (∼15 months of age). Also, just before the last DD exposure, these mice were exposed to a four-hour phase advance of the light dark cycle. On this day, lights on was at 0500 and lights off was 1700 instead of 0900 and 2100, respectively.
The Shift mice received a T21 light dark cycle (12L:9D; see Table 1) for six days, which was equivalent to a three-hour phase advance each day, at approximately 67 days of age. For the AD, Con, and Shift groups, the primary experimental outcome was behavioral changes in circadian rhythms either over time (AD and Con groups) or after T21 exposure (Shift group).
T21 schedule for the Shift group. T21 exposure commenced on day 12 and ended on day 17
Data analysis
The data were analyzed in two-week blocks, with a total of 24 blocks. The First nine days of DD occurred in between block 20 (ended day 281) and block 21 (started day 297). Block 24 ended on day 352. The four-hour phase advance occurred on day 360 and for analysis the five days before the shift were compared to the five days after the shift. Finally, the last DD period occurred from day 365 to day 373.
Clocklab analysis software was used to conduct all circadian analyses. Period refers to the time it takes to complete one oscillation and is perhaps the most important indicator of circadian rhythm quality. A chi squared periodogram was used to determine the period with one-minute bins tested between 20 and 26 hours at a significance of p = 0.001. Amplitude of the chi squared periodogram was used as a measure of rhythm prominence. Because in rodents most of the activity occurs in the dark, the onset of activity in relation to the start of the dark period can be used to measure the phase of circadian rhythms. Phase speaks to entrainment, so it is possible for phase to be alerted without the period of the oscillation to be affected. Clocklab activity onsets were calculated and then manually adjusted if needed so that the phase of activity onset in relation to lights off could be calculated. A positive number indicates that activity started after lights off. Much like period, the length of the active state can also be used to determine the state of the central circadian pacemaker. Alpha was calculated by subtracting the activity offsets from the activity onsets. The total number of wheel revolutions, which similar to amplitude of the chi squared periodogram, speaks to the strength of the rhythm, was calculated for each day and per each phase of the light dark cycle. Nocturnality, refers to how much activity is occurring during the dark phase of the light dark cycle and is another way to assess if the rhythm is entrained to the light dark cycle. It was then calculated by dividing the number of revolutions during the dark phase of the cycle divided by the total number of revolutions per day. Lastly, activity bouts can be used to determine if the activity is fragmented. Fragmentation is indicative of circadian rhythm degradation and is a hallmark of aging. Activity bouts had a threshold of six revolutions per minute and minimum interval between bouts of 20 min [30]. All of the above measures were analyzed for the AD and Con groups via mixed model ANOVAs with block as the repeated measures factor and group as the within subjects factor.
For the Shift group, the dominant period of the chi square was compared for the 10 days before, six days during, and 10 days after T21 exposure. Amplitude of the chi squared and nocturnality, were also analyzed this way. Repeated measures ANOVAs were used to compare these circadian behaviors before, during, and after the T21. As the time of lights on and off was continuously changing during the T21, phase was only analyzed before and after T21 exposure with a paired samples T-test. Also, to determine if the T21 had any long-lasting effects on behavior the Shift group was compared to the AD and Con groups at six months of age and nine months of age. For this analysis 14 days were selected at six and nine months of age for the Shift group and were then compared to the blocks in which the AD and Con mice were six and nine months of age, respectively. One-way ANOVAs were used to compare performance among the groups at six and nine months of age.
SPSS was used to conduct all statistical analyses SPSS 21 (IBM, Armonk, New York) and Prism (GraphPad, La Jolla, CA) was used to make all of the figures. Two-tailed tests with a critical p value of 0.05 were used in all instances. In the case of significant interactions for the mixed model ANOVAs or significant one-way ANOVAs, Bonferroni post-hoc comparisons were conducted. Standard error of the mean is used in all figures.
RESULTS
Long-term assessment of circadian rhythms
Period did not change over time, nor was it different between the groups (Figs. 2 and 3A). Nocturnality increased over time (Fig. 3C; F(23, 322) = 1.782, p = 0.016, η2p = 0.113), and although not different between the groups, there was a significant block×group interaction (F(23, 322) = 1.699, p = 0.025, η2p = 0.108). Follow up post-hoc tests revealed that in block 23 the AD mice had a significantly greater nocturnality index (p = 0.010). Although phase became significantly delayed over time (Fig. 2D; F(23, 322) = 5.455, p < 0.001, η2p = 0.280), there was no difference between the groups. Amplitude (Figs. 2 and 3B; F(23, 322) = 53.251, p < 0.001, η2p = 0.792) and total number of wheel revolutions (Fig. 4A; F(23, 322) = 48.140, p < 0.001, η2p = 0.775) decreased with time, and were higher in AD mice (amplitude: F(1, 14) = 5.107, p = 0.040, η2p = 0.267; wheel revolutions: F(1, 14) = 4.871, p = 0.045, η2p = 0.258). Alpha increased over time (Fig. 4B; F(23, 322) = 23.008, p < 0.001, η2p = 0.622) and despite the difference between the groups only approaching significance (F(1, 14) = 4.244, p = 0.058, η2p = 0.233), there was a significant interaction (F(23, 322) = 2.280, p = 0.001, η2p = 0.140). Follow up post-hoc tests indicated that the controls had a greater alpha in blocks 5 (p = 0.011), 7 (p = 0.047), 13 (p < 0.001), and 18 (p = 0.023), whereas the AD group had a greater alpha in blocks 6 (p = 0.018), and 11 (p = 0.001). Bouts increased over time (Fig. 4C; F(23, 322) = 30.139, p < 0.001, η2p = 0.683), but there were no differences between the groups.
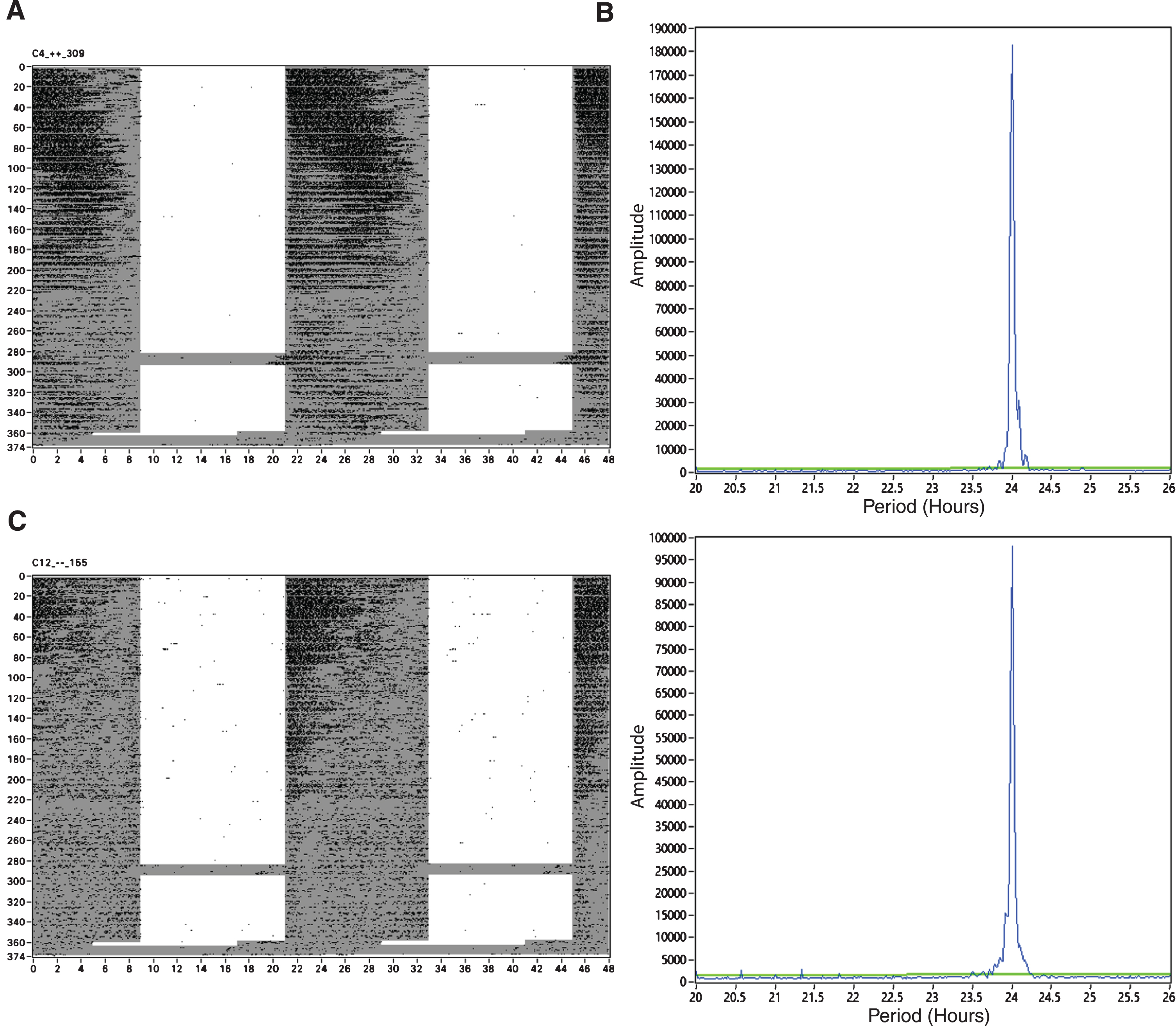
Longitudinal depiction of activity rhythms in APPNL-G-F mice. A) A representative actogram from an APPNL-G-F/NL-G-F mouse (n = 8). B) A representative χ2 periodogram from an APPNL-G-F mouse. C) A representative actogram from a control mouse (n = 8). D) A representative χ2 periodogram from an App-/- mouse. Figures represent behavior from approximately three months of age to 15 months of age. Days since placed in wheels are represented on the Y axis of the actograms. The grey shaded regions of the actograms indicate lights off.
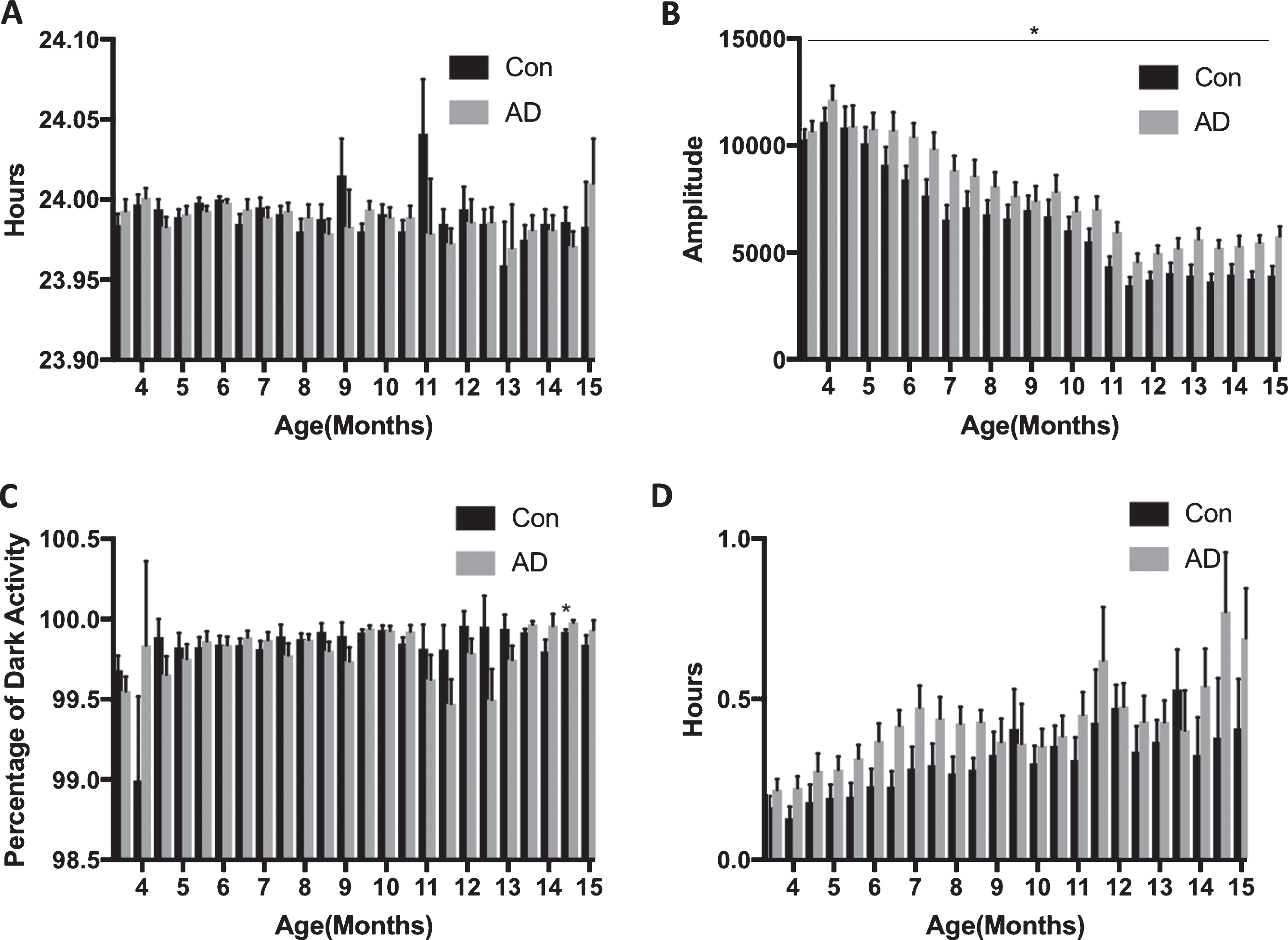
Activity rhythms are largely intact in APPNL-G-F mice up to 15 months of age. A) There were no differences in period between AD (n = 8) and Con mice (n = 8). B) Amplitude was greater for the AD mice (p = 0.040). C) Nocturnality (percentage of dark activity) was not different between the groups. D). The onset of activity in relation to lights off (phase) was not different between the groups.
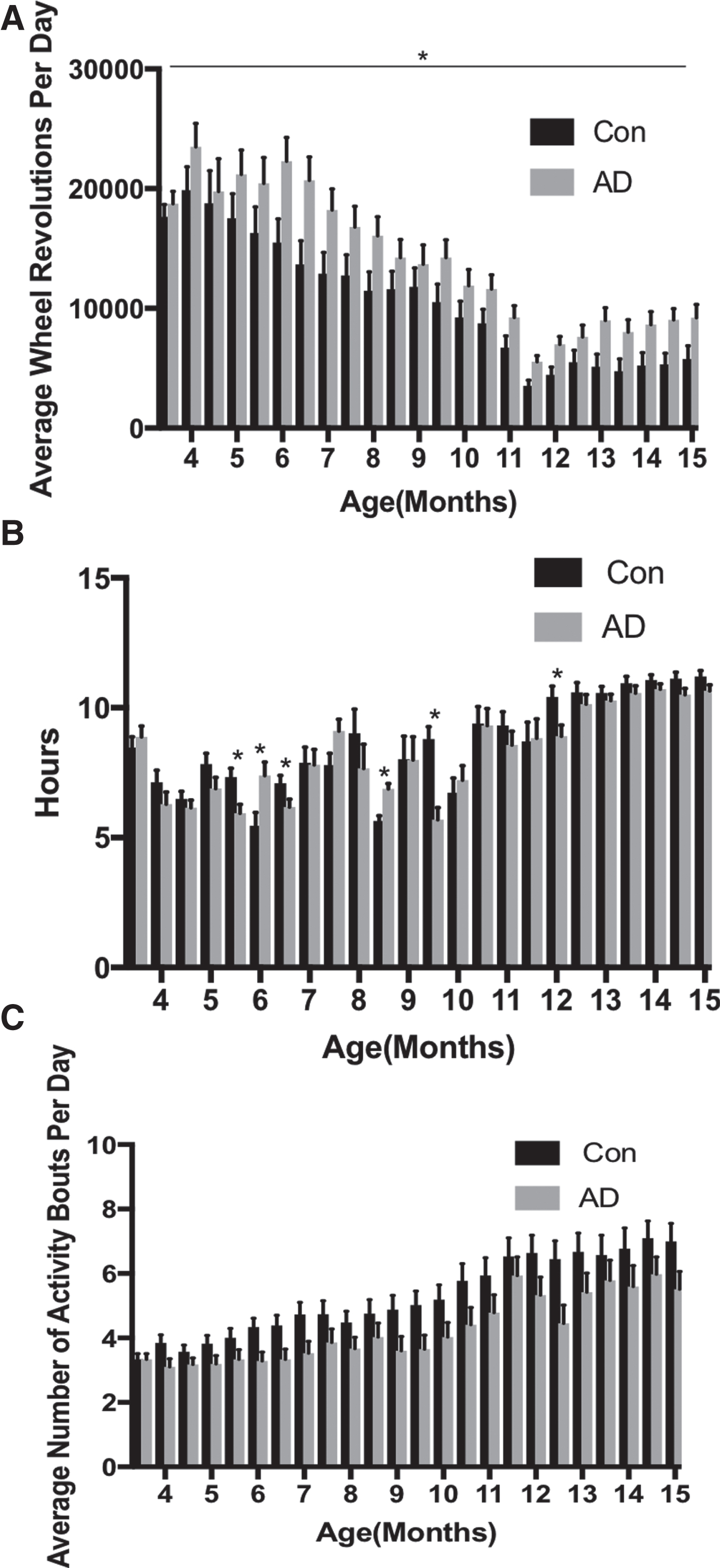
Activity rhythms are largely intact in APPNL-G-F mice up to 15 months of age. A) The average number of wheel revolutions per day was greater in the AD mice (n = 8) compared to controls (n = 8; p < 0.001). B) The length of alpha was different in some blocks with it longer for controls in blocks: 5 (p = 0.011), 7 (p = 0.0047), 13 (p < 0.001), and 18 (p = 0.023), and longer in the AD group in blocks 6 (p = 0.018) and 11(p = 0.001). C) The average number of activity bouts per day was not different between the groups.
Dark-dark
In the first dark-dark exposure, period was not different between the groups (Fig. 5A) but the amplitude was greater for the AD mice (Fig. 5B; F(1, 8.679) = 8.590, p = 0.017).
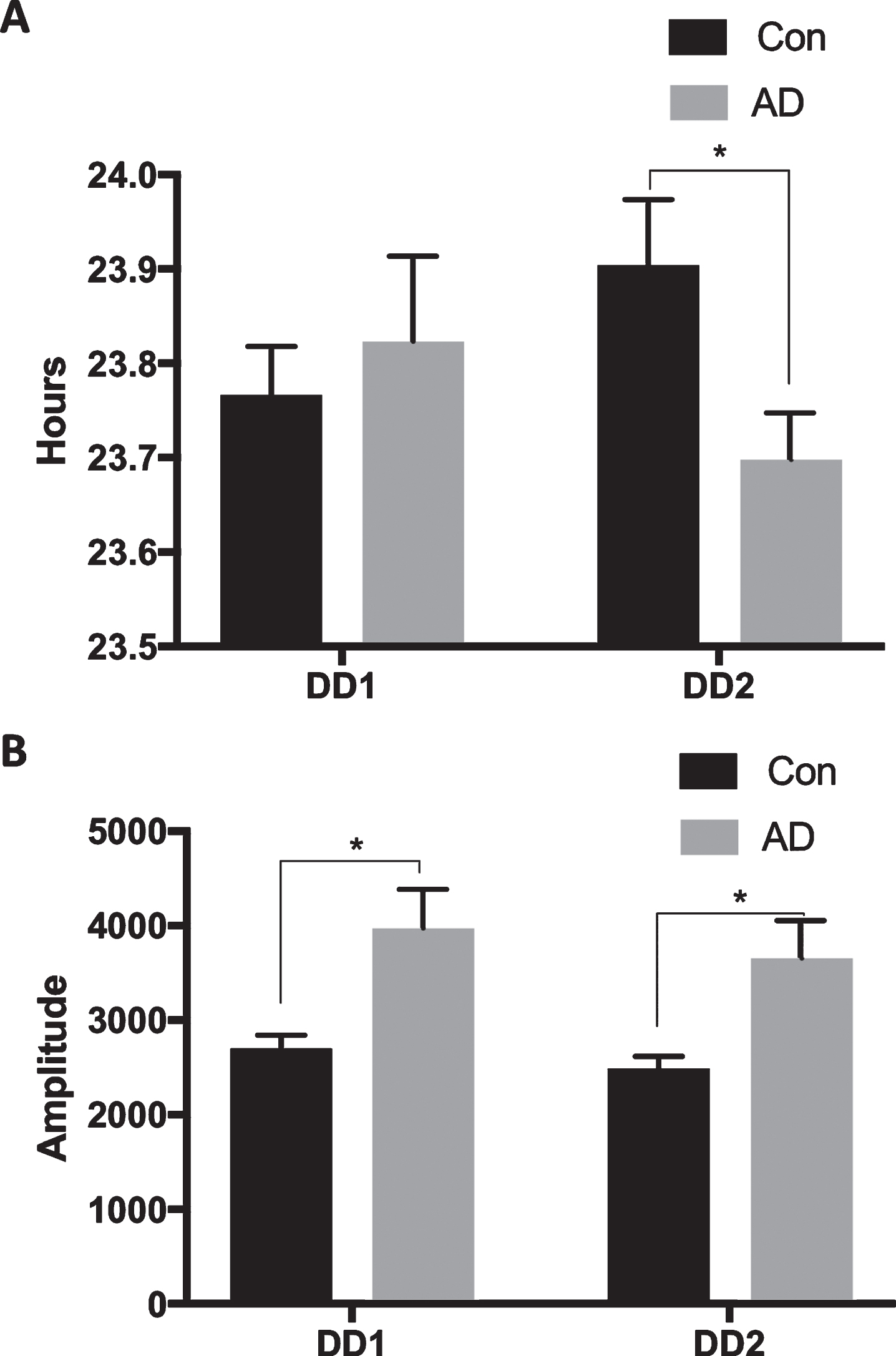
Slight difference in activity rhythms for APPNL-G-F mice in constant conditions. A) There were no differences between the groups in period during the first 9-day DD exposure (∼12 months of age), but period was advanced for the AD mice (n = 8) compared to controls (n = 8) in the second 9-day DD exposure (∼15 months of age; 0.031). B) Amplitude was greater for the AD mice in both the first (p = 0.017) and second (p = 0.023) DD exposures.
In the second dark-dark exposure, period was significantly shorter for the AD mice (Fig. 5A; F(1, 14) = 5.733, p = 0.031), but with a greater amplitude than the control mice (Fig. 5B; F(1, 8.434) = 7.726, p = 0.023).
Shift
Nocturnality did not change after the shift, nor was there a difference between the groups (Fig. 6A). Phase was significantly delayed after the shift (Fig. 6B; F(1, 14) = 34.447, p < 0.001, η2p = 0.711); however, there was no difference between the groups (F(1, 14) = 3.597, p = 0.079, η2p = 0.204), nor was the interaction significant (F(1, 14) = 2.895, p = .111, η2p = .171).

Response to a four-hour phase advance in lights on is not exacerbated in APPNL-G-F mice. A) Nocturnality (percentage of dark activity) was not different between the groups. B) The onset of activity in relation to lights off (phase) was not different between the groups.
T21
The mice were able to entrain to the T21 as indicated by a significant decrease in period to approximately 21 h during the six days of the T21 (Figs. 7 and 8A; F(1.019, 11.206) = 1184.717, p < 0.001, η2p = 0.991). Not surprisingly, the shift period was less than the periods during pre (p < 0.01) and post (p < 0.01); however, there was no difference between pre and post. Amplitude was also affected by the T21 (Figs. 7 and 8B; F(2, 22) = 39.642, p < 0.001, η2p = 0.783), with it being less during T21 exposure compared to pre (p = 0.008) and post (p < 0.01). Nocturnality and phase, also indicated entrainment as there was no change during or after the T21, respectively (Figs. 8C and 8D).

Two-month-old APPNL-G-F mice can entrain to six days of a T21 (L12:D9) cycle. A) A representative actogram from an APPNL-G-F mouse (Shift; n = 12). B) A representative χ2 periodogram from an APPNL-G-F mouse in the ten days prior to T21 exposure. C) A representative χ2 periodogram from an APPNL-G-F mouse in the six days of T21 exposure. D) A representative chi squared periodogram from an APPNL-G-F mouse ten days after T21 exposure. B, C, and D are from the same animal. Actogram represents behavior from approximately two months of age to four months of age. Days since placed in wheels are represented on the Y axis of the actograms. The grey shaded regions of the actograms indicate lights off.

Two-month-old APPNL-G-F mice can entrain to six days of a T21 (L12:D9) cycle. A) Period significantly decreased during T21 exposure to ∼21 hours (n = 12; p < 0.001) and increased post shift (p < 0.001). B) Amplitude significantly decreased during T21 exposure (p = 0.008) and increased post shift (p < 0.001) C) Nocturnality (percentage of dark activity) did not change during T21 exposure. D) The onset of activity in relation to lights off (phase) was not different after T21 exposure.
Six months of age
Period, amplitude, and nocturnality (F(2, 25) = 2.749, p = 0.083) were not different among the groups (Fig. 9A-C). However, there was a difference between the groups in phase (Fig. 7D; F(2, 25) = 12.857, p < 0.001), with the AD group having a significantly delayed phase compared to controls (p = 0.016), and the shift group (p < 0.001).
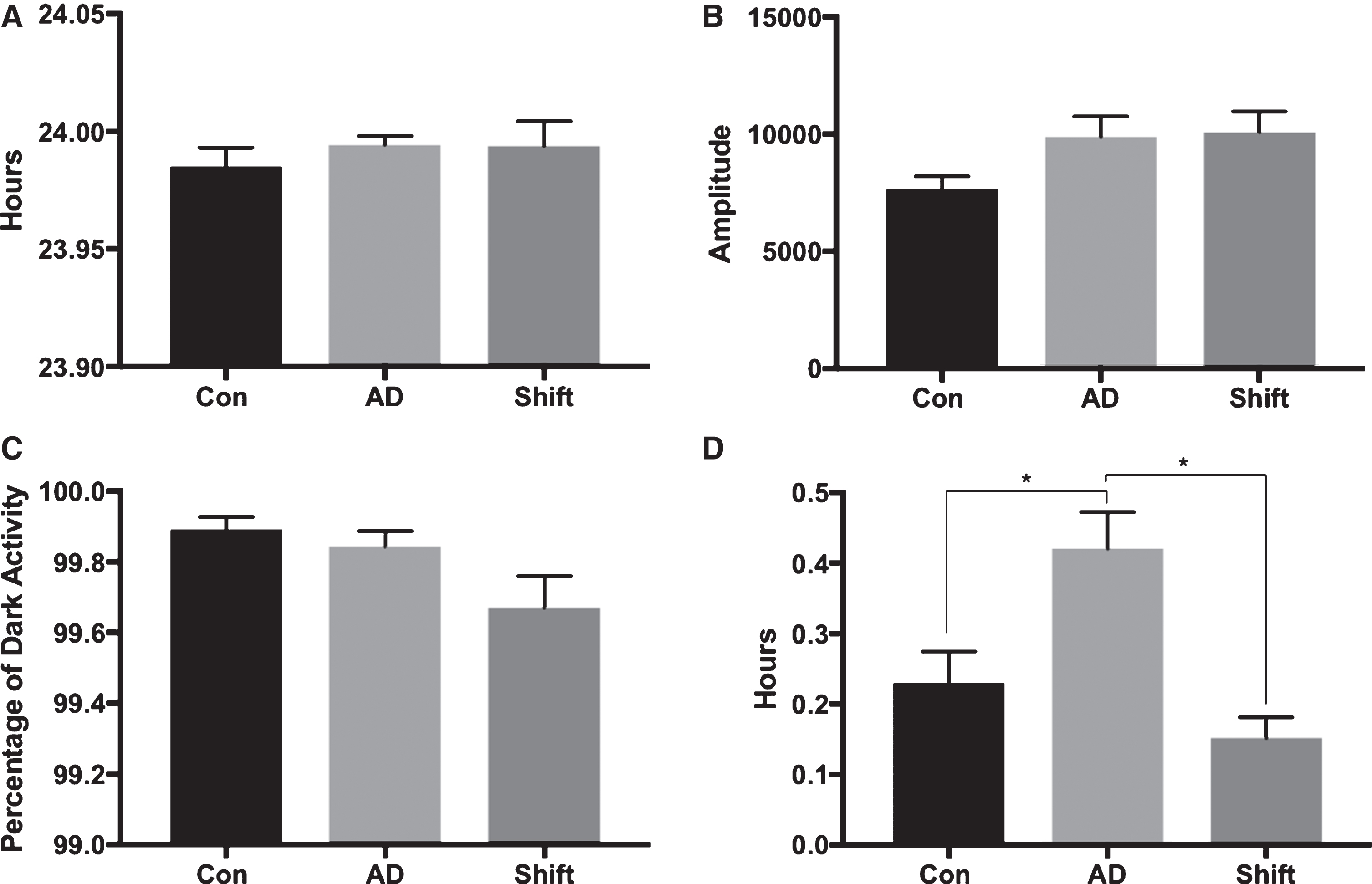
T21 exposure at two months of age does not affect the activity rhythm of six-month-old APPNL-G-F mice. A) Period was similar between the groups at six months of age. B) Amplitude was similar between the groups at six months of age. C) Nocturnality (percentage of dark activity) was not different between the groups at six months of age. D) Phase was not affected in the Shift group; however, the AD group had a significantly delayed phase compared to controls (p = 0.016) and the Shift group (p < 0.001).
Nine months of age
There were no differences between in the groups in period, amplitude, and phase (Fig. 10A, B, and D). However there was difference between the groups in nocturnality (Fig. 10C; F(2, 25) = 5.678, p = 0.009), with the shift group having decreased nocturnality when compared to the control (p = 0.028) and the AD (p = 0.027) groups.

T21 exposure at two months of age primarily does not affect the activity rhythm at nine months of age. A) Period was similar between the groups at six months of age. B) Amplitude was similar between the groups at six months of age. C) Nocturnality (percentage of dark activity) was slightly less for the Shift group compared to controls (p = 0.028) and AD mice (p = 0.027). D) Phase was similar between the groups.
DISCUSSION
There is a significant amount of variability in the circadian phenotype of the different transgenic mouse models of AD. This raises the question that circadian rhythm degradation in aging and AD, might be mediated by a pathological mechanism completely independent from changes in Aβ. The present study sought to characterize circadian rhythms in the novel APPNL-G-F mice that only involve upregulation of the Aβ fragment thought to underlie Aβ deposition in humans (Aβ42).
As the control and APPNL-G-F mice aged from 3–15 months of age, we observed hallmark changes in circadian rhythms such as, delayed phase of entrainment, more rhythm fragmentation, and decreased amount and amplitude of activity. In LD conditions, with aging we did not observe a change in period, but we did observe a lengthening in alpha, and surprisingly a very marginal increase in nocturnality.
There were slight differences between control and APPNL-G-F mice during the aging process. First, the APPNL-G-F animals were more active and had a higher rhythm amplitude than control animals in both LD and DD. Second, at approximately 15 months of age, the APPNL-G-F mice had a shorter period in DD than control animals. Except for these differences, other circadian parameters in LD conditions were similar between the groups, and there were no differences in response to a four-hour phase advance at approximately 15 months of age.
Lastly, in an attempt to accelerate circadian rhythm degradation, we subjected another group of APPNL-G-F mice to a non-24-hour light dark cycle at two months of age, which is when AD pathology begins in these animals. Surprisingly these mice were able to entrain to the 21-hour light dark cycle. Finally, nocturnality was decreased in these animals at nine months of age compared to the other control and APPNL-G-F mice at this age.
The finding of increased activity fits with other models: 3xTg [16], APP23 [13, 14], and AβPPswe/PSEN1A246E [31]. Although our animals were much older, as in the APPswe/PS1dE9 and AβPPswe/PSEN1A246E models we did not find a change in the circadian organization of behavior, which largely suggests normal functioning of the circadian system under a light dark cycle [19, 31]. A shorter free-running period in the APPNL-G-F mice in the present study is similar to the triple transgenic, 3xTg model [16]. Again, this is indicative of minor dysfunction as period with the light dark cycle was normal even at 15 months of age. Furthermore, we only found a shorter free-running period at 15 and not nine months of age, whereas in the 3xTg model, it was reported before the beginning of Aβ pathology which starts at approximately six months of age [12, 16].
In healthy humans, sleep deprivation increases overnight soluble Aβ production in cerebrospinal fluid by 25 to 30% [32]. With this in mind, we and others have tried to accelerate AD pathology by inducing circadian rhythm dysfunction. Except for the finding of less nocturnality at nine months of age, six days of a 21-hour day, which is equivalent to a 3-hour phase advance every day did not exacerbate circadian rhythm dysfunction. While a T21 is in the range of entrainment for C57Bl/6 mice, it is close to the limit of what is entrainable [33]. Entrainment to the 21-h cycle suggested that the circadian system is still very plastic and intact in the APPNL-G-F mice at two months of age; despite that, there are signs of AD pathology at this age. These data dovetail with the finding that compared to controls, a circadian rhythm disruption paradigm did not exacerbate circadian rhythm dysfunction in 3xTg mice [12]. It remains to be seen in this model, if more severe circadian rhythm manipulations such as sleep deprivation would exacerbate pathology.
The key finding of the present study is that the circadian dysfunction observed in the APPNL-G-F mice at 15 months of age was milder than other transgenic AD models, such as APP23, TgCRND8, 5xFAD, and 3xTg, which have altered activity patterns at this age [3, 34]. APPNL-G-F mice are similar to other models that have minor changes in activity rhythms, such as Tg2576, APPxPS1, and APPswe/PS1dE9, and AβPPswe/PSEN1A246E [17–19, 31]. Notably, the circadian rhythm dysfunction occurred long after the onset of AD pathology. Suggesting that in the APPNL-G-F model, early AD pathology does not include circadian dysfunction. As these animals show AD pathology from two months of age onward it is unlikely that circadian rhythm disruption is contributing to the AD phenotype. The finding that two-month-old APPNL-G-F mice could entrain to a 21-hour light dark cycle, further suggests that the circadian system is robust in these animals.
This study possessed a myriad of strengths and weaknesses. This is the first assessment of circadian rhythms in APPNL-G-F knock-in mice. This is noteworthy given the difference in how Aβ pathology is generated in the APPNL-G-F model compared to transgenic models. To our knowledge, continuous monitoring of activity for a year is quite rare in mouse models of AD as animals are usually monitored for a discrete amount of time surrounding different aging landmarks. As in Saito and colleagues [20], our control group were wildtype littermates (APP+/+), or in other words homozygous for the recessive trait of the knock-in. In contrast, it is quite common to use animals from a non-transgenic breeding stock. We view using controls from the same breeding stock as the transgenic animals as a more representative control as these animals have more similar genetic backgrounds than animals from a different breeding stock. However, it does not discount the fact that our control group also could have been experiencing circadian dysfunction. While the cognition of APPNL-G-F-/NL-G-F– is intact [20], future studies will need to confirm that circadian rhythms are also similar between these animals and C57BL/6 animals.
In terms of limitations, first, the use of wheels could have had a protective effect on circadian rhythm dysfunction. While it is possible that we would have observed more severe circadian dysfunction without wheels, as both groups were treated the same and given the similar genetic backgrounds, it is unlikely that the magnitude of the effects would change without wheels. Nonetheless, other studies that have found circadian effects in AD mouse models have also used wheels, so it is unlikely that this is the reason for the lack of an effect [16, 17]. Motion sensors, or the recent development of video-based monitoring systems [35], could measure circadian rhythms without the confounding effect of exercise on AD pathology and cognition. Second, one can argue that we stopped recording activity just when rhythms were starting to break down in the AD group. A shortening of period and increased amplitude could suggest that rhythms were about to break down further. It should be noted that amplitude was high in the APPNL-G-F mice throughout the entire period measured and period was only affected in constant and not in a light dark cycle. The fact that dysfunction was mild and occurred so late should negate this concern as well. Nonetheless, further studies should perhaps explore a longer recording period. Lastly, we only assessed male mice in the current study. In humans, the etiology of AD is different in males and females. However, this effect does not seem to carry over in most AD transgenic mouse models, which further questions the validity of these models. For example, there were not sex differences in the 3xTg model in terms of activity or response to phase advances of the light dark cycle [12, 16]. Nonetheless given the differences inherent to the model used here, we encourage the future assessment of female APPNL-G-F mice.
In summary, we observed mild circadian rhythm dysfunction at 15 months of age in a novel knock-in mouse model of AD. Most mouse models of AD typically report more dysfunction at this age. Two-month-old AD mice were also able to entrain to a non-24-hour light-dark cycle, which suggests that early AD pathology is not associated with circadian rhythm dysfunction. Our data open the possibility that some of the circadian rhythm dysfunction associated with transgenic models of AD might be a result of artificial phenotypes that are not representative of human AD.
Footnotes
ACKNOWLEDGMENTS
This work was supported by Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant #40352 and #06347 to MHM and RJM respectively, Alberta Innovate (MHM), Alberta Alzheimer Research Program (MHM & RJM), Alzheimer Society of Canada (MHM & RJM). We thank Takashi Saito and Takaomi C. Saido for establishing and graciously providing us with the APPNL-G-F/NL-G-F mice. Lastly, we thank Justin Q. Lee and Jogender Mehla for assisting with animal care.