
Abstract
Progressive neurodegenerative diseases represent some of the largest growing treatment challenges for public health in modern society. These diseases mainly progress due to aging and are driven by microglial surveillance and activation in response to changes occurring in the aging brain. The lack of efficacious treatment options for Alzheimer’s disease (AD), as the focus of this review, and other neurodegenerative disorders has encouraged new approaches to address neuroinflammation for potential treatments. Here we will focus on the increasing evidence that dysbiosis of the gut microbiome is characterized by inflammation that may carry over to the central nervous system and into the brain. Neuroinflammation is the common thread associated with neurodegenerative diseases, but it is yet unknown at what point and how innate immune function turns pathogenic for an individual. This review will address extensive efforts to identify constituents of the gut microbiome and their neuroactive metabolites as a peripheral path to treatment. This approach is still in its infancy in substantive clinical trials and requires thorough human studies to elucidate the metabolic microbiome profile to design appropriate treatment strategies for early stages of neurodegenerative disease. We view that in order to address neurodegenerative mechanisms of the gut, microbiome and metabolite profiles must be determined to pre-screen AD subjects prior to the design of specific, chronic titrations of gut microbiota with low-dose antibiotics. This represents an exciting treatment strategy designed to balance inflammatory microglial involvement in disease progression with an individual’s manifestation of AD as influenced by a coercive inflammatory gut.
INTRODUCTION
People are living longer now than ever before [1, 2]. With increasing aging populations in some of the most populous countries in the world, there is an unfortunate concomitant increase in those suffering from age-related, chronic diseases, such as dementia [2, 3]. Dementia is an umbrella term for brain diseases characterized by gradual cognitive decline of mental faculties, including impairment of memory, reasoning, judgment, and language, that are beyond the normal scope of healthy aging [4]. The most prevalent type of dementia, and the subject of this review, is Alzheimer’s disease (AD), a progressive neurodegenerative disorder defined by the extracellular deposition of amyloid plaques largely composed of amyloid β-protein (Aβ) and intracellular neurofibrillary tangles of hyperphosphorylated tau protein [5, 6]. The plaques and tangles that are the pathological hallmarks of AD delineate it further as a protein misfolding disease, where aberrant protein structures and aggregates have been shown to be synaptotoxic to neurons and associated with the debilitating memory deficits of AD [5, 7]. Curiously, the vast majority of AD presents sporadically, and the extent of plaque deposition does not always dictate whether a person will exhibit memory deficits [7]; facts both of which have led many to explore other peripheral factors that lead to neurodegeneration [8–11].
The peripheral factor of interest in this review is the gut microbiome and the extent to which its dysbiosis and metabolite production may influence neurodegenerative disease progression. We shall explore the emerging connections between age-related immune dysregulation and inflammation, neurodegenerative disease, and the gut. This review will also highlight the necessity for AD staging and associated AD microbiome/metabolite profiles as justifications for implementing the low-dose, chronic use of targeted antibiotics and other potential drugs to titrate toxic gut bacteria. This approach may enhance the innate immune effect of protective, peripheral microglia cells over time to treat neurotoxic mechanisms associated with the progression of AD before irreversible decline sets in.
Aging and immune function
The greatest risk factor for developing AD is age [12]. Aging is also associated with a chronic state of low-grade inflammation called “inflammaging” that is characterized by higher concentrations of circulating pro-inflammatory mediators such as IL-6 and C-reactive protein [13]. Persistent, dysregulated inflammatory responses are thought to be responsible, in part, for the elderly’s increased susceptibility to infection [14]. An aging person’s immune system typically reacts less robustly to environmental stimuli than a younger person’s does, with altered B and T cell development and differentiation resulting in an overall decrease in functional specific immunity [14]. The decline in lymphocyte diversity has also been associated with chronic inflammation, as well as with viral and bacterial infections, particularly by the expansion and over-stimulation of memory T cells and exhaustion of the naïve T cell pool [15]. Additionally, a cohort of 40 AD patients were discovered to have higher proportions of activated, late-differentiated CD4+ CD25+ T cells and decreased portions of a naïve T cell pool, potentially due to chronic stimulation of the immune system [16]. It is important to note that a generalized dysregulation of aging immune function does not singularly guarantee the development of dementia as gene expression and promotion of lymphocyte diversification can be dependent on diet, exercise, hormone regulation, chemical exposure, and pathogen infection [17, 18].
Due to the difficulty in separating age-causative inflammation from exogenous factors that induce inflammation, it remains unclear to what extent which inflammatory mediators relative to others will result in harmful systemic inflammation nor is there a known, definitive source of inflammaging [19]. During an acute inflammatory response, such as to an injury or infection, the innate immune system regulates the activation of an adaptive immune response that normally resolves after pathogen clearance that is followed by wound-healing mechanisms. Chronic inflammation is thought to be the result of chronic stimuli, such as cellular debris of damaged host cells or other damage-associated molecular patterns (DAMPs), that the adaptive immune system recognizes and upregulates an innate immune response for removal [20]. A modest decrease in number of circulating peripheral dendritic cells in aging may contribute to less robust adaptive immune responses as we age [21–23]. This effect is worsened with corticosteroid administration, marked by reduction in IFN-α production and a decrease in number of circulating dendritic cells [24]. Reduction in the body’s ability to adequately detect and address emergent infections may prolong inflammation through impairment of T cell differentiation to quiescent states [15], suggesting that the aging body is inclined to exhibit chronic inflammation due to dysregulation in the immune system.
There is no known, definitive cause of AD. However, in light of the multifactorial nature of the disease, it may be that AD is the cumulative result of a decades-long inflammatory response to CNS stressors. There is emerging evidence of viral and microbial presence in the AD brain and potential connections of gut microbiome diversity to neuroinflammation, amyloidosis, and neurodegeneration. For example, multi-omic experiments found that human herpesvirus HHV-6A and HHV-7 are more prevalent in AD brain compared to controls, and that multiple viruses, most notably HHV-6A, are linked to AD risk genes including ones related to processing amyloid-β protein precursor (AβPP) [25]. Importantly, amyloid β-protein (Aβ) results from the proteolytic cleavage of AβPP and Aβ fibrils are suggested to have broad spectrum anti-microbial properties capable of agglutination and disrupting cellular membranes to induce ion dyshomeostasis [26]. Further, low-grade herpesvirus, fungal, and bacterial infections have been associated with seeding Aβ deposits in the brain, suggesting that amyloidosis could be a generalized, innate immune response to the persistent invasion or incorrect perception of pathogens within the CNS [26–28]. The sources of peripheral inflammation are myriad but the progression of AD neurodegeneration is increasingly linked to heightened levels of circulating cytokines that stimulate further cytokine secretion in the CNS, by way of signaling via the vagus nerve or by directly crossing the blood-barrier to activate microglia and astrocytes toward a proinflammatory state [29]. Proinflammatory microglia activation is associated with other neurodegenerative diseases, and recent studies with transgenic ALS model mice show that peripheral macrophages and spinal cord microglia have distinct gene expression regulation patterns and that modulation of peripheral macrophages shift microglia away from damaging pro-inflammatory behavior to neuron and synapse support [30]. Many more studies will be required to better understand the interplay between the surveillance mechanisms at play in the CNS that contribute to neurodegeneration. Identifying how peripheral factors may influence inflammatory mechanisms for a particular neurodegenerative disease will provide much needed insight for potent, targeted therapeutics and low-dose, chronic treatments that could modulate harmful microbes.
Early neurodegenerative disease biomarkers and efforts to profile the microbiome
Owing to the fact there are no efficacious treatments available to slow or stop the progression of neurodegeneration that can develop over decades, an important proactive approach to combating dementia will involve identification of early biomarkers associated with increased risk for disease before symptomatic onset occurs. A cohort study of 3,565 individuals found that the relationship between cerebrospinal fluid (CSF) levels of Aβ and tau by categorization into amyloidosis and neurodegenerative NIA-AA profiles was profile-dependent and that higher Aβ42 CSF levels are related to earlier stages of AD [31]. This further supports evidence that Aβ pathology may precede and influence tau pathology in AD [32–34]. A longitudinal study of 2014 participants within the Alzheimer’s Disease Neuroimaging Initiative database developed a stochastic Markov model to predict the time it takes to present with clinical AD based on four broad biomarker categories, including: cognitive functional score; CSF and plasma levels of Aβ, neurofilament light (NfL), and tau; and normalized MRI volumetric measurements of the brain [35]. Although the study could not identify single biomarkers that delineate the clinical progression of AD for an individual, NfL was identified as a potential early marker for neurodegeneration [35]. Additionally, population CSF levels of Aβ42 increased at higher rates years before tau CSF levels did, before the symptomatic onset of AD [35]. Biomarker studies are widely important to help uncover how a disease presents itself, but age-related diseases that develop over long timeframes vastly complicate this process. CSF biomarkers observed at late stages of disease may define how the brain has compensated for extensive, irreversible neuronal damage but may not fully allude to earlier, deterministic neuropathologic mechanisms. Biomarkers present in serum may reveal information that is difficult to delineate from other concomitant health issues so it will be necessary to exploit a diverse array of biological sampling to capture a broad picture of a given neurodegenerative state so that targeted, efficacious treatment becomes a reality.
Determination of a neurodegenerative disease microbiome profile, through identification of pathogenic microbes and metabolic pathways, represents an appealing way to tailor a treatment plan based on a respective patient’s manifestation of dementia. The general concept of microbiome profiling has been thoroughly explored in animals and in small human cohorts. Some early efforts were made to identify the stable, healthy human microbiome composition into clustered groupings called enterotypes. The enterotype concept was introduced by comparative 16S rRNA metagenomic analysis of fecal samples from 22 individuals from 4 countries when three enterotypes emerged, each with Bacteroides, Prevotella, and Ruminococcus genera most abundant, respectively [36]. As the healthy human gut composition is complex, it is important to identify beneficial commensal organisms from pathogenic ones, particularly for the context of chronic disease. It has been noted that grouping together microbiome constituents into distinct clusters of species, as is done with enterotypes, may unfortunately over-simplify and overlook the contribution of lesser abundant species to the health of the host, or even mask the inter-variability of an individual’s enterotype constituents to another of the same enterotype [37]. This is further complicated by confirmed diagnosis of AD occurring at autopsy which is obviously much too late for early intervention. Inclusion parameters to determine the composition of the AD gut microbiome must be multi-factorial to account for any differences related to gender, age, cognitive function, medication use, APOE genetic markers, brain imaging, and biomarkers.
Treatment of neurodegenerative disease is hindered largely in part by a lack of analytical biomarker testing that may predict disease progression early on, most importantly before the advent of cognitive decline. To broadly address this need in the context of general health, a comparative study of 4,347 publicly available human fecal metagenomes produced a microbiome signature of 50 species associated with the healthy human gut, determined by the Gut Microbiome Health Index, and may predict whether an individual possesses a healthy microbiome based on the overall abundance of health-prevalent species relative to health-scarce species [38]. Even though the results of these kinds of studies vary considerably, a major asset is the data is very often publicly available and future efforts to determine a metabolic microbiome profile for the staging of neurodegenerative disease will be made through new and novel computational methods and statistical analyses to assess the immense interstudy data that is rapidly accumulating. An observational study is planned with cohorts of at least 250 subjects diagnosed with mild and moderate stage AD for stool collection and blood sampling analysis. Preparation of DNA libraries from stool samples for shotgun metagenomic sequencing, PCR amplification of 16S rRNA, and unbiased mass spectrometry-based metabolomics of blood samples are robust methods to determine microbial and protein composition from complex, human-derived media [39–42]. The study is proposed to be adequately powered (unpublished data, Elmaleh 2020), utilizing detectable effect sizes from the human microbiome project [43] for relative abundances among rare and common taxa. Although currently in early stages of progress, the study will broadly expand the ability to determine AD staging as it relates to gene-specific microbial and human metabolite profiles. It remains very hopeful that the pooling of such data generated, its creative analysis, and collaboration from many research groups will uncover the inter-related variables most associated with neurodegenerative processes, such as human metabolomic profiling of prodromal AD, that occur before and during the development of AD.
Figure 1 provides an overview for an approach that allows for targeted, chronic titrations of the gut microbiome based upon a specific neurodegenerative disease profile. An individual’s profile is generated to connect the extent of symptomatic decline through cognitive testing, such as global Clinical Dementia Rating-sum of boxes (CDR-SOB) and Mini-Mental State Examination (MMSE), to biological markers observed in circulating metabolites and constituents of an individual’s fecal microbiome. With a large enough cohort, cross-comparisons of individual profiles can be made to identify commonalities among specific neuroactive metabolites and microbiota to diagnose and treat early-stage dementia. In the proceeding sections, we will review the connections between aging, inflammation, the microbiome and neuroactive metabolites, and current work in this burgeoning field to highlight the potential use of chronic, low-dose antibiotics to treat neurodegenerative disease.
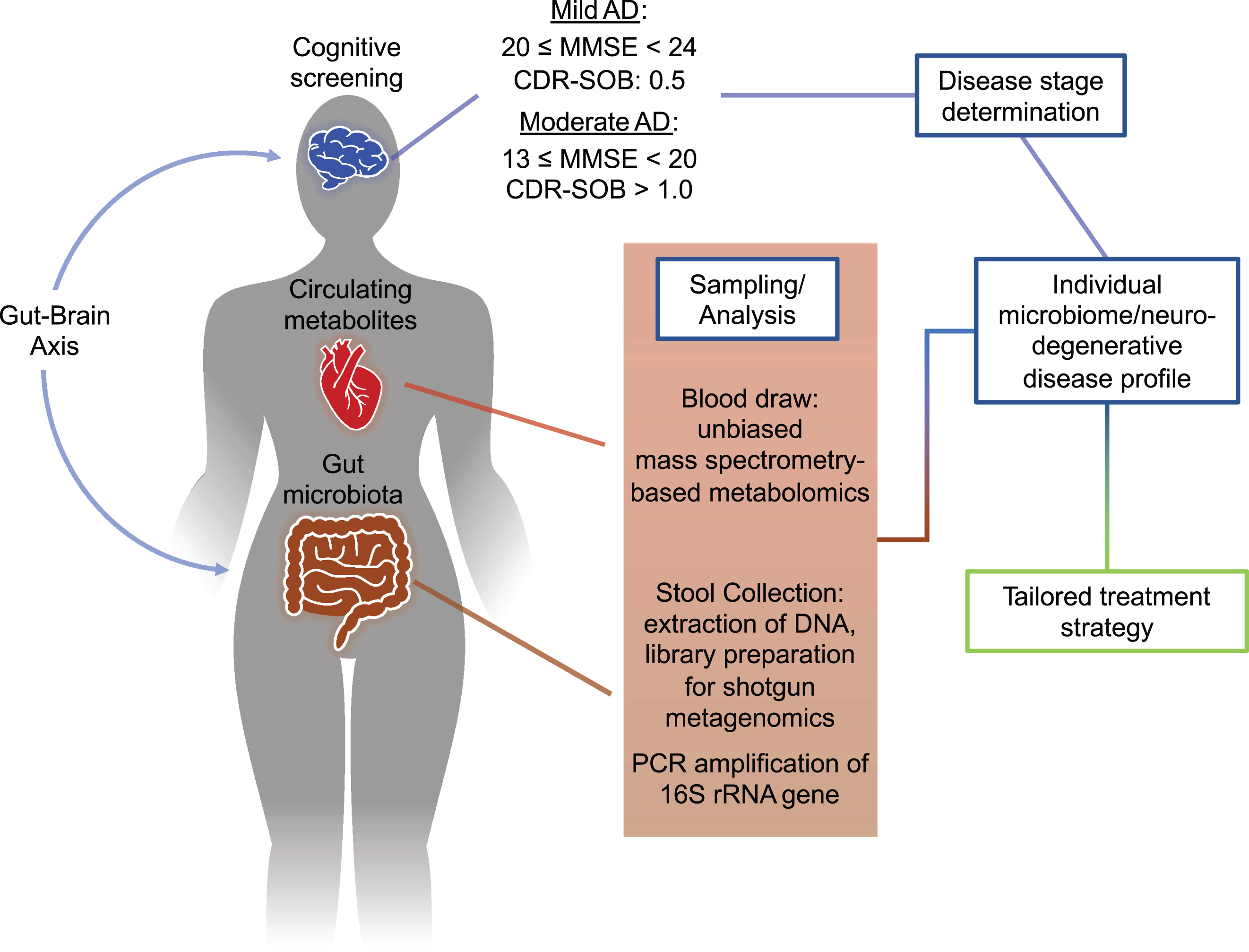
Safe, chronic titrations of bacterial metabolites as treatment for neurodegenerative disease. Intercommunication along the gut-brain axis by pathogenic resident microbes in the gut are able to influence the progression of neurodegenerative processes. Multi-omic analysis of circulating blood metabolites and feces, coupled with dementia screening, can develop an individual’s neurodegenerative profile at their respective stage of disease. This approach, when married with a large sample set, may uncover specific microbial species and metabolite levels relevant to a particular stage of cognitive decline. Using the generated profile, a tailored combination treatment plan can be constructed that addresses an individual’s presentation of disease. MMSE, Mini-Mental State Examination; CDR-SOB, Clinical Dementia Rating –sum of boxes.
Aging, microbiome, and inflammation
The body’s CNS is protected from exposure to peripheral media very selectively through the blood-brain barrier (BBB). Considering AD as a cumulative result of neuroinflammation-induced neurodegeneration is bolstered by the increase in permeability of the BBB as we age. Magnetic resonance imaging experiments have shown that age-related BBB deterioration begins in the hippocampus and worsens with mild cognitive impairment and in AD [44]. Numerous entities have been associated with BBB permeability including Aβ vascular deposition with decreased expression of tight junction claudin-1 and claudin-5 proteins and increase in matrix metalloproteases MMP-2 and MMP-9 [45], neuron-derived perivascular tau protein [46], reactive oxygen species [47], and short chain fatty acids (SCFA) produced from gut microbiota [48].
A more permeable BBB could be associated with heighten immune responses in the CNS. Microglia provide immune surveillance within their microenvironment in the CNS and are essential to the maintenance and health of neurons. In the aging brain, microglia may be morphologically de-ramified and “primed” toward a pro-inflammatory state, as evidenced by increased MHC II receptor and mRNA expression of TNF-α, IL-6, and IL-1β, as well as anti-inflammatory cytokines IL-10 and TGFβ1 upon lipopolysaccharide (LPS) challenge [49, 50]. Interestingly, LPS administration in aged mice increased expression of IL-10 in primed microglia but did not result in decreased production of pro-inflammatory IL-1β [50], a result that may be explained by an age-associated decline with IL-10 and TGFβ signaling pathways within aging microglia and astrocytes [51]. Astrocytes normally support BBB endothelial cell integrity and are involved in the recruitment and restriction of immune cells from the periphery into the CNS through cytokine and chemokine secretion after injury [52]. Transgenic APOE mouse models of AD show that astrocyte-derived APOE4 involves the inflammatory cytokine cyclophilin A (CypA) to induce BBB dysfunction through activation of a CypA—NF-κB—MMP-9 signaling pathway that causes neurovascular deficits and may initiate neurodegeneration [53].
Rethinking AD as more than just a neurodegenerative disease has brought about efforts to further delineate pathological from commensal or symbiotic residents of the gut by identifying their physiological abundance and exploring the contributions to metabolism and how their dysfunction may play a role in pathogenesis. Fecal transplant studies in animals have shown that the microbiome can exhibit substantial neurocognitive effects after transplantation, even to some degree restoring cognitive impairment (Table 1).
Fecal transplantation in AD animal models
ABX, antibiotic cocktail including kanamycin, gentamicin, colistin, metronidazole, and vancomycin; ADLPAPT, AD-like pathology with amyloid and neurofibrillary tangles; APP/PS1, amyloid precursor protein/presenilin-1; BBB, blood-brain barrier; CLDN1, claudin 1; dB, decibels; GFAP, glial fibrillar acidic protein; Iba1, ionized calcium-binding adaptor molecule; MWM, Morris water maze; ORT, object recognition test; PSD-95, postsynaptic density protein 95; SAMP8, senescence-accelerated mouse prone 8; SAMR1, senescence-accelerated mouse resistant 1; SD, Sprague-Dawley; WT, wildtype; XAN, xanthoceraside; ZO-1, tight junction protein 1.
The composition of the healthy human gut is highly variable, with thousands of possible bacterial species represented and potentially many more due to the individual diet and geographical environment, genetics, and immune response [54]. The population and density of gut flora is spatially diverse along the GI tract, which is likely dependent on the pH, oxygen partial pressures, and digestive role of a particular compartment, as well as commensal and symbiotic relationships with the host [55]. A large study of fecal samples from 1,000, age and sex stratified, healthy individuals analyzed by 16S rRNA gene sequencing found that species diversity is largely dominated by the two phyla Firmicutes and Bacteroidetes, and that gut microbiota composition is significantly determined by an individual’s environment and demographics [56]. Table 2 includes some of the most abundant taxa along the GI tract.
Prevalent taxa along the gastrointestinal tract
CFU/mL, colony-forming units per milliliter. Compiled from references [55, 177–181].
In the case of many acute pathological infections, it is tempting to find and accuse a specific microbe as having a causative, aberrant role in any one of these conditions, and understandably so, as the bulk of the microbiome is made up of bacteria, and a lesser extent fungi and parasites, many of which have known and negative connotations in regards to human health [57–59]. These detrimental attributes may be due in part to over expression of would-be normally commensal bacteria relative to others in the gut that can trigger robust inflammatory responses. The relative population of a mucin-degrading microbe, Ruminococcus gnavus, can increase in Crohn’s disease flare ups, secreting a glucorharmnan polysaccharide that induces significant TNF-α release from dendritic cells through direct activation of the toll-like receptor TLR4 [60]. A fecal microbiome metagenomics study of 20 ALS patients and 20 cognitively normal individuals discovered an increase in Bacteroidetes and decrease in Firmicutes abundance compared to controls [41]. Another study comparing fecal microbiome of 25 AD patients to those of 25 cognitively normal, age-matched individuals identified a relative increase in Bacteroidetes and decreased Firmicutes and Bifidobacterium in AD [61]. A transgenic Parkinson’s disease (PD) mouse study found the presence of gut microbiota was a prerequisite for motor deficits and the promotion of inflammatory microglia activation in an α-synuclein dependent manner, highlighting the interdependence of gut microbes and the neurodegenerative processes in the brain [62]. Interestingly, distinct gut microbiota alterations in Blautia, Akkermansia, Escherichia-Shigella, and Desulfovibrio in APP/PS1 AD model mice preceded onset of amyloid deposition and microglia-related neuroinflammation [63], suggesting further that neurodegenerative processes could be influenced by activity within the gut and early biomarker analysis may have diagnostic potential for disease later in life.
Additionally, the inter-individual variability of microbiome populations can be extensive among inflammatory disease states as well as among healthy individuals. Inflammatory bowel disease exhibits significant heterogeneity with dominant microbiota varying from individual to individual but can be of similar diversity levels for an individual regardless of inflammatory status, a factor that has been attributed to epigenetic factors like host DNA methylation and gene expression [64]. The gut microbiome collectively functions like another organ, aiding in digestion, elimination of harmful microbes, and the critical maintenance of the gut mucosa that are all essential for proper immune function [65]. The chronic dysregulation of microbiota populations, or dysbiosis, can subsequently impact the body’s ability to fight off infections without initiating a larger scale inflammatory response, which has been attributed to inflammatory bowel disease and chronic low-grade systemic inflammation [65]. Shotgun metagenomics analysis of fecal samples of the extremely old (e.g., over 105 years of age) in comparison to other, younger age groups have increased expression of genes involving xenobiotic degradation and altered carbohydrate and lipid metabolism, suggesting that healthy aging may be related to the body’s regulation of gut microbiota in response to environmental pollution stressors and dietary habits [42]. Advanced aging is often accompanied by increased weight loss and malnutrition, both of which are increased risk factors for developing AD [66]. Methanobrevibacter smithii is the dominant Euryarchaeon constituent of the human gut microbiota and is known to affect polysaccharide digestion by consuming bacterial fermentation end products, increasing the energy utility of ingested food [67]. Populations of methanogenic archaea have higher relative diversity in the gut microbiome of older individuals [68]. Anorexic individuals have increased M. smithii abundance compared to normal healthy adults [69] likely due to the body promoting M. smithii survival. Increased gut populations of methanogenic archaea are associated with increased adiposity, weight gain, and obesity [70], all of which are related to higher serum levels of pro-inflammatory biomarkers [71]. This systemic inflammation may further trigger primed microglia in progressive disorders like AD-related dementia, developing over decades and presenting differently for every person, and has subsequently made the identification of particular microbes that are directly related to generalized dementia inherently challenging.
The microbiome’s role in key neurochemical and neurometabolic pathways associated with stress and progressive neurodegenerative disorders is emerging. Microbial community sequencing approaches, notably 16S rRNA gene sequencing of fecal samples, have been used to identify altered relative microbe population distributions in the gut with several inflammatory conditions including inflammatory bowel disease [72], irritable bowel syndrome [73, 74]; as well as neurodegenerative and psychiatric disorders including Parkinson’s disease [75], AD [61, 76], autism spectrum disorder [77–79], schizophrenia [80], and major depressive disorder [81]. It is clear that peripheral microglia and macrophages are involved in the neurodegenerative process. However, these preliminary indication studies need a more substantive determination of the components of the microbiome genetic profiles that modify the CNS and increase or decrease neuroinflammation. It cannot be overstated that the volume of data that emerges from these kinds of studies are immense and careful methodology for its analysis is required. In our planned study, we intend to profile the gut microbial community in early AD subjects with CDR global score of 0.5 and in subjects with AD onset and correlate them with other human metabolic information to uncover specific lines of taxa and gene functions within the microbiome that is associated with both early and moderate disease stages. In contrast to other studies, this study will incorporate at least 250 subjects to allow a significant statistical difference for a more accurate indication of the specific AD staging profile. The identification of such a profile should lead to a tailored treatment design that would be based on the microbial and/or human metabolic profile.
Neuroactive metabolites of the gut
Gut microbiota are in many ways able to influence the CNS directly and indirectly, largely through the secretion of endotoxin and production of a range of metabolites (Table 3). Though some of the neuroactive metabolites provided in Table 3 may be endogenously produced by human metabolism or by microbiota, production of constituents of the tryptophan metabolic pathway is inherently dependent on dietary consumption of the essential amino acid tryptophan precursor that cannot be produced metabolically in humans. It is also of note that neuroactive catecholamines such as dopamine and norepinephrine are enzymatically produced from non-essential L-tyrosine [82]. Dopamine is produced in the cytosol in regions of the midbrain and forebrain and stored in synaptic vesicles until released into the synapse [82]. Dopamine is converted to norepinephrine by dopamine β-hydroxylase with cofactor ascorbate [82]. Both dopamine and norepinephrine, along with neurotransmitters serotonin and γ-aminobutyric acid, can also be produced by gut microbiota to influence host behavior and physiology [83].
Neuroactive metabolites produced by gut microbiota and other sources
AhR, aryl hydrocarbon receptor; FFAR3, free fatty acid receptor 3; IAA, indole acetic acid; IL-1β, interleukin 1 beta; MMSE, Mini-Mental State Exam; NCFB, National Collection of Food Bacteria; NCIMB, National Collection of Industrial, Food and Marine Bacteria; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; p38MAPK, p38 mitogen-activated protein kinase; SCFA, short chain fatty acid; ROS, reactive oxygen species; SOD, superoxide dismutase.
The cellular envelope of most gram-negative bacteria is lined with endotoxin, also known as LPS, that contributes to bacterium structural stability and membrane vesicle trafficking and can be excreted into the local cellular environment through bacterial outer membrane vesicles and also upon rupture of the bacterium cell wall [84]. The toxicity of LPS is thought to be due to the exaggerated host immune response to LPS rather than any inherent toxicity. The outermost domain of the LPS is composed of a glycan polymer O-antigen moiety that is specific for a given bacterium and is what the immune system develops antibodies against. The LPS inner domain composed of the relatively conserved Lipid A moiety is responsible for the toxicity associated with LPS and is recognized by the immune system through the formation of the TLR4-MD-2 heterodimer complex, although efficient activation of the complex is dependent on the number of lipid chains and conservation of both phosphate groups within Lipid A [85]. TLR4 is expressed by many cells of the immune system and TLR4 activation by LPS promotes the secretion of many pro-inflammatory cytokines [86] through a diverse set of potential signaling pathways including activation of NF-κB, MAP kinase, and IRF5 [87].
LPS can not only activate pro-inflammatory signaling pathways in the gut but also in the CNS [88]. Murine experiments show BBB permeability of LPS alone to be quite low and LPS breaches likely occur through non-BBB, peripheral LPS receptors [89]. It is not yet known exactly what mechanism LPS uptake into the CNS occurs, but experiments with primary human neuronal-glia co-cultures show that LPS accumulates within neuron parenchyma and impairs neuronal transcription products such as NfL more so than astroglia products such as glial fibrillary acidic protein (GFAP) [90], suggesting that LPS may interrupt neuronal homeostatic structural maintenance and post-injury repair processes, contributing to neurodegeneration transcriptionally as well as through neuroinflammation. Interestingly, heightened systemic LPS levels have been found to promote cognitive deficits in C57BL/6J mice by activating NF-κB signaling pathway to induce neuroinflammation as evidenced by increased TNF-α, IL-1β, prostaglandin, and nitric oxide [91], and increased retinal neurodegeneration in predisposed P23H rats [92]. Peripheral administration of a single dose of LPS in mice experienced increased TNF-α levels in the brain that was sustained over 10 months despite serum and liver levels reverting to baseline [93]. Experiments with organotypic hippocampal slice cultures from C57BL/6Babr mice show synaptophysin levels, a protein ubiquitously expressed in synapses, drop upon LPS administration, a process that is directly mediated by microglia and inflammatory cytokine IL-1β [94]. Cytokine treatment of IL-4, IL-6, IL-13, IL-1β, TNF-α, and IFN-γ to a variety of cell types have been associated with deterioration of endothelial cell tight junction proteins [95], suggesting that LPS may breach the BBB under chronic, systemic inflammatory conditions to further promote neuroinflammation. Porphyromonas gingivalis oral exposure to AβPP transgenic mice experienced increased amyloid deposition in hippocampus and cortex, heightened brain levels of pro-inflammatory cytokines TNF-α and IL-1β, and increased levels of LPS in the brain and serum, suggesting that periodontitis caused by P. gingivalis may induce neuroinflammation that brings about pathologic mechanisms related to AD [96]. It is clear that exposure to bacterial LPS induces a wide range of cascading effects that can be detrimental to neuronal function long-term.
Dysregulated tryptophan metabolism by the gut microbiome
Tryptophan is an essential amino acid whose metabolic pathway is dysregulated in AD, often associated with kynurenine arm of the tryptophan metabolic pathway. Decreasing circulating levels of tryptophan are shown to worsen cognitive decline of AD patients [97]. This is potentially related to increased immune activation with increasing age, leading to increased inflammation, upregulation of kynurenine through indoleamine 2,3-dioxygenase (IDO) activity and as much as a 50%decrease in available tryptophan for serotonin production [98,99, 98,99]. Availability of tryptophan could also be due to the ability of the resident microbiome to metabolize tryptophan. Indole-3-propionic acid (IPA) is a product of microbial tryptophan metabolism that induces the expression of tight junction proteins, such as occludins, and can inhibit NF-κB signaling to reduce production of pro-inflammatory cytokines TNF-α, IL-6, and IL-1β following LPS exposure [91].
Approximately 90%of the essential neurotransmitter serotonin in the human body is produced in the gut and many constituents of the gut microbiome are able to induce serotonin production by stimulating enterochromaffin cells in the intestine to promote gut homeostasis and regulate intestinal peristalsis [100]. Serotonin is important for cross-communication between the central and enteric nervous system and is regulated through, and aids in maintaining brain, nerve health, intestinal movements, and regulates behavioral effects such as emotion, stress response, and appetite [101]. As such, underproduction or lack of access to serotonin is associated with mood and depressive disorders, and the interplay between serotonergic system and cholinergic, dopaminergic, GABAergic system play important roles in memory retention and cognition [102].
Upregulation of the kynurenine metabolic pathway occurs in AD patients, with increased indoleamine 2, 3 dioxygenase (IDO) activity that also leads to increases in quinolinic acid in the hippocampus [103]. Quinolinic acid is an NMDA glutamate receptor agonist with high selectivity for the NR2B subunit (GRIN2B) in the forebrain, and high concentrations are associated with neuron excitotoxicity and may initiate disruption of neuronal cytoskeleton homeostatic mechanisms [104, 105]. Additionally, quinolinic acid stimulates lipid peroxidation in an Fe2 + dependent manner which may be abated by iron chelation therapy [104]. Intraperitoneal injection of E. coli LPS into C57Bl/6J mice increased the brain concentration of quinolinic acid approximately four-fold [106]. It is possible that bacterial secretions such as LPS originating in the gut that are potently pro-inflammatory could initiate a wide range of neurotoxic effects if the source of inflammation is sustained.
Challenges with analysis of the microbiome and current clinical trials
Despite the incredible amount of data that is produced, the appeal of identifying the functional proteome of gut microbiota (metaproteomics), the constituent taxa and gene functions of the gut microbiota (metagenomics), and the degree to which constituent genes are expressed (metatranscriptomics) is the potential to holistically relate the functional behavior of the gut microbiome and its production of metabolites (metabolomics) to host health, importantly with regards to neurodegenerative disease progression [107]. High resolution mass spectrometry-based quantitative metabolomics platforms are commonly used to identify a biological sample’s metabolite composition [108]. Robust quantitative metaproteomic methods have been utilized to determine gut microbiome proteomes from fecal samples in colorectal cancer [109], but the complex nature of exploring the microbiome has brought about many challenges and implementation strategies with omics workflows that have been reviewed at length elsewhere [110–113]. Comparative investigations into the microbiome constituent differences via fecal samples is quickly becoming a robust platform to understand inter-individual variability [114]. Challenges associated with the collection of fecal samples to determine microbiota composition are numerous and the distribution observed can be method-of-collection and storage-dependent [77]. For example, approximately 99%of bacteria within the colon are obligate anaerobes and can be sensitive to exposure to oxygen [67]. Thus, the environment in which feces are collected, the kits used for feces collection, storage conditions, time lapsed from collection, and its analysis in atmosphere can all have downstream effects on what microbiota distributions are observed in addition to variances and specificity in genomic sequencing methods [115, 116]. Full-length 16S rRNA high-throughput sequencing may be eventually required instead of less computationally-intensive short-read approaches to sort polymorphisms in the bacterial 16S gene to identify specific strains of the same species [116]. We foresee this aspect may be critically important for some individuals as different strains of the same bacterial species can elicit widely inconsistent adaptive immune responses [117], potentially aiding in the identification of inflammatory and neurodegeneration-related microbiota.
Perhaps the most significant challenge with collecting vast amounts of multi-omics data is in its interpretation. Fortunately, this downside is well recognized, and many researchers have begun to implement strategies to draw out inter-disciplinary connections between human and bacterial metabolism and disease. The Virtual Metabolic Human (VMH) database is one such interactive utility that links information of myriads of known metabolites, metabolic reactions, human and microbial genes, Mendelian diseases, specific microbes, and foods [118]. Others have expanded the VMH concept for virtual reconstruction of whole-organ metabolism in relation to the host microbiome to potentially model healthy inter-organ and co-metabolism dynamics to better predict effective therapeutics for a particular individual based on sex, sex-specific and shared organ types, bodily fluid and dietary metabolites, and host/microbe genes [119]. These virtual models can also be tailored to a specific disease. Mechanistic information of how multiple upstream disorders, including interconnections between β-cell dysfunction and insulin resistance, could be elucidated with VMH that result in symptomatic diabetes, providing an exciting opportunity to explore combination treatment strategies [120]. Genome-scale metabolic modeling has also been applied to AD and found several metabolic pathways were downregulated, including folate metabolism and mitochondrial trafficking [121], the latter of which is strongly associated with late-onset AD [122].
It will also be critical to have a simple-to-follow and unambiguous collection protocol to reduce compliance errors, especially if AD patients were to be asked to perform the collection and the surrender of stool samples independently. Because the microbiome is so variable, obtaining an individual’s baseline composition is exceedingly difficult. This inherently demands large cohort studies to determine what the healthy baseline is across many people for adequate comparison to a diseased or imbalanced microbiome [123]. As such, multiple considerations need to be taken into account for the particularly challenging nature of microbiome molecular analyses, and many best-approaches are continuously evolving with ongoing research and findings. In addition to sample collection and storage, these challenges can include facets of experimental design, metadata collection, data generation and processing, and data analysis and statistics. Choices in microbial molecular assays used, as mentioned above, are also challenging as they have independent and complementary benefits, but multi-omic studies are becoming increasingly common [124]. For longitudinal comparison of future, large-participant studies of the microbiome, the standardization of collection and storage methods for stool samples as well as robust comparison of multivariable statistical tools will be critical to determine the metabolomic, metagenomic, and transcriptional differences that may relate to neurodegenerative disease progression. Multiple exploratory and interventional clinical trials are currently exploring connections between the gut microbiome and AD (Table 4).
Clinical Trials Exploring Gut Microbiome and/or Metabolic Profiling with AD in Humans
ADAS-cog, Alzheimer’s Disease Assessment Scale-cognitive subscale; APOE4, apolipoprotein E; AVLT, auditory learning verbal test; BBB, blood-brain barrier; CDR-FTLD, Clinical Dementia Rating frontotemporal lobar degeneration; CDR-SOB, Clinical Dementia Rating sum of boxes; CDR, Clinical Dementia Rating; CFU, colony forming units; EGCG, epigallocatechin gallate; IL, interleukin; MCI, mild cognitive impairment; MMSE, Mini-Mental State Exam; MNA, Mini-Nutritional Assessment; MoCA, Montreal Cognitive Assessment; MRI, magnetic resonance imaging; NfL, neurofilament light; NINDS/ADRDA, National Institute of Neurological and Communicative Diseases and Stroke/Alzheimer’s Disease and Related Disorders Association; PCR, polymerase chain reaction; PD, Parkinson’s disease; RT-qPCR, reverse transcription quantitative polymerase chain reaction; SCFA, short chain fatty acid; TNF-α, tumor necrosis factor alpha.
The results of additional clinical trials exploring the effect of dietary supplements that have the capacity to modulate gut dysbiosis may provide further insight for improvement of gut-related cognitive decline. An ultimately underpowered randomized, double-blind controlled trial with the multinutrient Souvenaid found no significant impact on cognitive performance in prodromal AD patients as evaluated by a neuropsychological test battery over 24 months, but the observed cognitive decline post-trial was much improved over estimates [125–127]. Interestingly, increased plasma levels of docosahexaenoic acid (DHA), a brain-abundant omega-3 fatty acid ingredient in Souvenaid, are reported to increase gut microbiome alpha diversity that is linked to decreased gut inflammation [128] but phosphatidylcholine DHA, and not DHA, was associated with a reduced risk of developing dementia [129, 130]. The hormone melatonin commonly used as a dietary supplement as a sleep aid is an antioxidant whose production in the brain is reduced with aging and in AD [131]. Melatonin can increase gut microbiota Firmicutes:Bacteroidetes ratio and goblet cell differentiation in a TLR4-dependent manner in mice, and has been associated with decreasing Aβ-induced neuroinflammation in vivo. Resveratrol is an orally available polyphenol derived from many common foods and reduces amyloid plaque formation in Tg19959 AD mice model [133]. Resveratrol is well-tolerated in humans and was demonstrated to reduce pro-inflammatory MMP9 in CSF [134, 135] and has been found to modulate gut microbiota populations in various animal models as well as in humans [136]. Other dietary supplements associated with modulation of the gut microbiome and neuroinflammation include curcumin [137], alpha-tocopherol (vitamin E) [138, 139], and vitamin D [140, 141].
Required cohort sizes
As seen in Table 4, most microbiome cohorts studied to date were relatively small, and in most cases include only a few dozen patients. This may have limited discoveries. For example, in recent work [142], we showed that by profiling both the microbiome and serum metabolite levels of 491 healthy participants, we were able to apply machine-learning algorithms to predict metabolite levels in held-out individuals on the basis of their gut microbiome composition. We obtained statistically significant predictions for hundreds of metabolites, in some cases explaining more than 50%of the observed variance in the metabolite levels. We further validated these microbiome-related predictions by showing a high replication rate in two geographically independent cohorts from the U.K. and Denmark that were not available to us when we trained the algorithms. These data also allowed us to reveal specific bacterial interactions with metabolites. Unlike in other settings, using standard statistical power analysis to compute the required cohort size in advance may be challenging, as we need assumptions on the size of the effect that various microbiome parameters will exhibit in order to apply the statistical power calculation methodologies. Given the large interpersonal variability that exists in the microbiome composition across people, we thus believe that cohort sizes of several hundreds of individuals are needed in order to unravel novel and robust associations.
Antibiotic resistance concerns with long-term antibiotic use
Because peripheral dysbiosis in the gut can influence microglial activation in the CNS and lead to neurodegenerative processes, chronic administration of low dose, specific antibiotics to titrate access and influence of proinflammatory bacteria or fungi may prove useful for slowing down or inhibiting progressive neurodegenerative disease without inducing further dysbiosis. Elucidating the inherent anti-inflammatory behavior of an antibiotic outside of its bacteriostatic or bactericidal qualities may prove beneficial as a dual-action treatment option for neurodegenerative disease (Table 5).
Antibiotics with Anti-Inflammatory Effects
CCI, controlled cortical impact; CD, cluster of differentiation; DCFDA, 2’,7’–dichlorofluorescin diacetate; ECP, eosinophilic cationic protein; GFAP, glial fibrillary acidic protein; HLMVEC, human lung microvascular endothelial cells; Iba-1, Ionized calcium binding adaptor molecule 1; IL, interleukin; LPS, lipopolysaccharide; MMP-9, matrix metalloproteinase 9; MPO, myeloperoxidase; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; SD, Sprague-Dawley; SOD, superoxide dismutase; THP-1, Tamm-Horsfall Protein 1 human monocyte cell line; TNF-α, tumor necrosis factor alpha.
Antibiotics are primarily developed to control or eliminate pathogenic overgrowth of bacteria responsible for an acute infection. Two main concerns are associated with such an approach; over-prescription of antibiotics have contributed to the rise in antibiotic-resistant mutations in bacteria, making the treatment of infections more complex and heightening the demand for new antibiotics that are effective against drug-resistant bacteria [143]. Further, prolonged chronic administration of antibiotics is associated with major gut and peripheral side effects [144, 145]. Depending on the type of infection and specific antibiotic indicated, antibiotics are usually prescribed for treatment courses lasting 3–10 days at doses ranging from 300 mg/day to approximately 2,000 mg/day [146]. Standard antibiotic regimens can alter the host microbiome long-term. A one-year study of the administration of a weeklong course of ciprofloxacin or clindamycin in humans impacted species diversity in feces for one and four months, respectively; however, a weeklong course of minocycline resumed to normal diversity after one month and amoxicillin had no significant change from baseline over one year [147]. Recent studies have explored the ability of low-dose antibiotic administration for chronic infections. Low-dose antibiotic regimen (50 mg nitrofurantoin, 100 mg trimethoprim, 250 mg cefalexin) taken orally once daily for 12 months reduced the number of urinary tract infections (UTI) in individuals that use self-catheterization to empty their bladder [148]. However, there was also an overall increase in antibiotic resistance to commonly-prescribed antibiotics in bacterial urinary isolates [148]. The authors propose the observed antibiotic resistance and concurrent reduction in symptomatic UTIs could be due to changes in host response to chronic low-dose antibiotic treatment that promotes growth of colonizing bacteria and opposes virulent growth [148]. These are promising results for the pursual of chronic low-dose antibiotic treatments if populations of commensals can be maintained to discourage colonization of others but further study warrants exploring cumulative low-dose antibiotic regimens for use with specific microbiome profiles to expand effectiveness and lower drug resistance and toxicity from chronic administration.
Use of antibiotics for treatment of neurological disease
Antibiotic treatment is widely known to affect the populations of the gut microbiome and murine experiments have explored the effect that broad spectrum antibiotics can have in inducing gut dysbiosis and impact gut-brain signaling. A C57BL/6N mice study found through an 11-day oral gavage treatment of a high-dose antibiotic cocktail (including bacitracin, neomycin, ampicillin, meropenem, and vancomycin) was sufficient to impair object recognition memory associated with expression of tight junction proteins TJP1, claudin-5, and occludin in the hippocampus and amygdala [149]. Interestingly, even though serum concentrations of cytokines IL-1β, IL-6, IL-10, TNF-α, and IFN-γ did not differ between control and antibiotic-treated groups despite treatment with antibiotics substantially decreasing bacterial load, species diversity, and production of SFCAs; neural-signaling molecules were affected including BDNF reductions in medial prefrontal cortex, hypothalamus, and hippocampus, and increases of NMDA receptor GRIN2B and serotonin transporter SLC6A4 in amygdala [149].
One way gut dysbiosis is suspected to influence the CNS is through communication via the vagus nerve. Experiments with male, 8-week-old (adult) C57BL/6 mice subjected to chronic social defeat stress (CSDS) with CD-1 aggressor mice found that mice, after 14-day antibiotic treatment (ampicillin, neomycin sulfate, metronidazole) in ad libitum drinking water, given fecal transplants from CSDS-susceptible mice upregulated IL-6 in plasma, downregulated expression of synaptic proteins GluA1 and PSD-95 in prefrontal cortex, and Lactobacillus intestinalis and Lactobacillus reuteri were associated with depressive behavior in a vagus nerve-dependent manner [150].
Altering the gut microbiome with high-dose antibiotics may deleteriously affect neurotrophic factors, but the targeting of a particular gut microbe may have benefits. Though limited by a small sample set, one study found by eliminating Helicobacter pylori, a bacterial species more abundant in AD, in the gut with a 1-week cocktail of clarithromycin (500 mg bid), amoxicillin (1,000 mg bid), and omeprazole (20 mg bid), followed by daily 20 mg dose of omeprazole for 1 month, gradually improved cognitive function two years following treatment [151]. Although the current way antibiotics are prescribed to treat acute infections and curb microbial dysbiosis may not have the same benefit in treating neurocognitive decline, there is justification to explore the possible concurrent beneficial effects of antibiotic treatment of neurodegenerative disorders.
But like with any drug, dosing matters. Reducing inflammation due to dysbiosis in the gut through the use of antibiotics may still be an alternative approach to treating neurodegenerative disorders. Pivotal experiments in APP/PS1 mice models of early-onset AD have shown that chronic, broad-spectrum antibiotic treatment regimens (from age 2 weeks to 6 months) are able to reduce amyloid deposition and increase soluble Aβ levels, alter microglial morphology surrounding plaques, and alter gut microbiota composition with marked increase in phyla Firmicutes (family Lachnospiraceae) and Verrucomicrobia (genus Akkermansia) [152]. Akkermansia mucinophila is a commensal gram-negative eubacterium that is positively correlated to gut barrier integrity and gut mucosal thickness, particularly in obesity and metabolic disorders [153]. Its abundance has been inversely related to histopathological inflammation in the human bowel [154]. Prolonged absence of complex polysaccharides and fiber derived from plants in the diet have been associated with decreased substrate for commensal, SCFA-producing microbiota and higher abundances of mucin-degrading microbes, including Akkermansia mucinophila; potentially leading to increased permeability of the gut mucosal layer, increased interaction of gut luminal contents with the epithelium, and subsequent increases in inflammation [155]. Further extensive experiments with APPPS1-21 mice exposed to a long-term antibiotic cocktail (including kanamycin, gentamicin, colistin, metronidazole, and vancomycin) observed a general decrease in Aβ pathology and morphological alterations in microglia that were present in only male mice [156].
The use of tetracycline class of antibiotics for potential treatment of neurodegenerative disorders has been investigated. In vitro experiments with low-dose doxycycline were shown to promote α-synuclein oligomerization into off-pathway, non-toxic oligomeric structures [157]. Though no effect on astrogliosis was observed, intraperitoneal doxycycline administration to Huntington’s disease R6/2 model mice exhibited decreased microglial activation and enhanced neuronal survival [158]. Anti-amyloid properties of doxycycline are also seen with Aβ aggregates [159]. Molecular dynamics simulations show that doxycycline binds the hydrophobic C-terminal residues of Aβ42 to prevent further fibril stabilization [160]. Low-dose doxycycline has also been shown to decrease LPS-induced inflammatory cytokine expression of TNF-α, IL-6, and IL-8 in vitro. Long-term treatment of doxycycline (up to 200 mg/day) and/or rifampicin (up to 300 mg/day) over 1 year provided no significant improvement in cognitive decline in patients with mild to moderate AD, though the effect on the individuals’ microbiome, or on Aβ or tau serum, or in CSF levels is unknown as the study only assessed changes in cognition, daily function, and behavior [162]. It is possible that issues with clinical treatment of AD with doxycycline reside within treating more-progressed stages of disease and that the potential anti-amyloid and anti-inflammatory benefits seen in in vitro experiments may translate better to prodromal patients if doxycycline were to be used as an AD prophylactic [159]. In a large, randomized Phase IIb clinical trial of two-year treatment with the tetracycline antibiotic minocycline with patients with mild AD (determined only by standardized Mini-Mental State Exam (sMMSE)≥24), neither the 400 mg/day or the 200 mg/day treatment group slowed or improved cognitive decline over control as measured by the sMMSE or Bristol Activities of Daily Living Scale, with the higher dose of minocycline poorly tolerated long-term [163]. It is possible, because no biomarker analyses were performed to strengthen an AD diagnosis for inclusion in the trial, there may have been trial participants without AD diagnosed as such.
The cephalosporin antibiotic ceftriaxone is reported to exhibit neuroprotective effects by increasing expression of the glutamate transporter GLT-1 in various in vivo models of AD, PD, and cerebral ischemia; as well as reduction of pro-inflammatory mediators, such as TNF-α, IL-1β, IFN-γ, and NF-κB in animal models of PD, cerebral ischemia, and traumatic brain injury [164, 165]. Many analogues of sulfonamide antibiotics exhibit free radical scavenging activity in vitro, including some Aβ fibril and oligomer inhibition properties [176]. The 4-quinolone scaffold of quinolone antibiotics shows antioxidant activity against lipid peroxidation as well as hydroxyl radical production [166], although fluoroquinolone use has been associated with increased production of reactive oxygen species [167] and rare, but lasting, serious adverse effects including tendon rupture and nerve damage [168, 169].
It is unsurprising that diet has a large effect on the microbiome as its overall composition and proliferation is determined in no small part by what we eat, but the effect of diet on antibiotic efficiency in microbiota populations is emerging. In C57BL/6J mice, administration of amoxicillin decreased cecum glucose levels and significantly expanded the relative abundance of the phylum Bacteroidetes, specifically B. thetaiotaomicron and B. vulgatus, in harvested cecum compared to controls [170]. Interestingly, this change was attributed to amoxicillin, but not ciprofloxacin, treatment affording a metabolically-favorable environment for Bacteroides species. Further, by increasing the mice’s dietary glucose, amoxicillin’s ability to promote B. thetaiotaomicron amounts relative to control were diminished. In other experiments where C57BL/6J mice were fed a Western-style (high-fat, high-sugar) diet, large increases were found in phyla Bacteroidetes and Verrucomicrobia and a decrease in Firmicutes relative to vehicle [171]. Upon ciprofloxacin treatment an expansion of Firmicutes was only observed in the Western diet with no significant change with normal diet, potentially related to the diet-dependent ciprofloxacin-induced transcriptional changes in mucin substrate utilization and metabolic processes [171]. Collectively these studies suggest that diet and the metabolic environment in the gut may play a role in how effective some antibiotics are in modifying microbiome composition.
Indeed, administration of antibiotics to treat a neurodegenerative disorder may benefit with an accompanying dietary regimen prescription, of sorts, to stabilize aberrant populations to support healthy commensal growth and discourage opportunistic harmful growth. However, many more omics studies with large sample sets will be required to understand the complex interplay between antibiotics-induced host and microbial gene expression, sustained inflammatory processes and metabolite release, and the extent to which diet intervention will be efficacious in the long term. Unfortunately, there are few results for antibiotic treatment for neurodegenerative diseases in humans. This more than likely stems from the difficulty in containing and relating exogenous differentiating factors, such as diet, exercise, genetics, age, and medications, to the progression of disease. Future experiments will necessarily require targeted biomarker panels that would include direly-needed information on diagnosable ranges of pro-inflammatory mediators associated with AD progression, in concert with an analysis of gut microbiome dysbiosis present at early, potentially treatable stages of AD. The main approaches included in this personalized treatment plan are included in Fig. 2.
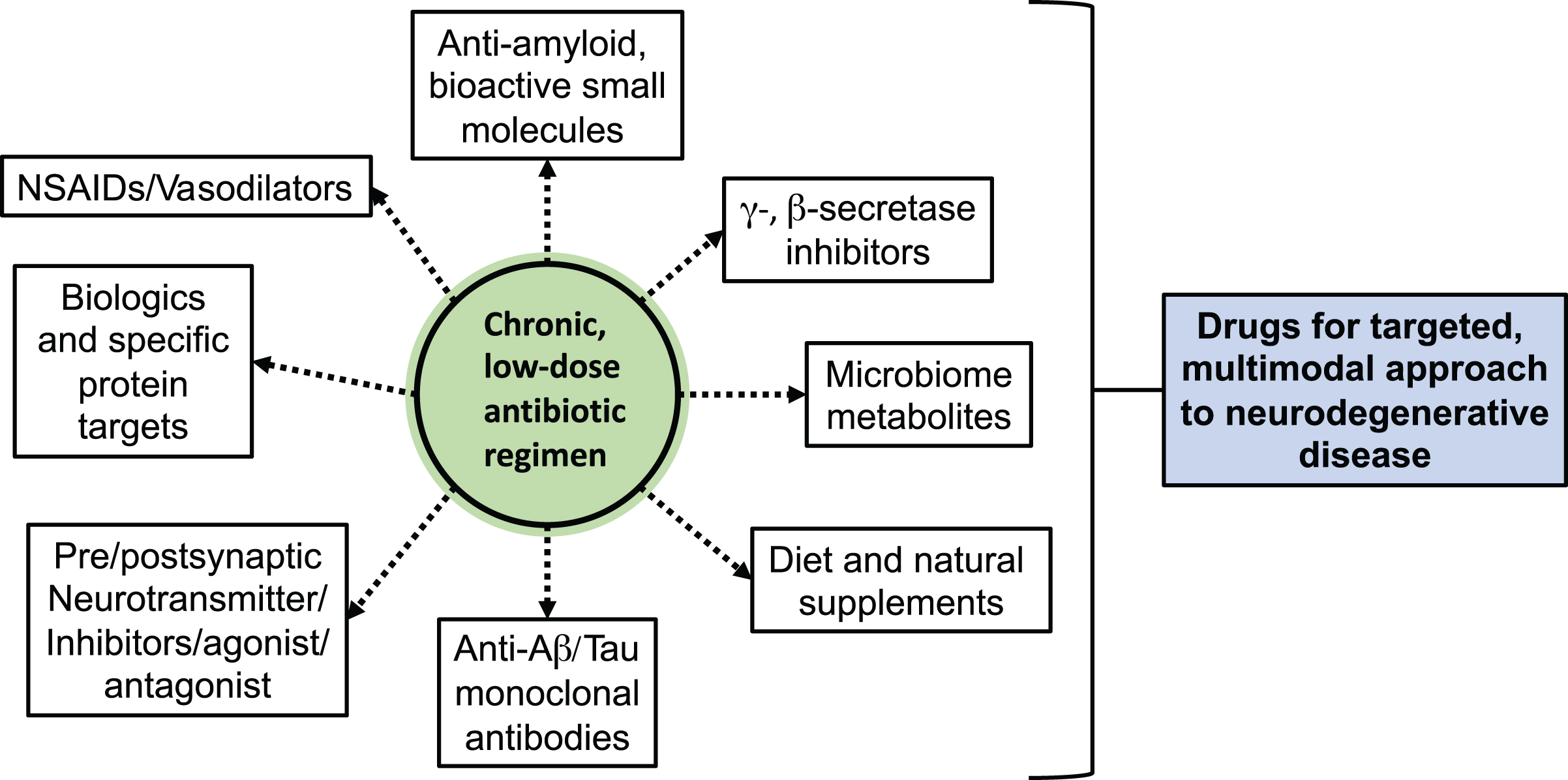
Main approaches for personalized combination treatment of low-dose antibiotics with other neurodegenerative disease-modifying or -ameliorating drugs.
CONCLUSION
The effect of the gut microbiome and other infections on CNS degeneration is supported by a large body of evidence of animal studies and relatively-limited substantive efficacy studies in humans. Many approaches for altering gut microbiota propose supplementing deficient gut microbiota with probiotic supplements, diet, metabolites, antibiotics, and fecal transfer from normal healthy subjects to sick subjects. Unfortunately, these proposed modes of treatment have demonstrated correlation to cognitive decline in a limited number of participants. However, a systematic profiling and staging of an AD profile coupled with a tailored treatment approach has yet to be demonstrated for efficacy and safety of chronic administration in controlled, randomized clinical trials. Here we emphasized the ability of slowing early AD by enhancing peripheral microglia recruitment to the CNS as a disease-modulating approach for profiling disease staging that intuits designing a specific treatment to titrate harmful bacteria and fungi access in early disease. We propose an expansive trial to establish the fecal microbial composition and serum metabolic profile of 500 patients diagnosed with mild to moderate AD to discover species and metabolic connections related to the severity of AD. The ambitious scope of this kind of study will be appropriately powered to assist with early diagnosis of AD as it relates to the microbiome. These efforts will contribute tremendously to the development of future probiotic or antibiotic treatments that have the potential to address the complex, multi-faceted nature of an individual’s neuroinflammatory progression of AD, with the intent of eventually ameliorating its downstream cognitive effects by titrating the gut microbiome of harmful constituents and metabolic irregularities.