
Abstract
The cell surface level of apolipoprotein E receptor 2 (ApoER2) increases by cyclic transport of ApoER2 and then activates Reelin signaling pathway to exert neuroprotective function in AD. ApoER2 ligand Apolipoprotein E4 (ApoE4) inhibits the recycling of ApoER2 to the cell surface rendering neurons unresponsive to Reelin. Carnosic acid (CA) is proven to possess neuroprotective and neurotrophic functions in Alzheimer’s disease (AD) mouse model. However, there are few reports about how ApoE4 impairs the recycling of ApoER2 and if CA can affect the cyclic transport of ApoER2. In this study, we demonstrated that ApoE4 attenuates the binding of sorting nexin 17 (SNX17) to ApoER2 and inhibits the recycling of ApoER2, resulting in decreased cell surface level of ApoER2. Further, we found that CA enhances the binding of SNX17 to ApoER2, counteracts the negative effects of ApoE4 on the cell surface level of ApoER2 to reverse the ApoE4-induced reduction in Reelin signaling activation by increasing the phosphorylation of the N-methyl-D-aspartate receptor (NMDAR) and cAMP-response element-binding protein (CREB) and the expression of Gria2. Thus, CA promotes neurite growth inhibited by ApoE4. Our work suggests that CA may be a potential approach to attenuate the risk of ApoE4-associated AD.
Keywords
INTRODUCTION
Apolipoprotein E receptor 2 (ApoER2) is a member of the Apolipoprotein E (ApoE) receptor family initially identified as a transmembrane receptor to bind apolipoproteins in order to transport lipoprotein particles [1]. In addition to apolipoproteins [2], ApoER2 was reported to bind other developmental proteins such as Reelin [3]. By binding Reelin, cell surface ApoER2 activates the phosphorylation of Dab1, leading to the phosphorylation and activation of NMDA receptors to increase long-term potentiation (LTP) [4]. NMDA receptors activated by the Reelin-ApoER2 signaling pathway counteract NMDA receptor-dependent synaptic suppression and LTP suppression induced by amyloid-β protein (Aβ) [4, 5]. Therefore, Reelin and cell surface ApoER2 levels play an important role in the pathological process of Alzheimer’s disease (AD).
ApoE4 most strongly influences AD risk and age of onset. Clinical studies and amyloid mouse models have shown that ApoE4 increases the risk of AD by accelerating and initiating the accumulation, aggregation, and deposition of Aβ in the brain [1]. In addition, ApoE4 is one of the strongest competitors blocking ApoER2 from binding to Reelin [6]. ApoE4 also reduces cell surface ApoER2 by inhibiting the recycling of ApoER2 [7]. There are few reports on how ApoE4 blocks the recycling of ApoER2 to the cell membrane surface. It is known that inhibiting Na+/H+ exchangers on endocytic membranes by specific inhibitors may increase intracellular vesicle acidity, then help restore ApoER2 trafficking in the presence of ApoE4 [8]. However, it has not been reported if ApoE4 involves in the ApoER2 endocytic trafficking pathway. SNX17, which resides in the early endosome membrane, promotes the recycling of ApoER2 by directly binding with ApoER2 and facilitates the transportation of ApoER2 back to the plasma membrane [9–11]. Therefore, there is a reason to speculate that ApoE4 may reduce the level of ApoER2 on the membrane by interfering with the interaction of SNX17 with ApoER2.
Carnosic acid (CA), a polyphenolic diterpene isolated from the herb rosemary (Rosmarinus officinalis), has multiple neuroprotective and neurotrophic properties including prevention of neurotoxin-induced neuronal cell death [12], promotion of neurite outgrowth in neurons [13], and the protection against Aβ-induced injury in vitro and in vivo [14, 15]. In addition, CA activates the Nrf/ARE signaling pathway to improve learning and memory in an AD mouse model, increases dendrites and synaptic markers, and reduces Aβ plaques and phosphorylated tau staining in the hippocampus of AD mouse models. [16]. However, it has not been reported if CA can affect cyclic transport of neuronal membrane surface receptors such as ApoER2.
In this study, we demonstrated that ApoE4 inhibited the recycling of ApoER2 to the cell membrane by blocking the binding of SNX17 to ApoER2. Further, CA induced cell surface accumulation of ApoER2 by enhancing the interaction of SNX17 with ApoER2, followed by phosphorylation of NMDAR and CREB and finally promoted neurite growth through the CREB-target gene Gria2. Importantly, we found that CA counteracted ApoE4-induced reduction of ApoER2 membrane accumulation and inhibited the Reelin pathway and neurite growth. Taken together, our results suggest that CA may be a potential approach to attenuate the risk of ApoE4-associated AD.
MATERIALS AND METHODS
Reagents and antibodies
CA (>95% purity) was obtained from Biobiopa (Yunnan, China). ApoE4 was purchased from PeproTech (Rock Hill, NJ, USA). The antibodies used were as follows: rabbit anti-ApoER2 (Cat. No: ab108208), mouse anti-ApoER2 (Cat. No: ab58216), and rabbit anti-NMDAR2B (Cat. No: ab183942) from Abcam (Cambridge, MA, USA); mouse anti-SNX17 (Cat. No: sc-166957) and mouse anti-Dab1 (Cat. No: sc-271136) from Santa Cruz Biotechnology (Santa Cruz, CA, USA); rabbit anti-CREB (Cat. No: 9197 S) /P-CREB (Cat. No: 9198 S), rabbit anti-NMDAR2A (Cat. No: 4205 S) /P-NMDAR2A (Cat. No: 4206 S), and mouse anti-phosphotyrosine (Cat. No: 9411) from Cell Signaling Technology (Beverly, MA, USA); secondary anti-mouse and anti-rabbit antibodies conjugated to Alexa Fluor 488 or 598 from Abcam (Cambridge, MA, USA).
Cell culture
PC12 cells were purchased from ATCC. The PC12 cells were cultured in DMEM (Dulbecco’s modified Eagle’s medium, DMEM, Biological Industries, Israel) with 1% penicillin-streptomycin (Thermo Fisher Scientific, MA, USA) and 10% FBS (fetal bovine serum, FBS, Biological Industries) at 37°C with 5% CO2 and saturated humidity.
Neuronal cultures
Pregnant Sprague-Dawley rats were acquired from the animal center at Shandong University. Hippocampal neurons were extracted and cultured as previously described [17, 18]. In brief, the hippocampus was dissected from embryos at embryonic day 18 and digested with 0.25% EDTA/trypsin at 37°C for 30 min before transferring to F12/DMEM medium containing 10% fetal bovine serum. Neurobasal medium containing 2% B27 (Thermo Fisher Scientific, MA, USA), 0.5 mM glutamine (Thermo Fisher Scientific, MA, USA) and 10% penicillin-streptomycin (Thermo Fisher Scientific, MA, USA) was used for culturing neurons at 37°C with 5% CO2 and saturated humidity.
Cell surface biotinylation
Cells were incubated with 0.3 mg/mL EZ-Link Sulfo-NHS-SS-Biotin (Thermo Fisher Scientific, MA, USA) and lysed using RIPA lysate (Beyotime Biotechnology, Jiangsu, China). Streptavidin beads (Thermo Fisher Scientific) bound to biotinylation surface proteins were incubated with the cell lysate overnight at 4°C. The streptavidin-biotinylation surface protein complex was boiled for 5 min in SDS-PAGE loading buffer (CWBiotech, Beijing, China). The protein samples were then loaded onto SDS-PAGE gels for electrophoresis and detected by western blotting with ApoER2 antibody.
Immunofluorescence analysis
The cells were fixed, permeabilized, and blocked. ApoER2 (Abcam, Cat. No: ab58216) or SNX17 antibodies was used for incubating cells at 4°C overnight, respectively. Cells were then incubated for 1 h at room temperature using Alexa Fluor 488 conjugated or Alexa Fluor 594 conjugated secondary antibody, respectively. Images were captured with a Zeiss LSM780 confocal microscope (Microstructural Platform, Shandong University). The number of ApoER2 and SNX17 aggregates was assessed by quantifying points of overlap of red and green fluorescence. Fluorescence intensity was analyzed using ImageJ software (>50 cells per data set). Specifically, a straight line was drawn onto a magnified image and the red and green fluorescence intensities on the line were measured. Fluorescence intensity values were then plotted graphically using GraphPad Prism software.
Immunofluorescence analysis of membrane surface ApoER2
A fluorescence-based antibody binding assay was used for analyzing surface receptors described elsewhere [19, 20]. Briefly, cells were fixed, blocked, and then incubated with ApoER2 antibody (Abcam, Cat. No: ab58216, corresponding to amino acids 83-171 of Human ApoER2). Goat anti-mouse IgG H&L (Alexa Fluor 594) (Abcam, Cat. No: ab150116) was used for incubating cells. Then, the cell slides were permeabilized and blocked. ApoER2 antibody was used for incubating cells and then incubated with goat anti-mouse IgG H&L (Alexa Fluor 488) (Abcam, Cat. No: ab150113). Images were captured using a Zeiss LSM780 confocal microscope. Fluorescence intensity was analyzed using Image-Pro Plus 6.0 software (>50 cells per data set). Briefly, the grey value on the axon was measured and converted to the absolute expression value.
Western blotting
The procedure for western blotting is described in detail in our previous publication [21]. Briefly, cells were lysed in RIPA lysis buffer. Protein samples were then electrophoresed, transferred, blocked, and incubated with primary antibodies. The membranes were then incubated with peroxidase-coupled anti-mouse/rabbit IgG. Finally, the membranes were visualized using ECL (Millipore). Images of membranes were obtained using BioImaging Systems (UVP Inc., Upland, CA, USA).
NMDA receptor and Dab1 phosphorylation assays
Cells were lysed using RIPA lysis buffer. The protein supernatant was then transferred to a fresh EP tube and rotated overnight with an anti-phosphotyrosine antibody (Cat. No: 9411, Cell Signaling Technology) at 4°C. Immune complexes were precipitated by incubation with 30μL Protein A/G Magnetic Beads (MCE, NJ, USA) at 4°C for 2 h. Precipitated proteins were analyzed by western blotting using anti-Dab1, NMDA2A, or NMDA2B antibodies.
Immunoprecipitation
Cells were lysed in RIPA lysis buffer. The protein supernatant was subsequently rotated overnight at 4°C with anti-ApoER2 or anti-SNX17 antibody. Immune complexes were precipitated by incubation with 30μL Protein A/G Magnetic beads at 4°C for 2 h. Precipitated proteins were analyzed by western blotting with anti-SNX17 or anti-ApoER2 antibody.
Molecular simulation docking
The Discovery Studio software was used to assay the possible binding regions of CA with ApoER2 and SNX17. The crystal structure of the ApoER2 LA12 domain (PDB accession number 5B4Y) and the SNX17 PX domain (PDB accession number 3LUI) were acquired from the PDB database. The CA molecular structure was drawn using Discover Studio 4.1 software. The Discovery Studio software was then used to simulate how the respective domains of ApoER2 (5B4Y) and SNX17 (3LUI) may interact with CA.
RNA-seq analysis
The RNAs were isolated from PC12 cells treating with CA/APOE4 and control cells using Trizol Reagent. The libraries were obtained from the Illumina TruSeq1 RNA Sample Preparation. The samples were sequenced on an Illumina HiSeq 2500 Sequencing system. The TopHat2 software was used to map to the genome assembly. The HTSeq software was used to quantify the gene-level expression. The Deseq2 software was used to obtain differential expression.
Statistical analysis
The Student’s t-test or one-way ANOVA was used to assess statistical significance. Results are presented as mean±S.D., and p < 0.05 was considered significant.
RESULTS
CA increases cell surface level of ApoER2 to counteract the inhibitory effect of ApoE4
In AD, the surface level of ApoER2 has better resistance to Aβ via the Reelin-ApoER2-NMDAR pathway [4, 5]. In addition, CA can also resist Aβ damage and reduce Aβ plaques in the AD mouse model [16]. Therefore, we wanted to explore the relationship between CA and ApoER2 surface expression. PC12 cells were treated with 10μM CA for 0.5 and 2 h. Although no significant difference was found in the total expression of ApoER2 (Fig. 1A, B), surface biotinylation analyses showed that the cell surface level of ApoER2 was significantly elevated after CA treatment (Fig. 1A, C).
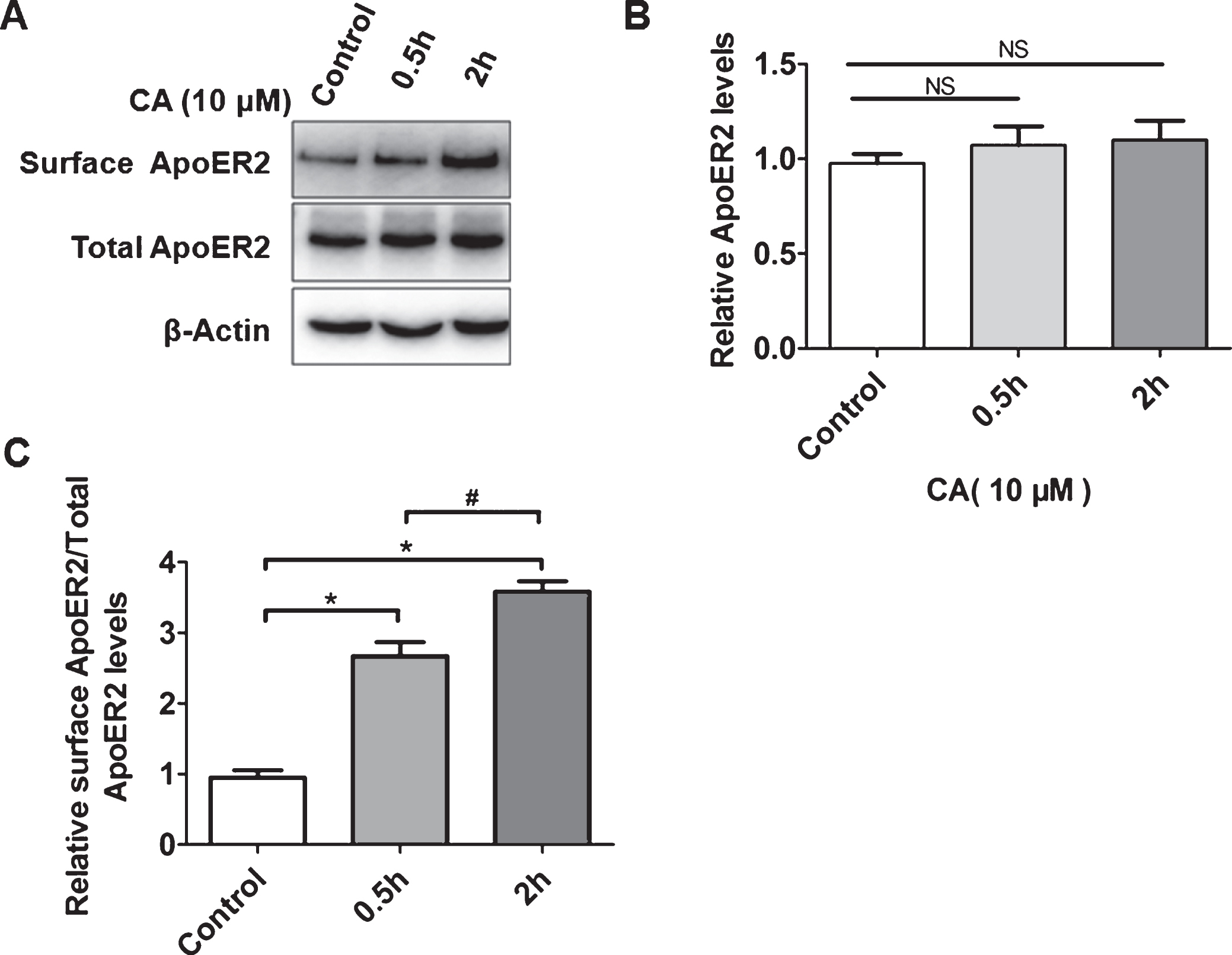
Carnosic acid (CA) promotes the accumulation of ApoER2 on the cell surface. PC12 cells were incubated with CA (10μM) for 0.5 h and 2 h, and either lysed and analyzed for total cellular ApoER2 level by western blotting or subjected to surface biotinylation before lysis, followed by affinity precipitation and analysis of biotinylated proteins by western blotting for cell surface ApoER2 (A). Bands from Total ApoER2 in A were quantified by ImageJ software (B) (n = 5; NS = no significance, 0.5 h or 2h versus control; Students t-test). Bands from surface ApoER2 in A were quantified by ImageJ software (C) (n = 5; *p<0.05, 0.5 h or 2 h versus control; #p<0.05, 2 h versus control; Student’s t-test).
As ApoE4 has been reported to inhibit the recycling of ApoER2 to the cell surface [7], we next analyzed whether CA could reverse the suppressive effect of ApoE4. PC12 cells and hippocampal neurons were incubated for 2 h with CA (10μM), ApoE4 (3μg/mL), or a combination of both. As previously reported, ApoE4 reduced the cell surface level of ApoER2, and this effect was reversed by CA treatment in both PC12 cells and primary hippocampal neurons (Fig. 2A, B). Immunofluorescence analyses in PC12 cells and hippocampal neurons confirmed that CA reversed the reduction in cell surface ApoER2 levels induced by ApoE4 (Fig. 2C-F).
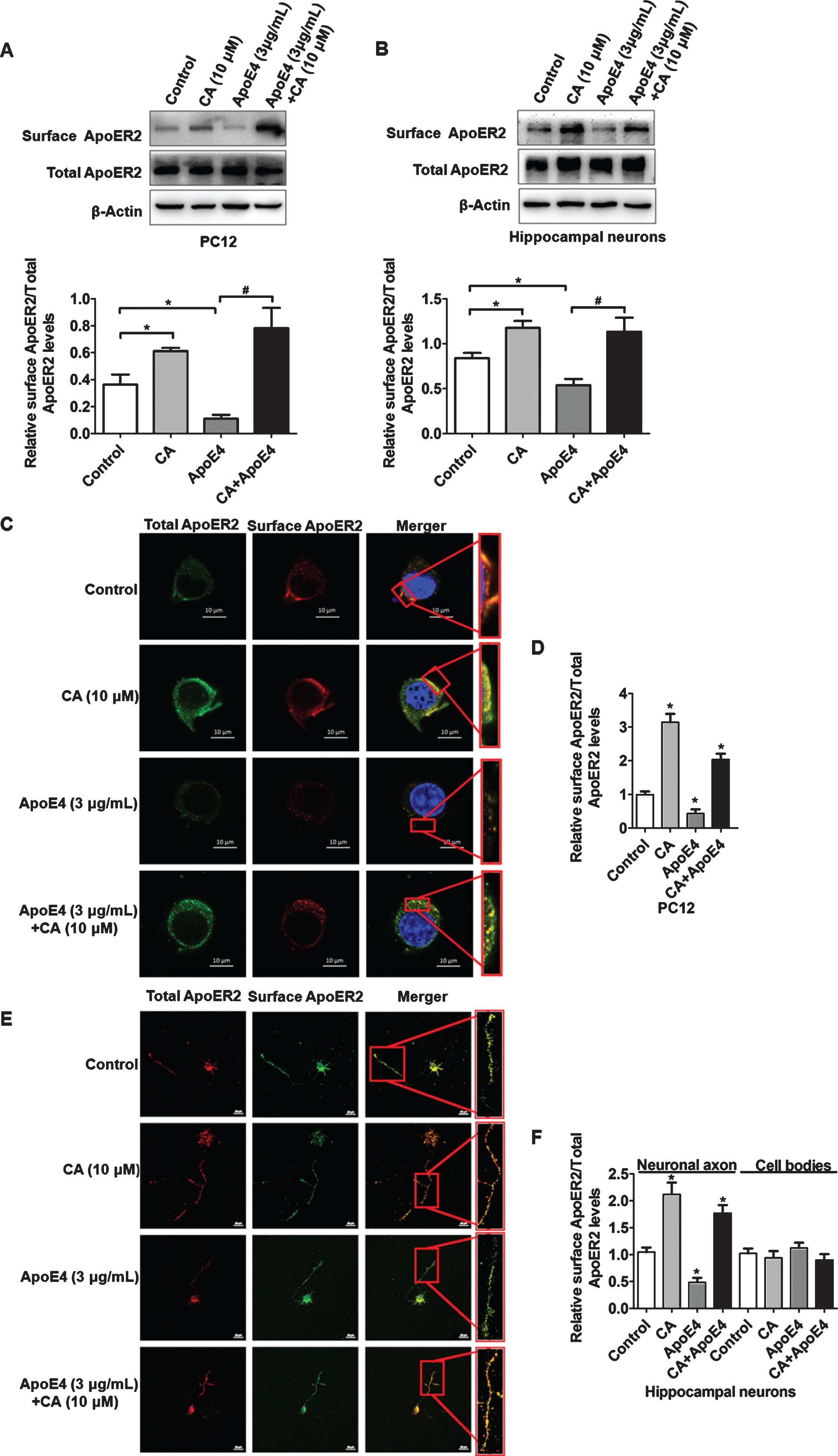
Carnosic acid (CA) reverses the inhibitory effect of ApoE4 on cell surface ApoER2. PC12 cells (A) or hippocampal neurons (B) were treated as indicated and were subjected to surface biotinylation. After lysis and affinity precipitation, biotinylated proteins were analyzed by western blotting for surface ApoER2. Bands from surface ApoER2 in A and B were quantified by ImageJ software (n = 4; *p<0.05, CA or ApoE4 versus control; #p<0.05, CA and ApoE4 versus ApoE4; Students t-test). PC12 cells (C) or hippocampal neurons (E) were incubated with CA or ApoE4 as indicated. Surface (red) and total ApoER2 (green) were detected by immunofluorescence. The surface expression of ApoER2 was mainly observed in the neuronal axon, rather than on cell bodies (E). Relative surface levels of ApoER2 to total ApoER2 were quantified and calculated (D, F) (n = 3; *p<0.05; Students t-test). Scale bars: 10μm in (C) and 20μm in (E).
CA increases cell surface level of ApoER2 by enhancing the interaction between SNX17 and ApoER2 inhibited by ApoE4
We then try to clarify how CA counteract the inhibitory effect of ApoE4 on the cell surface level of ApoER2. It has been reported that ApoE4 sequesters ApoER2 into intracellular recycling compartment [7] and SNX17 binds to endocytic ApoER2 to promotes its cell surface accumulation [9]. Based on these reports, we investigated whether ApoE4 affects the interaction of SNX17 with ApoER2. Immunoprecipitation and subsequent western blotting in PC12 cells revealed that ApoE4 inhibited the interaction between SNX17 and ApoER2 in a time-dependent manner (Fig. 3A, B). We then tested if CA may increase the cell surface level of ApoER2 by reversing the inhibitory effect of ApoE4 on the interaction. As we found CA enhanced the interaction between SNX17 and ApoER2 in PC12 cells (Fig. 3 C, D).
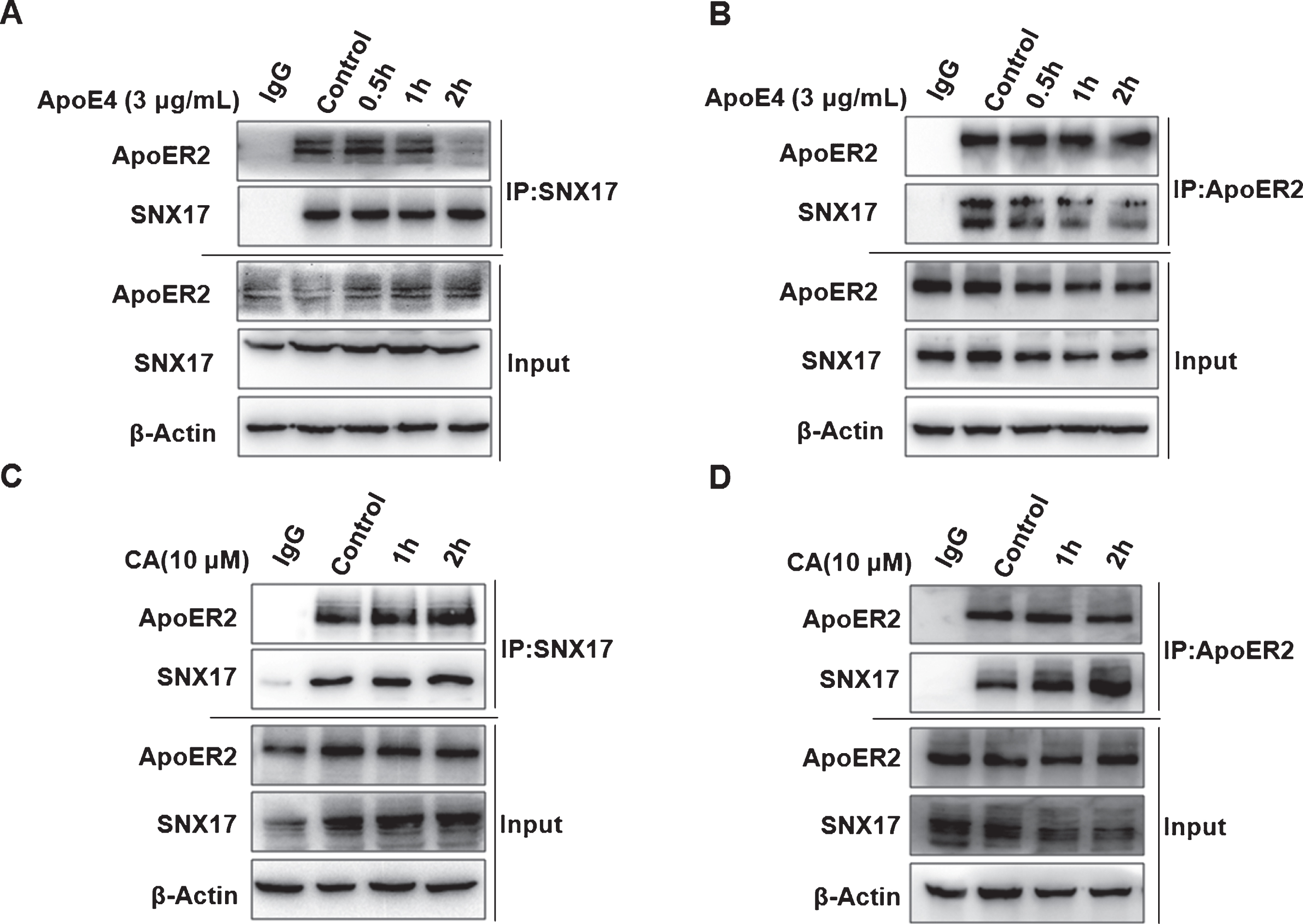
Carnosic acid (CA) and ApoE4 have opposing effects on the interaction of SNX17 with ApoER2. PC12 cells were treated with ApoE4 (3μg/mL) for 0.5 h, 1 h, and 2 h. Co-immunoprecipitation was performed using SNX17 antibody, and western blotting was used to detect the binding ability of SNX17 to ApoER2 (A). Co-immunoprecipitation was performed using ApoER2 antibody, and western blotting was tested for the binding ability of ApoER2 to SNX17 (B); PC12 cells were treated with CA (10μM) for 1 h and 2 h. Co-immunoprecipitation was performed using SNX17 antibody, and western blotting was used to detect the binding ability of SNX17 to ApoER2 (C). Co-immunoprecipitation was performed using ApoER2 antibody, and western blotting was used to detect the binding ability of ApoER2 to SNX17 (D). The above experiment was repeated three times independently.
We then treated PC12 cells with CA (10μM), ApoE4 (3μg/mL), or a combination of both. Subsequent immunoprecipitation and western blotting demonstrated that, in addition to increasing the binding of SNX17 with ApoER2, CA reversed the ApoE4-induced inhibition of the interaction between SNX17 and ApoER2 (Fig. 4A). The same result was observed in cultured primary hippocampal neurons (Fig. 4B). In addition, double staining of SNX17 and ApoER2 by immunofluorescence in PC12 cells and hippocampal neurons showed that ApoE4 suppressed co-localization of SNX17 and ApoER2, whereas CA significantly restored the co-localization (Fig. 4C-F, Supplementary Figure 1).
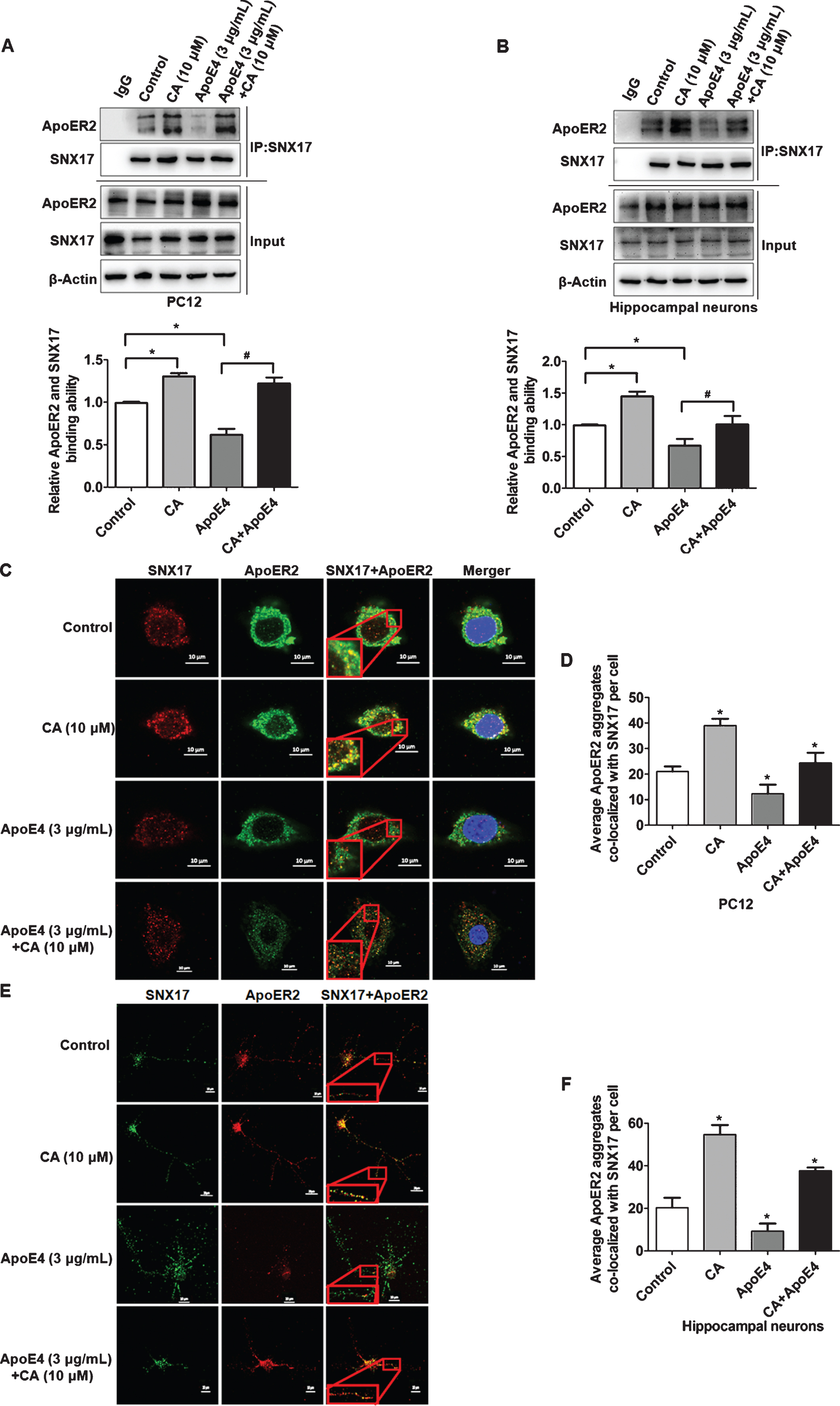
Carnosic acid (CA) promotes the interaction of SNX17 with ApoER2 and reverses the inhibitory effect of ApoE4. PC12 cells and hippocampal neurons were treated with CA (10μM; 2h) or ApoE4 (3μg/mL; 2 h) alone and together. Co-immunoprecipitation was performed using SNX17 antibody, and western blotting was used to detect the binding ability of SNX17 to ApoER2. The binding ability of SNX17 to ApoER2 was quantified by measuring the strips in PC12 cells (A) or hippocampal neurons (B) by ImageJ software (n = 5; *p<0.05, CA or ApoE4 versus control; # p < 0.05, CA and ApoE4 versus ApoE4; Students t-test). Cultured PC12 cells (C) and hippocampal neurons (E) were treated as indicated, followed by immunofluorescence staining for SNX17 (red) and ApoER2 (green). The nuclei were stained with DAPI (blue). Co-localization of SNX17 and ApoER2 (yellow) is indicated by the inserted frames in the magnified image. The average number of ApoER2 aggregates co-localizing with SNX17 in PC12 cells (D) or hippocampal neurons (F) were calculated by ImageJ software (n = 3; *p<0.05, CA, ApoE4, and CA and ApoE4 versus control; Student’s t-test). Scale bars: 10μm in the inserted frames of C and E.
As the interaction between SNX17 and ApoER2 promotes the recycling of ApoER2 and facilitates the transportation of ApoER2 back to the plasma membrane [9–11], our results indicated that CA increases cell surface level of ApoER2 by inhibiting the effect of ApoE4 on the interaction between SNX17 and ApoER2.
CA activates Reelin signaling pathway suppressed by ApoE4
The Reelin signaling pathway is mediated by cell surface ApoER2. ApoE4 has been reported to decrease the phosphorylation of the Reelin signaling downstream targets NMDAR and CREB by sequestering ApoER2 into the intracellular compartments [7]. Now that we have proved CA promotes cell surface ApoER2 at the presence of ApoE4, we next want to investigate whether CA could reverse ApoE4-induced inhibition of NMDAR and CREB phosphorylation. We first tested the effect of CA on the cytosolic adaptor protein Disabled-1 (Dab1), which is also known to be phosphorylated in response to Reelin signaling. We found that CA increased the phosphorylation levels of Dab1 (Fig. 5A, B). PC12 cells were then incubated for 2 h with CA (10μM), ApoE4 (3μg/mL), or a combination of both. Western blotting revealed that CA reversed the reduced phosphorylation of NMDAR (Fig. 5 C) and CREB(S133) (Fig. 5D) induced by ApoE4. A similar result was observed in cultured primary hippocampal neurons (Fig. 5E, F). These results indicated that CA could activate Reelin signaling pathway suppressed by ApoE4.
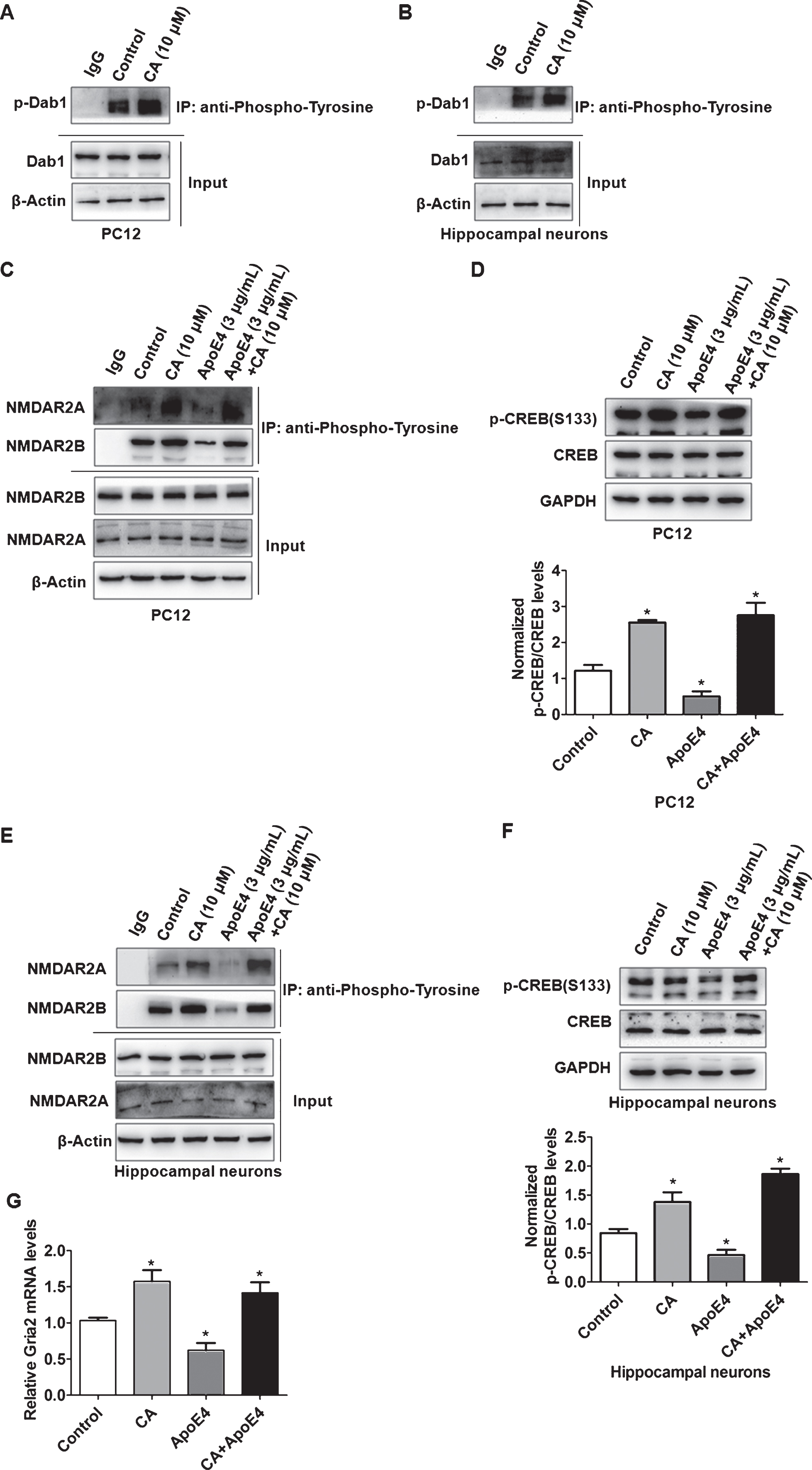
Carnosic acid (CA) increases the phosphorylation of the Reelin signaling pathway proteins NMDAR and CREB and reverses the inhibitory effect of ApoE4. PC12 (A) and Hippocampal neurons (B) were treated with CA. followed by co-immunoprecipitating with tyrosine-phosphorylated antibody and western blotting with Dab1 antibody. PC12 cells or hippocampal neurons were incubated with CA (10μM; 2 h), ApoE4 (3μg/mL; 2 h), or a combination of both. PC12 cells (C) and hippocampal neuronal (E) protein lysates were co-immunoprecipitated with tyrosine-phosphorylated antibodies, and western blotting was used to detect NMDAR phosphorylation levels using NMDAR2A and NMDAR2B antibodies (The co-immunoprecipitation experiment was repeated independently three times). The PC12 cells (D) or hippocampal neurons (F) were treated as above, and the expression of CERB and P-CERB (S133) was detected by western blotting. CERB and P-CERB bands were quantified using ImageJ software (n = 5; *p<0.05, CA, ApoE4, and CA and ApoE4 versus control; Students t-test). RT-qPCR confirmed the Gria2 expression in CA, ApoE4, and CA and ApoE4 treated PC12 cells (G) (n = 5; *p<0.05, CA, ApoE4, and CA and ApoE4 versus control; Student’s t-test).
We then further tried to identify the downstream effector of the Reelin pathway affected by CA and ApoE4. As CREB, a transcription factor regulates a variety of genes involved in Reelin that affect neuronal development and function [22], we performed transcriptome sequencing in PC12 cells. Differentially expressed genes between ApoE4 and CA treated PC12 cells were analyzed (Supplementary Figure 2). We found 45 downregulated genes in the ApoE4 group, 74 upregulated genes in the CA group compared with control (Supplementary Tables 1 and 2). Comparing these differential genes with the CREB gene database [23], we found Gria2 (glutamate ionotropic receptor AMPA type subunit 2), as a common target gene. Further real-time quantitative PCR results confirmed that CA increased Gria2 expression levels and reversed the inhibition of Gria2 expression by ApoE4 (Fig. 5G).
CA promotes neurite outgrowth and reverses the inhibitory effects of ApoE4
We have confirmed that CA increased the cell surface level of ApoER2 and enhanced the Reelin-NMDAR-CREB signaling pathway to induce Gria2 expression in the presence of ApoE4. As Gria2, a subunit of the AMPA receptor, was reported to express in the presynaptic axonal growth cones of neurons [24] and regulate neuronal growth [25]. We then explored the effects of CA on the regulation of neurite growth. Cultured hippocampal neurons were treated with ApoE4 (3μg/mL), CA (10μM), or a combination of both (Fig. 6A). Immunofluorescence analysis confirmed that CA promoted, while ApoE4 inhibited neurite growth. Furthermore, CA treatment reversed the inhibitory effect of ApoE4 on neurite growth (Fig. 6B, C).
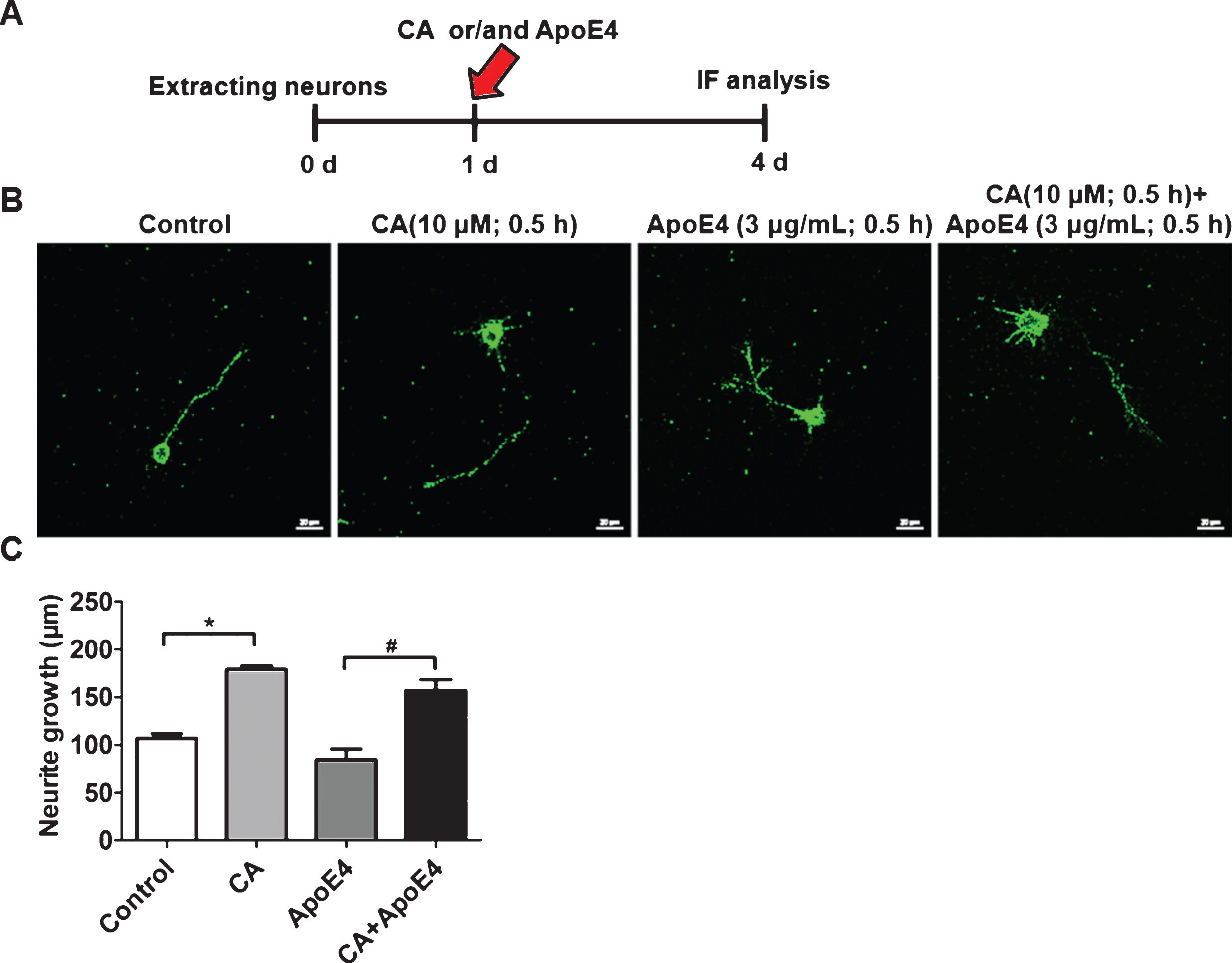
Carnosic acid (CA) promotes neuronal growth and reverses the inhibitory effect of ApoE4. Hippocampal neurons were extracted and cultured for 1 day, followed by pre-treatment with CA (10μM), ApoE4 (3μg/mL), or a combination of both for 0.5 h. Following the culture of hippocampal neurons for a further 3 days (A), the cells were fixed and analyzed by immunofluorescence (B) (Scale bars: 20μm). The normalized neuronal length was quantified and calculated using ImageJ software (C) (n = 3; *p<0.05, CA, ApoE4, and CA and ApoE4 versus control; Student’s t-test).
DISCUSSION
Carnosic acid (CA) has multiple neuroprotective and neurotrophic properties by protecting against Aβ-induced injury in vitro and in vivo [14], promoting dendrites and synaptic markers, and reducing Aβ plaques and tau phosphorylation staining [16]. Cell surface expression of ApoER2, which is an important component of the Reelin signaling pathway, is maintained by cyclic transport and plays important roles for resistance to Aβ-induced damage [4, 5]. However, the role of CA in cyclic transport of ApoER2 has not been previously reported. In this study, we demonstrated that CA increased cell surface accumulation of ApoER2 via enhancing the interaction of ApoER2 with SNX17, activated NMDAR and CREB in the Reelin signaling pathway, and promoted neurite growth, thus, reversed the inhibitory effects of ApoE4 on these functions (Fig. 7).
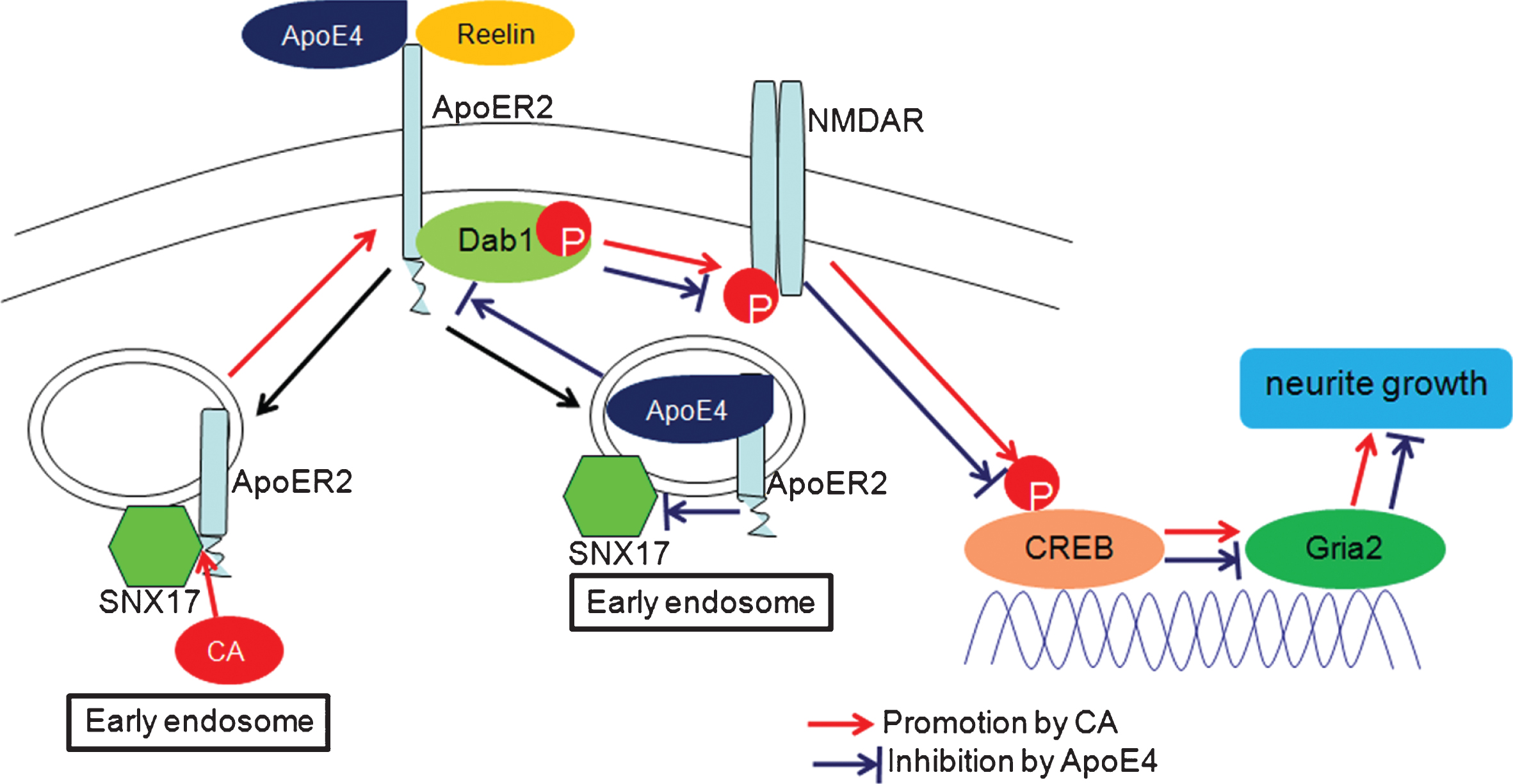
Schematic diagram depicting how carnosic acid (CA) reverses the negative effect of ApoE4. CA increased cell surface accumulation of ApoER2 via enhancing the interaction of ApoER2 with SNX17, activated NMDAR and CREB in the Reelin signaling pathway to promote neurite growth via increasing Gria2 expression, thus, reversed the inhibitory effects of ApoE4.
Endocytic trafficking of ApoE receptors (ApoER) plays an important role in maintaining ApoER function. By binding to the cytoplasmic domain of ApoER2, SNX17 directs ApoER2 from early endosomes to recycle it to the cell surface [11, 26–28]. In this study, we found that CA increases the cell surface level of ApoER2 in PC12 cells and primary hippocampal neurons by enhancing the interaction of SNX17 with ApoER2. To verify the function of CA in regulating the interaction of SNX17 with ApoER2, we constructed the 3D structures of CA (Supplementary Figure 3A). Therefore, we first identified the crystal structure of the ligand-binding LA12 domain of ApoER2 in the RCSB Protein Data Bank. When the LA12 domain of ApoER2 binds to a ligand, this promotes a conformational change from an open state to a closed state, which facilitates receptor recycling [29]. Through software simulation docking, we found that CA forms hydrogen bonds with Pro60 in the LA12 domain of ApoER2, suggesting that CA may promote conformational changes in the ligand-binding domain (Supplementary Figure 3B). We also used the RCSB Protein Data Bank to identify the crystal structure of the PX domain of SNX17, which directly binds to ApoER2 [9, 30]. Through software simulation docking, we found that CA forms a hydrogen bond with the surface Ser94 of the PX domain of SNX17. We, therefore, speculate that CA binds to the PX domain of SNX17, which in turn may affect the interaction of ApoER2 with SNX17 (Supplementary Figure 3 C).
The ApoER family is an evolutionarily highly conserved receptor family. In the brain, ApoE binds a variety of ApoERs to mediate lipid trafficking and metabolism, which are vital for the formation and maintenance of synapses [31]. ApoERs also play vital roles in synaptic modulation and signal transduction [32]. Reelin signaling is predominantly transmitted by the very low-density lipoprotein receptor (VLDLR) and ApoER2 [33, 34]. By binding to ApoER2, Reelin activates the cytosolic adaptor protein Disabled-1 (Dab1), which then activates Src family kinase (SFK). This leads to tyrosine phosphorylation of the NMDAR, resulting in increased CREB phosphorylation and enhancement of LTP [35, 36]. In our study, we found that CA increased the phosphorylation of Dab1 and restored the phosphorylation levels of NMDAR and CREB, which were reduced by ApoE4.
Furthermore, we identified Gria2 as a new downstream target of Reelin-NMDAR-CREB signaling pathway in mediating the function of CA. Expression of Gria2 was inhibited by ApoE4 and induced by CA. Finally, we confirmed that ApoE4 inhibited neurite growth and CA reversed the inhibition of neurite growth induced by ApoE4, which is consistent with the previous reports on the function of Gria2 [37–39].
In conclusion, our study revealed that CA increased cell surface accumulation of ApoER2 via enhancing the interaction of ApoER2 with SNX17, activated Reelin signaling pathway to promote neurite growth, reversed the inhibitory effects of ApoE4 on these functions (Fig. 7). Furthermore, our study suggests that ApoE4 may inhibit the ApoER2-NMDAR-CREB signaling pathway, thereby assisting the inhibition of synaptic and LTP by Aβ during the pathogenesis of AD. Therefore, our findings suggest that CA may be a potential approach to attenuate the risk of ApoE4-associated AD.