
Abstract
Recent studies suggest that severity of asthma can be modulated by neuropsychiatric conditions, while the underlying mechanisms are not clear. Here, we used ovalbumin (OVA) to induce asthma in APP/PS1 mice, a mouse model of Alzheimer’s disease (AD), or in their wildtype control C57BL/6J mice. We found that all hallmarks of asthma by OVA were significantly attenuated in APP/PS1 mice, compared to age- and gender-matched C57BL/6J mice. Interestingly, significantly higher number of regulatory T cells (Treg) was detected in the APP/PS1 mouse lung, compared to those in the C57BL/6J mouse lung. Since Foxp3 is crucial for differentiation of naive T cells into Treg and is the most important marker for Treg, we examined the Foxp3 levels in the T cells from the lung of these mice. We found that the Foxp3 levels in the APP/PS1 mouse lung were significantly higher than those in the C57BL/6J mouse lung, likely resulting from reduced Foxp3 promoter methylation. Thus, our study suggests that AD may affect severity of asthma through methylation control of Foxp3 promoter in T cells.
INTRODUCTION
Asthma has several pathological features, including airway hyper-responsiveness (AHR), bronchial inflammatory responses, and airflow blockage. Most asthma cases belong to a specific asthma called allergic asthma, with characteristics of abnormal activation of type-2 T-helper cells, overproduction and secretion of IgE, eosinophilia, and T-cell dysfunction [1–3].
Regulatory T cell (Treg) is a specific subtype of T-cell that plays an inhibitory role during immune responses and AHR to maintain controllable immunity [4–6]. Treg is characterized with co-expression of surface markers of CD4 and CD25, and expresses a specific factor, forkhead box transcription factor p3 (Foxp3) [6–10]. Foxp3 mediates the differentiation of naive T cell towards Treg as well as the amplification of the differentiated Treg [11]. Therefore, Foxp3 is not only a specific marker for activated Treg, but also a trigger for Treg differentiation.
Recent studies caused attention to the interaction between asthma and neuropsychiatric conditions [12–19]. Although the knowledge on the impact of cognitive decline on asthma severity remains poorly established, cognitive decline has been shown as a core symptom in patients with Alzheimer’s disease (AD) [20], a chronic neurodegenerative disease characterized by continuous and progressive cholinergic failure [21]. It is intriguing that clinical evidence suggests that the status of AD may decrease AHR in response to cholinergic stimulation, resulting in attenuation of symptoms of asthma [22]. Moreover, altered T-cell functionality occurs in both asthma and AD, indicating that T-cell may be a bridge of the interaction between asthma and AD [23]. Since the molecular mechanisms that underlie this phenomenon has not been studied previously, here we addressed this question and focused on the role of Treg in an asthmatic AD mouse model, in which a traditional ovalbumin (OVA)-induced allergic asthma and a commonly applied ASS/PS1 AD model were used in combine.
MATERIALS AND METHODS
Experimental approval and animal handling
All experiments and mouse work were approved by the IACUC of TongJi University, and were performed in strict accordance to the Care and Use of Laboratory Animal Guideline, issued by TongJi University. Male C57BL/6J and APP/PS1 mice (Jackson Lab, Bar Harbor, ME, USA) were involved in this study.
Social recognition test
At 16 weeks of age, the mice were subjected to a social recognition test for evaluation of social recognition and memory. In this test, male mice at home cage were challenged 4 times with the same ovariectomized female mouse, and subsequently challenged in the 5th time with a novel ovariectomized female mouse. Each challenge lasted 5 min, followed by a 15 min break between two challenges. The time for an examined mouse to spend sniffing in the procedure was regarded as the social investigation time.
Morris water maze
Mice were trained for locating the visible escape platform in 3 trials with an interval of 30 min in the first 2 days. From the 3rd to the 7th day, the platform was placed under the water surface for each examined mouse to explore 3 times per day at an interval of 1 min. The time latency for the mouse to locate the hidden platform in each trial was recorded.
Plus-maze discriminative avoidance task (PM-DAT)
Mice were placed at the center of the apparatus with exposure to both illuminating light and cold air blowing, to allow them to enter the enclosed arm in the training trials. One day later, mice were placed in the same position for 3 min without aversive stimuli. The relative time for the mice to stay in the aversive enclosed arm during training and testing was recorded.
Ovalbumin-induced allergic asthma model
Male mice of 16 weeks of age (immediately after behavior tests) were sensitized by an intraperitoneal injection of 50μg OVA (grade V; Sigma-Aldrich, St. Louis, MO, USA) with 2 mg aluminum hydroxide gel (Sigma-Aldrich) at a frequency of one injection per week for 3 weeks (week 0– 2). Afterwards, the mice were challenged with an intranasal administration of 50μg OVA at a frequency of one injection per two weeks for the next 7 weeks (week 3– 9). Control mice received injection of saline of same dosage and frequency. At week 10, the mice were examined before sacrifice for specimens.
Airway hyper-responsiveness
AHR was assessed by restrained invasive plethysmography 24 h after the last intranasal OVA challenge. A small incision was made onto the anesthetized mice for trachea exposure to allow a cannula connecting an inline nebulizer and ventilator to be inserted. Mice were then challenged with aerosolized PBS followed by increasing doses of methacholine (Sigma-Aldrich). Airway resistance and dynamic lung compliance were assessed.
Bronchoalveolar lavage, lung digestion, and isolation of treg
Bronchoalveolar lavage fluid (BALF) was obtained by instilling 3 washes of 0.5 ml PBS with 0.1% BSA. Centrifugation was performed to enumerate cells for further analysis. Lung digestion was initiated with infusion of 0.8 ml 300 U/ml collagenase type I (Sigma-Aldrich), and 50 U/ml DNase I (Roche, Nutley, NJ, USA) in RPMI 1640 culture media (Sigma-Aldrich), followed by tissue removal, mincing and further at 37°C for 30 min. Treg was analyzed and purified by flow cytometry with FITC-conjugated anti-CD4 and PE-cy7-conjugated anti-Foxp3 antibodies (Becton-Dickinson Biosciences, San Jose, CA, USA) on a FACSAria cell sorter (Becton-Dickinson Biosciences). Data were analyzed using Flowjo software v11 (Flowjo LLC, Ashland, OR, USA).
Quantitative reverse transcription polymerase chain reaction (RT-qPCR) and methylation-specific PCR (MS-PCR)
TaqMan quantitative PCR (RT-qPCR, two step) was applied to examine the relative levels of the genes, using GAPDH as an internal control. The total RNA was isolated using TRIzol® reagent (Invitrogen), according to the manufacturer’s instructions. RNA was then reverse transcribed into cDNA using the High Capacity cDNA Reverse Transcription kit (Applied Biosystems, Foster City, CA, USA). TaqMan Gene Expression Assays were done using pre-designed primers on an ABI 7500 Fast Real-time PCR system (Applied Biosystems). Relative quantification of gene expression was performed using the 2-Δ ΔCt method.
MS-PCR was performed to identify the methylation status of Foxp3. Bisulfite conversion of DNA was done with an EZ-96 DNA Methylation-Gold kit (Zymo Research, Irvine, CA, USA). Each 15 ml MS-PCR included 7.5 ml Qiagen Multiplex Master Mix (Qiagen, Hilden, Germany), 3 ml Q solution (Qiagen), 0.3 ml forward primers, 0.3 ml reverse primers, 2.9 ml of RNAse-free water and 1 ml of bisulfite-converted template DNA. The primer sequences used in the MS-PCR were shown in Table 1. MS-PCR products were analyzed by agarose gel electrophoresis. Samples were defined as methylated or unmethylated depending upon the visual band amplified with methylated or unmethylated primers.
Immunostaining
Immunostaining for Aβ was performed using a rabbit anti-Aβ antibody (EMD Millipore, Billerica, MA, USA). Immunostaining for phosphorylated Tau (p-Tau) was performed using a goat anti-p-Tau antibody (Santa Cruz Biotechnology, Dallas, TX, USA). Quantification was based on 3 slides with a distance of 100μm in between.
Statistics
GraphPad Prism software version 7 (GraphPad Software, Inc., La Jolla, CA, USA) was applied in the statistical analysis. Analysis was done by one-way ANOVA with a Bonferroni correction, followed by Fisher’s Exact Test upon necessity. All values are depicted as mean±standard deviation from 10 individuals and are considered significant if p < 0.05. Data were represented as mean±SD.
RESULTS
APP/PS1 mice exhibit compromised performance in behavioral tests
APP/PS1 is a mouse model of AD, and its wildtype control is C57BL/6J. Age-matched male APP/PS1 and C57BL/6J were kept till 16 weeks of age and received behavior tests. In the Social Recognition Test, APP/PS1 mice exhibited significantly poorer habituation responses to the repeatedly introduced stimulus and significantly poorer dishabituation responses to the novel stimulus compared to the control C57BL/6J mice (Fig. 1A). APP/PS1 mice exhibited similarly poorer performance in a Morris Water Maze test (Fig. 1B) and in a PM-DAT (Fig. 1C, D). These data confirm that both recognition and learning capacity are impaired in APP/PS1 mice.

APP/PS1 mice exhibit compromised performance in behavioral tests. Sex- and age- matched APP/PS1 and control C57BL/6J mice at 16 weeks of age received behavioral tests. A) Social Recognition Test. B) Morris Water Maze test. C, D) Plus-Maze discriminative avoidance Task. *p < 0.05. N = 10.
Hallmarks of OVA-induced asthma are attenuated in APP/PS1 mice
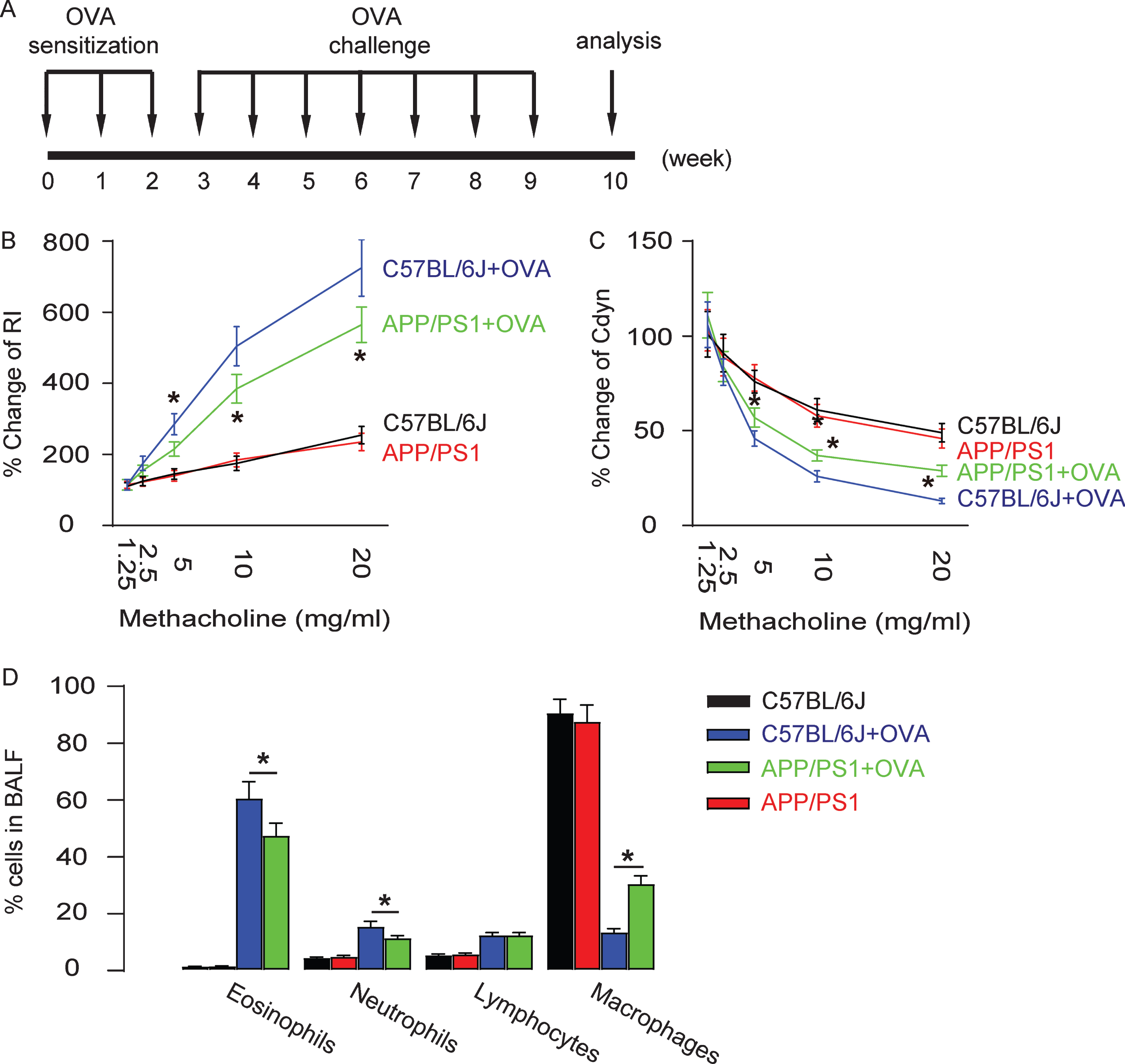
A model of OVA-induced asthma was then applied to these sex- and age-matched APP/PS1 and control C57BL/6J mice. The experiment took a duration of 10 weeks, including 2 weeks of sensitization of the mice by alum-adsorbed OVA and 6 weeks of challenge of exposure of the mice to repeated airway provocation (Fig. 2A). At the 10th week, the features of asthma were analyzed. The establishment of airway hypertension was demonstrated by a dose-dependent increase in lung resistance (Fig. 2B) and a dose-dependent decrease in dynamic lung compliance in response to a cholinergic stimulus (methacholine; Fig. 2C), both of which were found significantly attenuated in APP/PS1 mice, compared to those in control C57BL/6J mice. Eosinophilic accumulation in the pulmonary airway is another feature of OVA-induced allergic asthma, and was assessed at the end of the experiment. We found that the influx of inflammatory eosinophils increased up to 60% of total BALF cells in OVA-treated mouse lung in control C57BL/6J mice, but the degree of this increase was significantly attenuated in APP/PS1 mice (Fig. 2D). These data suggest that OVA-induced asthma is attenuated in APP/PS1 mice.
Hallmarks of OVA-induced asthma are attenuated in APP/PS1 mice. A) Experimental illustration for OVA sensitization, inhalation challenge, and analysis on sex- and age-matched APP/PS1 and control C57BL/6J mice at 16 weeks of age when AD-features were established in APP/PS1 mice. Lung resistance (B) and dynamic lung compliance (C) in response to increasing doses of methacholine. (D) Percentage of leukocytes in BALF. *p < 0.05. N = 10.
Augmentation of amyloid-β deposition and p-Tau levels in APP/PS1 mouse brain
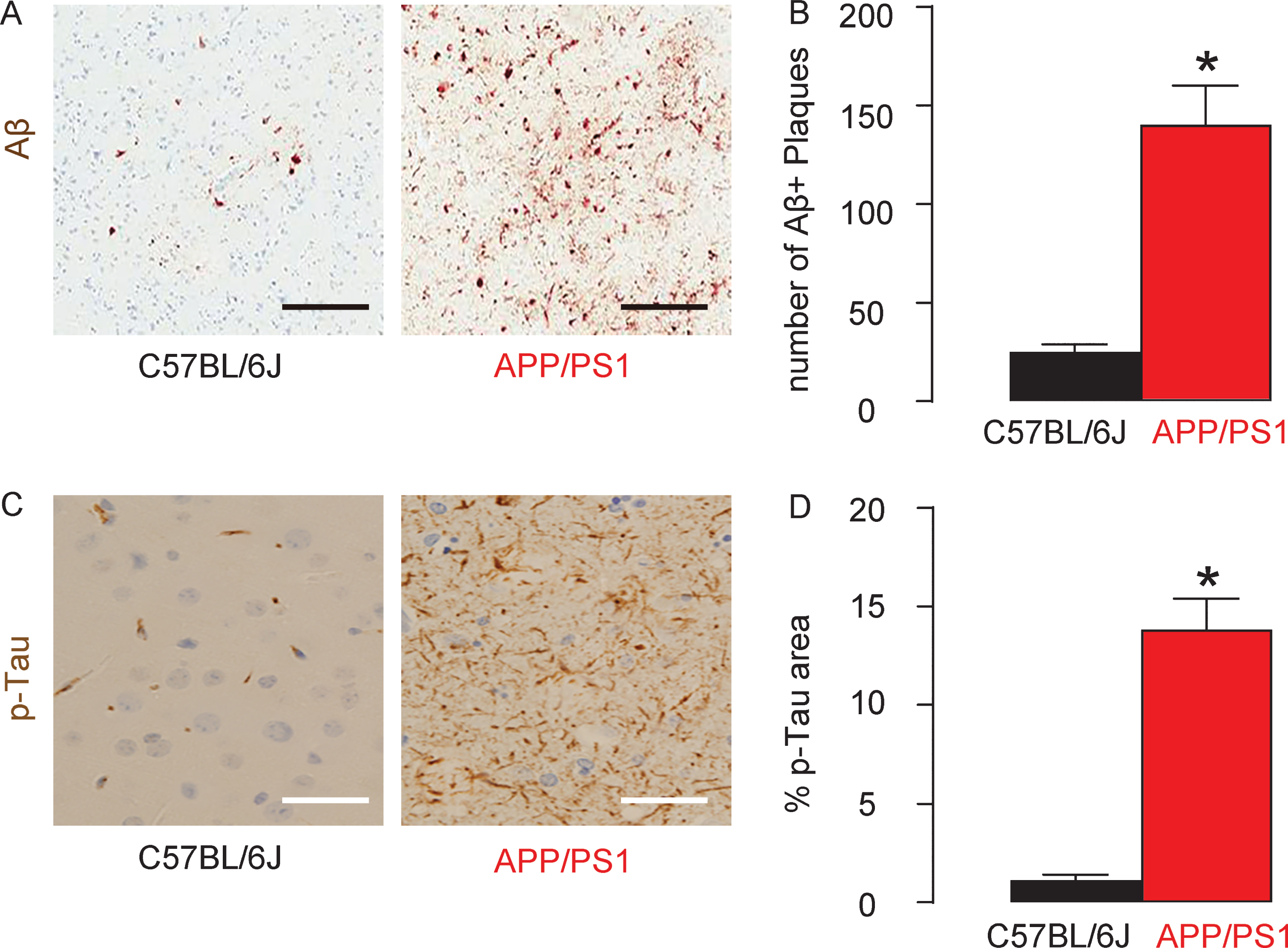
At sacrificed, the mouse brain was evaluated the levels of amyloid-β (Aβ) plaques and p-Tau in mouse hippocampus. We detected a significantly higher number of Aβ plaques in the hippocampus region in APP/PS1 mice, compared to the control C57BL/6J mice, shown by representative images (Fig. 3A) and by quantification (Fig. 3B). Similarly, we detected a significantly larger p-Tau area in the hippocampus region in APP/PS1 mice, compared to the control C57BL/6J mice, shown by quantification (Fig. 3C), and by representative images (Fig. 3D). These data confirmed that APP/PS1 mice automatically develop neurodegenerative pathological changes in the brain.
Augmentation of Aβ deposition and p-Tau levels in APP/PS1 mouse brain. The number of Aβ plaques in the mouse brain was quantified in the hippocampus, shown by representative images (A) and by quantification (B). The phosphorylated Tau (p-Tau) area in the mouse brain was quantified in the hippocampus, shown by quantification (C), and by representative images (D). *p < 0.05. N = 7. Scale bars are 100μm.
An APP/PS1 background is susceptible for Treg activation by OVA
Since Treg is a potent inflammation suppressor in asthma, we analyzed the percentage of Foxp3+ cells (Treg) in the total CD4+ T helper cells by flow cytometry in the lung digests of these mice (Fig. 4A). We found no difference in Treg between APP/PS1 and the control C57BL/6J mice that were not treated with OVA, while OVA hardly induced increases in Treg in the control C57BL/6J mice, but did induce increases in Treg in OVA-treated APP/PS1 mice (Fig. 4A, B). Similarly, Foxp3 mRNA levels in CD4+ T-cells were not different between APP/PS1 and the control C57BL/6J mice that were not treated with OVA, while OVA hardly induced increases in Foxp3 mRNA levels in CD4+ T-cells in the control C57BL/6J, but did induce increases in Foxp3 mRNA levels in OVA-treated APP/PS1 mice (Fig. 4C). Together, these data suggest that an APP/PS1 background is susceptible for Treg activation by OVA.
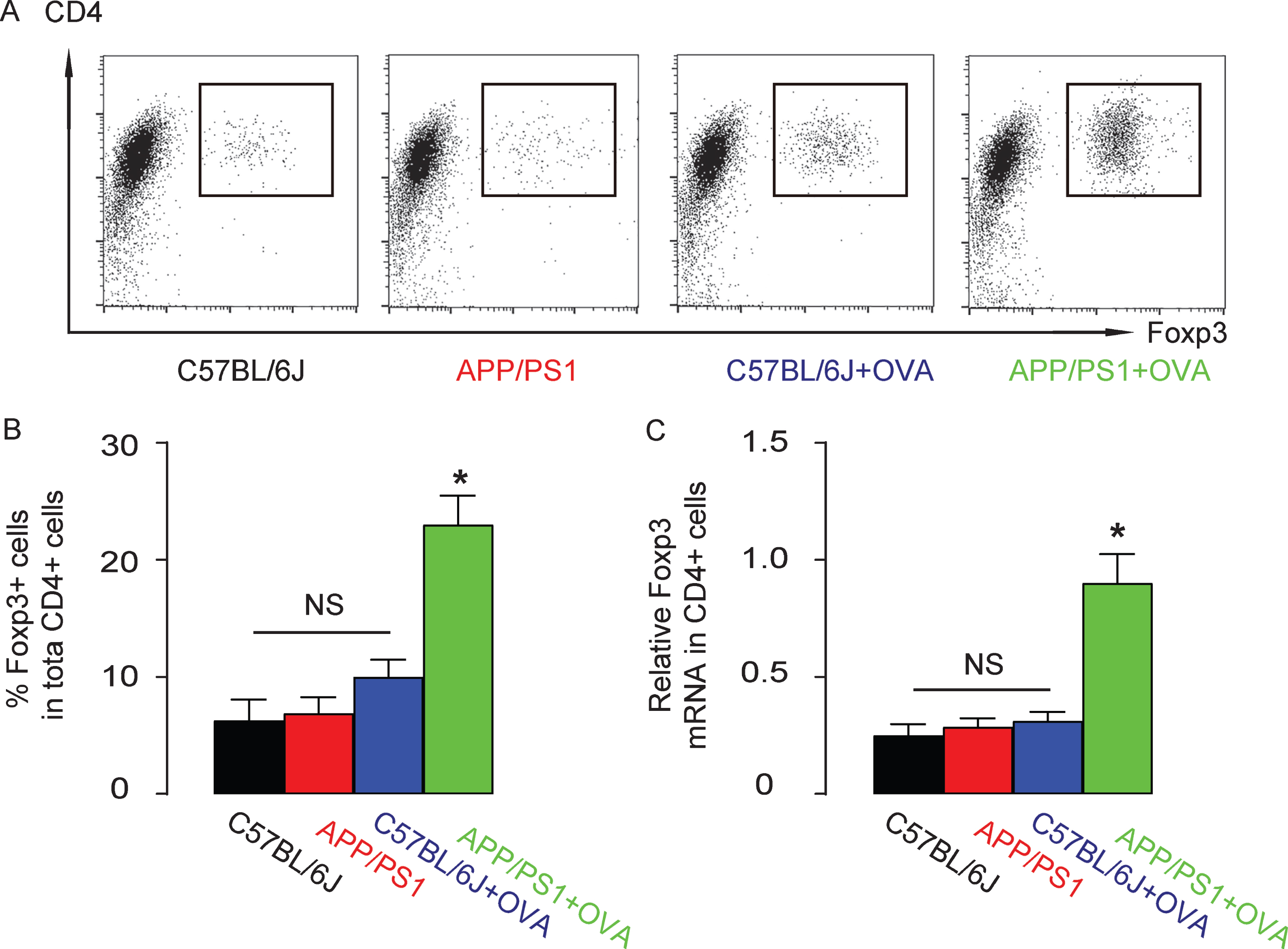
An APP/PS1 background is susceptible for Treg activation by OVA. Treg cells were purified from the lung digests of the mice from 4 groups based on CD4 and Foxp3 positivity by flow cytometry, shown by representative flow charts (A) and by quantification (B). (C) Foxp3 mRNA levels in CD4+ T-cells by RT-qPCR. *p < 0.05. NS, non-significant. N = 10.
Promoter and CpG regions are major methylation sites for Foxp3 promoter in APP/PS1 mice
The high Foxp3 levels are responsible for the Treg activation. Transcriptional activation of Foxp3 expression is known to be epigenetically regulated, e.g., methylation. Foxp3 has 3 known sites that are susceptible for methylation, which are promoter region, enhancer region and CpG region. We analyzed methylation status at these sites in purified airway CD4+ T-cells in different groups of the mice. We found that the methylation status of Foxp3 on promoter region was only significantly reduced in OVA-treated APP/PS1 mice, compared to the other 3 groups (Fig. 5A). The methylation status of Foxp3 on enhancer region was not significantly altered in all 4 groups (Fig. 5B). The methylation status of Foxp3 on CpG region was also only significantly reduced in OVA-treated APP/PS1 mice, compared to the other 3 groups (Fig. 5C). Thus, promoter and CpG regions are major methylation sites for Foxp3 promoter in APP/PS1 mice

Promoter and CpG regions are major methylation sites for Foxp3 promoter in APP/PS1 mice. Methylation status of 3 known sites on Foxp3 [promoter (A), enhancer (B), and CpG region (C)] were assessed by MS-PCR. *p < 0.05. NS, non-significant. N = 10.
DISCUSSION
The intriguing relationship between neurodegenerative diseases like AD and asthma has been surprisingly detected in recent years. Here, we showed that in an AD background, the Treg was activated and expanded through transcriptional activation of Foxp3, resulting from reduction in Foxp3 promoter methylation. This is consistent with the numerous reports acknowledging the control of Foxp3 expression by promoter methylation [24–26]. Indeed, promoter methylation is a key suppressor of gene expression, and Foxp3 is a key determinant for Treg, the expression of which is coordinated at a level of primarily epigenetic modification of the promoter [27]. The epigenetic modifications, including sumoylation/desumoylation, acetylation/deacetylation, methylation/demethylation, etc. These biological events play a substantial role in control of gene expression [28]. Typically, methylation increases the difficulty of access of transcriptional factors to the promoter region and thus inhibits gene expression. On the other hand, demethylation opens the access and promotes gene expression [28].
Treg is activated in the setting of AD, and then affects the severity of asthma. Treg is well-known to be the key controller of the process of asthmatic pathology. Clonal deletion, anergy, immune deviation, and Treg-mediated immune suppression all contribute to the immune tolerance to self- or exogenous antigens/allergens [11]. The activated and amplified Treg by AD thus replenishes the need for Treg to reduce the AHR, bronchial inflammatory responses, and airflow blockage in asthma [1–3], as found in the current study. The molecular mechanisms that underlie the activation of Treg by AD are somehow less understood. In one pioneer study, the authors used an adoptive transfer model to figure out that Treg improved cognitive function and reduced deposition of Aβ plaques in AD animals and concluded that systemic Treg administration has a potential to ameliorate AD progression and could be an effective AD treatment. Thus, the activation of Treg by AD could be a protective mechanism of the body to contradict the pathological progression of the disease.
Our findings suggest that decreased methylation of Foxp3 promoter on CD4+ T-cells was beneficial in both AD and asthma. The Foxp3 has 3 sites that are important for expression control and are regulated by methylation. The first site is the promoter, which is a classic TATA and CAAT-box-containing region located 6.5 kb upstream of exon one [28]. Demethylation of this promoter site opens binding sites for activator protein 1 (AP-1), signal transducer and activator of transcription 5 (Stat5), nuclear factor of activated T cells (NFAT) and TGFβ-inducible early gene 1 (TIEG1) [28]. The second site is at the enhancer, and works as a TGFβ sensor, since it is a TGFβ-sensitive element with binding sites for NFAT and mothers against decapentaplegic homologues (SMADs) [28]. The third site is a Treg-cell-specific demethylated region characterized with highly conserved CpG-rich nuclear acids [28]. This region was already known to be fully demethylated in Treg cells and fully methylated in non-Treg T-cells [28]. Since we found that the methylation status of promoter and CpG-rich regions was greatly altered in AD, transcriptional factors associated with these two regions may be essential for post-demethylation gene activation.
To the best of our knowledge, this is the first study to address the molecular mechanisms that underlie the interaction between AD and asthma using Treg as a bridge. Future studies may address the effects of asthma-associated Treg changes on the progression of AD, to help fully interpretation of this crosstalk.