
Abstract
Upregulation of Rho-associated protein kinase 2 (ROCK2) hallmarks the progression of neurodegenerative diseases. However, the molecular mechanisms underlying the regulation of ROCK2 expression are not clear and thus addressed in the current study. We generated a subacute model of Parkinson’s disease in mice with a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) method. The MPTP model was validated by impaired rotational behavior of the mice upon apomorphine exposure, astrocytic activation, and reduction in tyrosine-hydroxylase-positive neurons in the mouse striatum. Moreover, MPTP induced increases in ROCK2 protein but not in ROCK2 mRNA. Further analysis showed that MPTP inhibited the expression of microRNA-291 (miR-291), which suppressed ROCK2 mRNA via 3’-UTR-binding in neuronal cells to increase ROCK2 protein. Intracranial injection of miR-291 four days before MPTP alleviated the impaired rotational behavior of the mice upon apomorphine exposure, MPTP-induced astrocytic activation, and the reduction in tyrosine-hydroxylase-positive neurons in the mouse striatum. Together, these data suggest that miR-291 has a protective role in neurodegeneration, likely through regulation of ROCK2.
INTRODUCTION
Alzheimer’s disease and Parkinson’s disease are the most common neurodegenerative diseases in aged people [1–4]. Selective loss of dopaminergic (DA) neurons and a shortage of dopamine in the striatum hallmark the most important pathological characteristics of Parkinson’s disease [5]. L-DOPA is currently used in clinic and has only limited symptom-releasing effects [6–8]. Since the etiology of Parkinson’s disease is not completely discovered, extensive research is needed to locate novel therapeutic targets for effective treatments.
Rho-associated protein kinase 2 (ROCK2) is highly expressed in the central nervous system, including the brain and the spinal cord [9]. Its expression level increases with age and appears to be associated with neurodegenerative processes with experimental evidence, e.g., ROCK2 activates LIM (“LIM” derives from the first 3 proteins with these 2 domains at their N-terminus and a homeodomain at their C-terminus: lin-11, Is11, and mec-3) domain kinase, which subsequently promotes the activation of tensin homolog deleted on chromosome ten (PTEN) and apoptosis antigen 1 (APO-1 or Fas) [10]. Recently, ROCK2 was discovered to be a major controller of axonal degeneration, neuronal death, and axonal regeneration in the central nervous system, and is suggested to play critical roles in the development of Parkinson’s disease [11].
MicroRNAs (miRNAs) are a group of non-coding RNAs of 15∼22 nucleotides and their function is control of protein translation through interference with mRNA at the 3′-untranslated region (3′-UTR) site [12–15]. Although miRNAs have been a hot topic in the past few years and many miRNAs have been extensively studied in different diseases, the involvement of microRNA-291 (miR-291) in any diseases or models has not been reported.
Here, we generated a subacute model of Parkinson’s disease in mice with a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) method. The MPTP model was validated by impaired rotational behavior of the mice upon apomorphine exposure, astrocytic activation, and reduction in tyrosine-hydroxylase (TH)-positive neurons in the mouse striatum. Moreover, MPTP induced increases in ROCK2 protein but not in ROCK2 mRNA. Further analysis showed that MPTP inhibited the expression of miR-291, which suppressed ROCK2 mRNA via 3′-UTR-binding in neuronal cells to increase ROCK2 protein. Intracranial injection of miR-291 four days before MPTP alleviated the impaired rotational behavior of the mice upon apomorphine exposure, MPTP-induced astrocytic activation, and the reduction in TH-positive neurons in the mouse striatum.
MATERIALS AND METHODS
Protocol approval
The experimental protocol and approaches including ethic issues on animal work have been approved by the research committee at the Wenzhou Medical University.
Mouse treatment
Fourteen 1-year-old male C57BL/6J mice were obtained from the SLAC Laboratory Animal Co. Ltd (Shanghai, China), and were fed standardly. In the first experiment, mice were randomly assigned into two groups: a saline-only treated group (n = 5), which received subcutaneous saline injection daily for 5 days; and a MPTP group (n = 5), which received subcutaneous MPTP injection (Sigma-Aldrich, St. Louis, MO, USA) daily for 5 days at a dose of 25 mg/kg. Four days after the last injection of MPTP of saline, mice first underwent rational behavior test and then were sacrificed for other analyses. In the second experiment, mice were randomly assigned into two groups: an MPTP + scrambled group (n = 5), which received intracranial injection of scrambled plasmid at the beginning of the experiment and subcutaneous MPTP injection (Sigma-Aldrich) daily for 5 days at a dose of 25 mg/kg, beginning 4 days after intracranial injection of scrambled plasmid; an MPTP + miR-291 group (n = 5), which received intracranial injection of miR-291 plasmid (5′-TTTCACGAAGGTGAAACACACG-3′) at the beginning of the experiment and subcutaneous MPTP injection (Sigma-Aldrich) daily for 5 days at a dose of 25 mg/kg, beginning 4 days after intracranial injection of miR-291 plasmid. Four days after the last injection of MPTP, mice first underwent rational behavior test and then were sacrificed for other analyses.
Rational behavior test
At the end of experimental, the animals’ tendency to rotate in response to apomorphine was assessed. In the test, the mice were secured in a rotometer bowl for counting rotations. Apomorphine (Sigma-Aldrich) was subcutaneously given at the back of mouse neck at 0.5 mg/ml diluted in sterile 0.02% ascorbic acid saline solution prepared in dark and protected with aluminum foil. The number of neck rotations was continuously recorded for 1 h.
Prediction of microRNA targets and 3′-UTR luciferase-reporter assay
TargetSan (https://www.targetscan.org) was used to predict targets for miRNAs. The wildtype ROCK2 3′-UTR reporter plasmid (WT ROCK2 3′-UTR) and ROCK2 3′-UTR reporter plasmid with a mutate at the miR-291 binding site (ROCK2 3′-UTR mut) were both purchased from Creative Biogene (Shirley, NY, USA). Cells were collected 24 h after transfection for dual-luciferase reporter assay (Promega, Beijing, China), according to the manufacturer’s instructions.
Human cortical neuron cell and transfection
A human cortical neuron cell line, HCN-1A, was purchased from ATCC (ATCC, Rockville, MD, USA), and were cultured in Dulbecco’s Modified Eagle’s Media (DMEM) supplemented with 15% fetal bovine serum (Invitrogen, Carlsbad, CA, USA) in a humidified chamber with 5% CO2 at 37°C. For control of miR-291 levels, miR-291 or antisense of miR-291 (as-miR-291) was expressed under a CMV promoter in a plasmid. A plasmid carrying a scrambled sequence (scrambled) was used as a control. All the plasmids were obtained from Origene (Shanghai, China). The plasmids were transfected into HCN-1A cells at a concentration of 150 nmol/l using Lipofectamine-3000 (Invitrogen), receiving about 85% transfection efficiency, according to co-expression of GFP reporter in the plasmid construct.
Immunohistochemistry
Perfusion with 4% paraformaldehyde (PFA) was done at the end of the experiments before collection of brain samples, which were again fixed in 4% PFA at 4°C overnight, transferred to 30% sucrose overnight, and then sectioned into 6μm sections. The slides were stained with ABC method (Sigma-Aldrich) with rabbit-anti-mouse antiglial fibrillary acidic protein (GFAP; Millipore, Billerica, MA, USA), followed by hematoxylin counterstain, or with immunofluorescent method, using rabbit-anti-mouse TH antibody (ab112, Abcam, Billerica, MA, USA).
ELISA
Protein was extracted with RIPA lysis buffer (Sigma-Aldrich) on ice. Protein concentration was determined with BCA protein assay kit (Sigma-Aldrich). ELISA was done with either a mouse ROCK2 ELISA kit (LSBio LifeSpan BioSciences, Seattle, WA, USA) or a TH ELISA kit (LSBio LifeSpan BioSciences).
RT-qPCR
Total RNA was extracted with a miRNeasy mini kit (Qiagen, Hilden, Germany). Complementary DNA (cDNA) was primed from total RNA using the Omniscript reverse transcription kit (Qiagen). Quantitative PCR (RT-qPCR) were performed with QuantiTect SYBR Green PCR Kit (Qiagen). All primers were purchased from Qiagen. Data were collected and analyzed using 2–▵▵Ct method for quantification of the relative mRNA expression levels. Values of genes were obtained by sequential normalization to GAPDH and experimental controls.
Statistical analysis
GraphPad prism version 8.0 (GraphPad Software, Inc. La Jolla, CA, USA) was used to analyze the data with a one-way analysis of variance (ANOVA) test followed by the Fisher’s Exact Test to compare two groups. All values represent the mean±standard deviation (SD). A value of p < 0.05 was considered statistically significant after Bonferroni correction.
RESULTS
MPTP mouse model
Mice were randomly assigned into two groups: saline-only treated group (n = 5), which received subcutaneous saline injection daily for 5 days; MPTP group (n = 5), which received subcutaneous MPTP injection daily from for 5 days at a dose of 25 mg/kg. Four days after the last injection of MPTP of saline, mice first underwent rational behavior test and then were sacrificed for other analysis. In rational behavior test, we found that MPTP significantly increased the number of the turns of the mice (Fig. 1A). In astrocytes, MPTP can be converted into 1-methyl-4-phenylpyridinium by monoamine oxidase-B to create neurotoxicity. We found that MPTP induced significantly increases in the number of antiglial fibrillary acidic protein (GFAP)-positive astrocytes in the mouse striatum, shown by representative images (Fig. 1B), and by quantification (Fig. 1 C). TH is capable of catalyzing L-DOPA, a precursor for dopamine, and this process requires presence of co-factors O2 and iron. Loss of TH promotes neurodegeneration [16, 17]. We found that MPTP induced significantly decreases in TH (Fig. 2C). Together, these data suggest that MPTP induces neurodegenerative features of H2S may alleviate the low dopamine levels induced by MPTP, a characteristic for Parkinson’s disease.
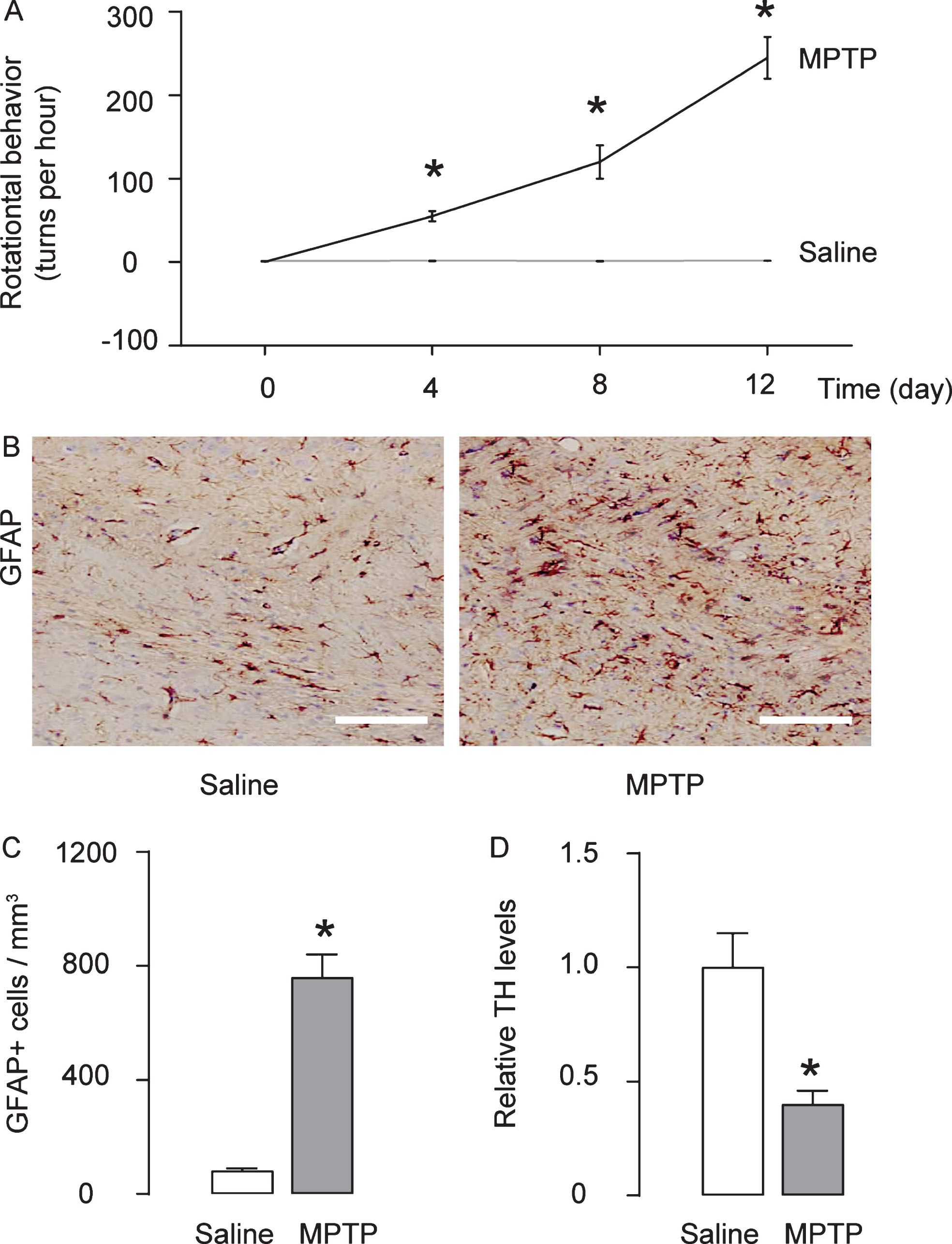
Establishment of mouse model of Parkinson’s disease by MPTP. Mice were randomly assigned into two groups: saline-only treated group (n = 5), which received subcutaneous saline injection daily for 5 days; MPTP group (n = 5), which received subcutaneous MPTP injection daily for 5 days at a dose of 25 mg/kg. Four days after the last injection of MPTP of saline, mice first underwent rational behavior test and then were sacrificed for other analysis. A) The effects of MPTP on the behavioral symptoms of the mice were examined in an apomorphine-induced rotational behavior test. B, C) The number of antiglial fibrillary acidic protein (GFAP)-positive astrocytes was determined in the mouse striatum, by representative immunohistochemistry images (B), and by quantification (C). D) ELISA for antityrosine hydroxylase in mouse brain. *p < 0.05. N = 5. Scale bars are 50μm.

MPTP upregulates ROCK2 protein without affecting ROCK2 mRNA. A, B) The levels of ROCK2 in mouse brain were analyzed by ELISA (A) and by RT-qPCR (B). *p < 0.05. NS, non-significant. N = 5.
MPTP upregulates ROCK2 protein without affecting ROCK2 mRNA
We analyzed ROCK2 levels in the mouse striatum, and found that the levels of ROCK2 protein were significantly increased in MPTP-treated mice (Fig. 2A). However, the ROCK2 mRNA levels were not changes in MPTP-treated mice (Fig. 2B), suggesting presence of post-transcriptional regulation of ROCK2.
MiR-291 inhibit protein translation of ROCK2
We did bioinformatics analysis on the ROCK2-targeting miRNAs, and found that miR-291 was the one that significantly suppressed by MPTP (Fig. 3A). A specific miR-291 binding site on the 3′-UTR (from 2809th to 2815th base site) of the ROCK2 mRNA was shown in the illustration (Fig. 3B). For assessing whether binding of miR-291 to ROCK2 mRNA may alter the ROCK2 levels, we either overexpressed miR-291, or inhibited miR-291 in a human neuronal cell line HCN-1A) and confirmed the alteration of miR-291 levels in these cells by RT-qPCR (Fig. 3C). HCN-1A cells were then co-transfected with one of these plasmids that alter miR-291 levels and one plasmid carrying either a luciferase reporter for 3′-UTR of ROCK2 mRNA or a luciferase reporter for 3′-UTR of ROCK2 mRNA with mutate at the miR-291 binding site (mut). The luciferase activities were determined (Fig. 3D), suggesting that miR-291 targets 3′-UTR of ROCK2 mRNA to inhibit its translation in neuronal cells.
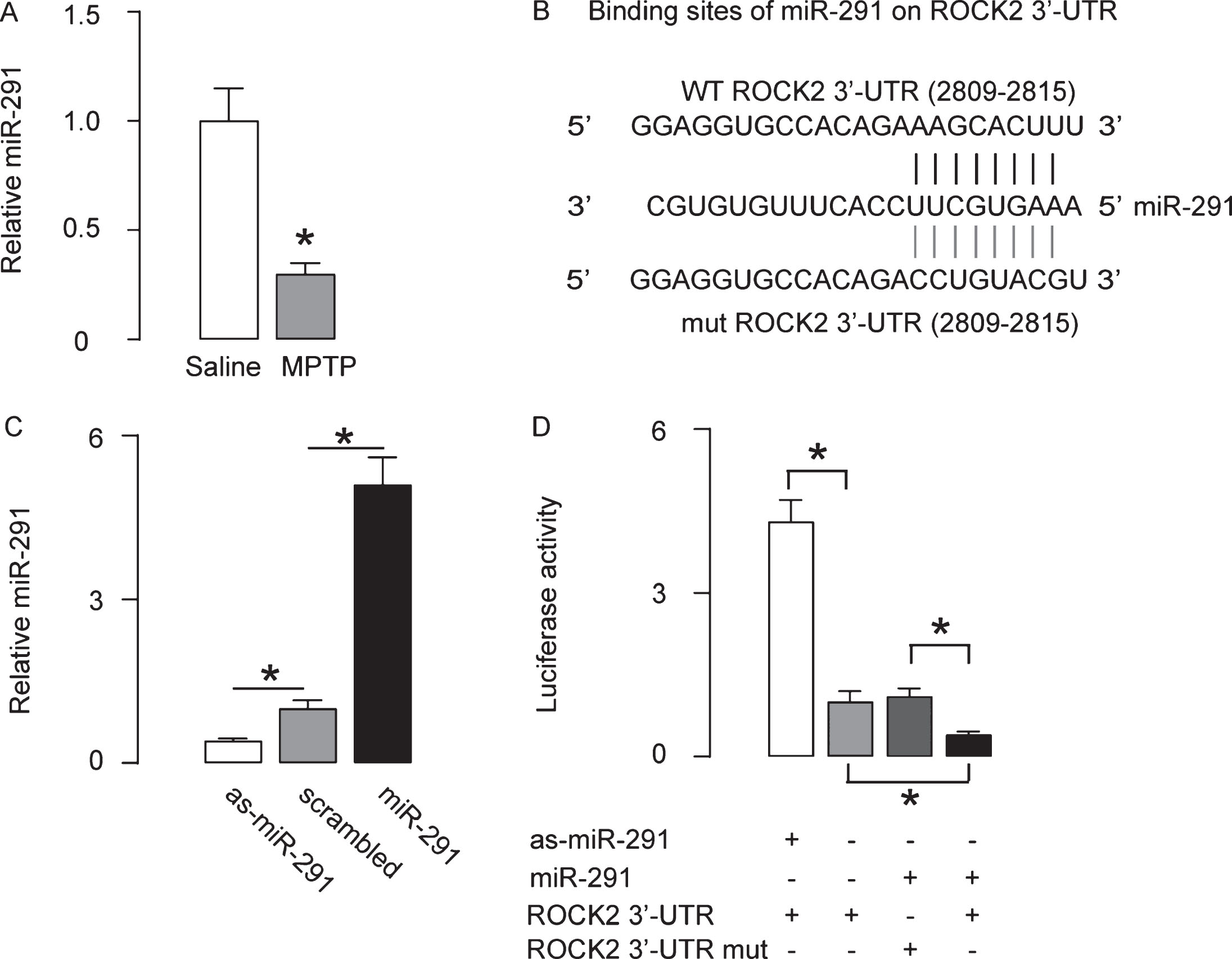
MiR-291 inhibit protein translation of ROCK2. A) The miR-291 levels in mouse striatum by RT-qPCR. B) Bioinformatics analysis showing a specific miR-291-binding site on the 3′-UTR of the ROCK2 mRNA. C) RT-qPCR for miR-291 on miR-291-, or as-miR-291-, or scrambled- transfected HCN-1A cells. D) HCN-1A cells were then co-transfected with one of these plasmids that alter miR-291 levels and one plasmid carrying either a luciferase reporter for 3′-UTR of ROCK2 mRNA or a luciferase reporter for 3′-UTR of ROCK2 mRNA with mutate at the miR-291 binding site (mut). The luciferase activities were determined. *p < 0.05. NS, non-significant. N = 5.
Intracranial injection of miR-291 alleviates MPTP-induced features of Parkinson’s disease
In order to evaluate if miR-291/ROCK2 contributes to MPTP-induced features of Parkinson’s disease, we did another experiment. Mice were randomly assigned into two groups: MPTP + scrambled group (n = 5), which received intracranial injection of scrambled plasmid at the beginning of the experiment and subcutaneous MPTP injection daily from for 5 days at a dose of 25 mg/kg, ever since 4 days after intracranial injection of scrambled plasmid; MPTP + miR-291 group (n = 5), which received intracranial injection of miR-291 plasmid at the beginning of the experiment and subcutaneous MPTP injection daily for 5 days at a dose of 25 mg/kg, beginning 4 days after intracranial injection of miR-291 plasmid. Four days after the last injection of MPTP, mice first underwent rational behavior test and then were sacrificed for other analysis (Fig. 4A). We found that intracranial injection of miR-291 impaired rotational behavior of the mice upon apomorphine exposure (Fig. 4B), reduced ROCK2 protein levels (Fig. 4C), alleviated MPTP-induced astrocytic activation (Fig. 4D, E), reduced TH levels (Fig. 4F), and reduced the size of TH-positive fibers in the mouse striatum (Fig. 4G). Together, these data suggest that miR-291 has a protective role in neurodegenerative diseases like Parkinson’s disease, mediated through regulation of ROCK2 (Fig. 5).
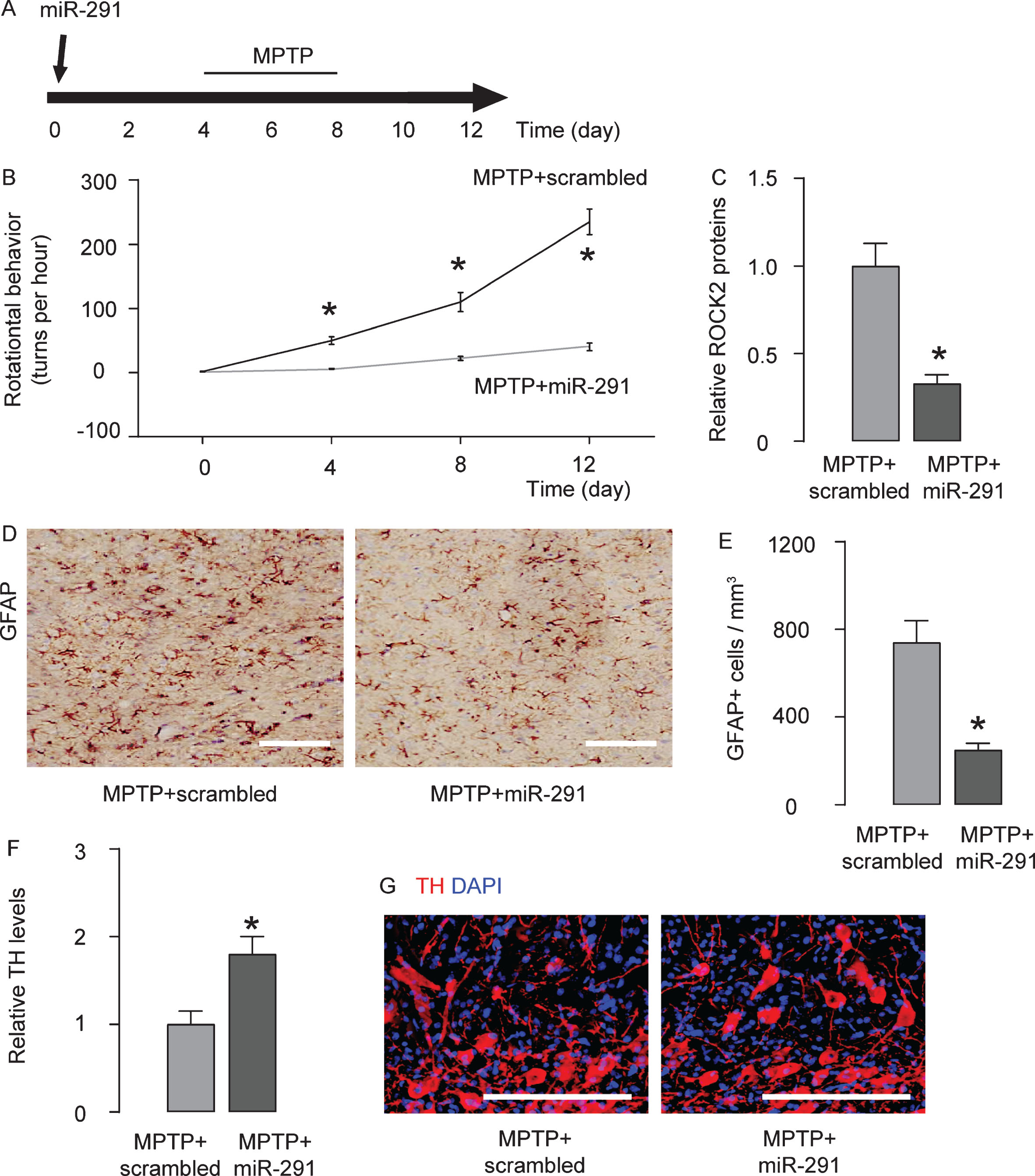
Intracranial injection of miR-291 alleviates MPTP-induced features of Parkinson’s disease. A) In order to evaluate if miR-291/ROCK2 contributes to MPTP-induced features of Parkinson’s disease, we did another experiment. Mice were randomly assigned into two groups: MPTP + scrambled group (n = 5), which received intracranial injection of scrambled plasmid at the beginning of the experiment and subcutaneous MPTP injection daily from for 5 days at a dose of 25 mg/kg, ever since 4 days after intracranial injection of scrambled plasmid; MPTP + miR-291 group (n = 5), which received intracranial injection of miR-291 plasmid at the beginning of the experiment and subcutaneous MPTP injection daily for 5 days at a dose of 25 mg/kg, begining 4 days after intracranial injection of miR-291 plasmid. Four days after the last injection of MPTP, mice first underwent rational behavior test and then were sacrificed for other analyses. B) The behavioral symptoms of the mice were examined in an apomorphine-induced rotational behavior test. C) ELISA for ROCK2 levels. D, E) The number of antiglial fibrillary acidic protein (GFAP)-positive astrocytes was determined in the mouse striatum, by representative immunohistochemistry images (D), and by quantification (E). F) ELISA for antityrosine hydroxylase (TH) in mouse brain. G) Staining for TH in mouse brain. DAPI, nuclear staining. *p < 0.05. N = 5. Scale bars are 50μm.
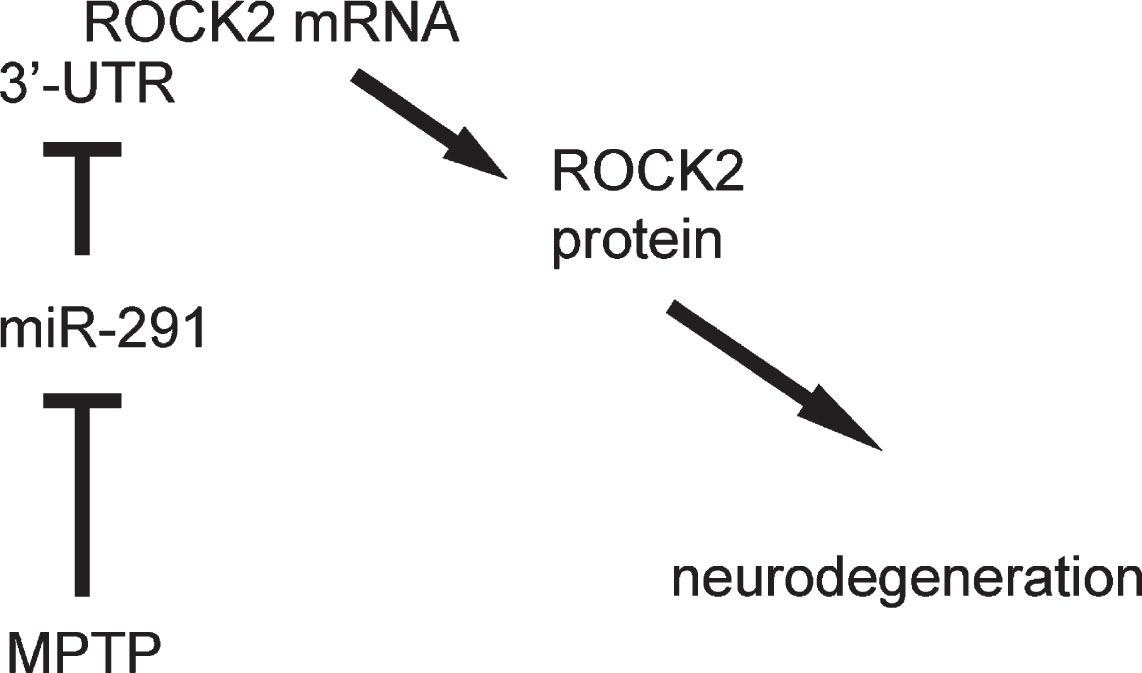
Schematic of the model. miR-291 has a protective role in Parkinson’s disease, likely through regulation of ROCK2.
DISCUSSION
Rho/ROCK/LIMK pathway plays a critical role in the progression of neurodegeneration, but the control of ROCK2 is poorly defined [18–21]. GFAP is a cellular marker for astrocytes, which exert both neuroprotective and neurodegenerative roles, depending on the microenvironment. MPTP induces injury and causes abnormal increases in astrocytes, which subsequently induced neurodegeneration. In the current study, using a mouse model for Parkinson’s disease, we showed that ROCK2 mRNA and protein levels were differentially affected by the MPTP, a lipophilic protoxin that rapidly crosses the blood–brain barrier following systemic injection and enters astrocytes causing damages to the neuronal cells by its catalyzed form, 1-methyl-4-phenyl-2, 3, dihydropyridinium [22]. These data suggest presence of a post-transcriptional control of ROCK2, probably by miRNAs. Indeed, miRNAs may bind to 3′-UTR of mRNA of target gene, and this binding either abolishes protein translation without affecting bound mRNA, or induces mRNA instability to enhance its degradation [23]. In the first case, protein is inhibited but the mRNA is not affected. In the second case, both protein and mRNA will decrease [23]. Here, it appeared that the mRNA of ROCK2 was not affected and thus it seemed that it belonged to the case 1, in which instability of the bound mRNA did not occur.
The miR-291 is a rarely studied miRNA. Actually, we did not find any published studies on the role of miR-291 in any diseases. Interestingly, here we detected significant alteration of the miR-291 level by MPTP. By bioinformatic analysis, we found that miR-291 has many predicted target genes, like mitochondrial coiled-coil domain 1, ligand dependent nuclear receptor corepressor, zinc finger protein 800, clock circadian regulator, RAB22A, member RAS oncogene family, suppressor of variegation 3–9 homolog 1, left-right determination factor 2, B-cell translocation gene 1, anti-proliferative cyclin-dependent kinase inhibitor 1A (p21, Cip1), E2F transcription factor 2, lin-52 homolog (C. elegans), ankyrin repeat domain 32, nuclear factor I/B, etc., suggesting that miR-291 may have undiscovered functions that deserves further analysis in future [24]. Of note, p21 and E2F are both included in this list, suggesting a possible paradoxical role of miR-291 in controlling cell cycle progression in stem cell and tumor cell [25, 26].
To the best of our knowledge, it is the first study to show the involvement of miR-291 and ROCK2 in the pathogenesis of Parkinson’s disease. Our study should provide new insights into the mechanisms underlying development and progression of Parkinson’s disease.
DISCLOSURE STATEMENT
Authors’ disclosures available online (https://www.j-alz.com/manuscript-disclosures/19-0832r1).