
Abstract
Mesenchymal stem cells (MSCs) promote functional recoveries in pathological experimental models of the central nervous system and are currently being tested in clinical trials for neurological disorders. However, no studies have examined the various roles of embryonic stem cell derived (ES)-MSCs in eliciting therapeutic effects for Alzheimer’s disease (AD). In the present study, we investigated the neuroprotective effect of ES-MSCs in cellular and animal models of AD, as well as the safety of the intra-arterial administration of ES-MSCs in an AD animal model. ES-MSCs displayed higher cell viability than that of bone marrow (BM)-MSCs in amyloid-β (Aβ)-induced cellular models. Moreover, the efficacy of autophagy induction in ES-MSCs was comparable to that of BM-MSCs; however, intracellular Aβ levels were more significantly reduced in ES-MSCs than in BM-MSCs. In a rat model of AD, ES-MSCs significantly inhibited Aβ-induced cell death in the hippocampus and promoted autophagolysosomal clearance of Aβ, which was concomitantly followed by decreased levels of Aβ in the hippocampus. Furthermore, ES-MSC treatment in Aβ-treated rats featured a higher memory performance than that of rats injected solely with Aβ. Finally, intra-arterial administration of an appropriate cell density of ES-MSCs was safe and free from in situ occlusion or cerebral ischemia. These data support the therapeutic potential of ES-MSCs and clinical applications of the intra-arterial route of ES-MSC administration in AD.
INTRODUCTION
Alzheimer’s disease (AD), the most common neurodegenerative disorder, is characterized pathologically by synapse loss and the presence of neuritic plaques and neurofibrillary tangles [1, 2]. Of the biochemical alterations that occur in multiple pathways in the AD brain, such as amyloid-β (Aβ) metabolism, tau phosphorylation, and lipid regulation, Aβ plays an important role in AD pathogenesis. Aβ is specifically toxic to neurons through a series of downstream events that lead to increased intracellular calcium levels, impaired mitochondrial redox activity, and increased free radical generation, culminating in neuronal dysfunction and death [3, 4]. AD is becoming a serious social problem in the aging global society, as the number of AD patients over 65 years of age has increased rapidly. Many clinical trials have sought unsuccessfully to modify disease progression in AD [5].
Mesenchymal stem cells (MSCs) are characterized by their homing effect, multi lineage differentiation potential, and immunomodulatory function [6, 7] under appropriate conditions. MSCs have neuroprotective effects through complex mechanisms, such as modulation of neuroinflammation, enhancement of cell survival signals, increased neurogenesis, and modulation of ubiquitinated proteins [8–12]. Human bone marrow (BM)-derived MSCs can reduce Aβ levels and tau phosphorylation with improved cognitive function in animal models of AD [13–15]. In addition, BM-MSCs significantly enhance autolysosome formation and clearance of Aβ in AD models, which may lead to increased neuronal survival [16]. Moreover, human umbilical cord blood-derived or placenta-derived MSCs can significantly improve learning and memory through the regulation of neuroinflammation or Aβ metabolism [9, 11]. MSCs also have great therapeutic potential for other neurological diseases and age-related diseases [17–19].
Based on these results, many clinical trials using MSCs are in progress in various neurodegenerative diseases [20]. However, the use of MSCs in cellular therapy poses several limitations. First, autologous MSCs are difficult to generate from aged donors, and do not function as well as MSCs produced in younger donors [21–24]. The use of allogenic MSCs derived from young donors provides a means to overcome the limitations of autologous MSCs, but introduces new problems, including donor-to-donor variability and the possibility of immune rejection [25, 26]. Repeated transplantations are also necessary because of the short duration of the activity and survival of transplanted MSCs [27, 28]. Therefore, mass production is essential for repetitive transplantation. One way to overcome the limitations inherent in the use of cultured MSCs for clinical trials is to use embryonic stem cell (ES)-derived MSCs. ES-MSCs readily proliferate during in vitro expansion [29] and the efficacy of ES-MSCs has been explored in difference neurological diseases [30, 31].
To date, no studies have addressed the various roles of ES-MSCs in eliciting therapeutic effects for AD. It is important to explain the potential effects and clinical applicability of ES-MSCs in AD. In the present study, we investigated the neuroprotective effect of ES-MSCs in cellular and animal models of AD, as well as the safety of intra-arterial administration of ES-MSCs in an AD animal model.
MATERIALS AND METHODS
Aβ preparation
Aβ peptide (AnaSpec, Inc., Freemont, CA, USA) was dissolved in 1% NH4OH and this solution was immediately diluted with phosphate buffered saline (PBS; Hyclone, Logan, UT, USA) and incubated at 37°C for 72 h to induce aggregation.
Cell culture
SH-SY5Y human neuroblastoma cell line was obtained from the Korean Cell Line Bank (Seoul, South Korea). Human BM-MSCs were obtained from CORESTEM Inc. (Seoul, South Korea). BM-MSCs and SH-SY5Y cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; HyClone) supplemented with 10% fetal bovine serum (HyClone) and an antibiotic mixture of penicillin and streptomycin (100 U/mL; Hyclone). Human ES-MSCs were obtained from Daewoong Pharmaceutical Co., Ltd. (Seoul, South Korea) were treated as previously described [32]. Human ES-MSCs were cultured with commercialized serum and xeno-free medium (Gibco, Carlsbad, CA, USA) containing L-glutamine (2 mM, Gibco) and a mixture of penicillin and streptomycin (100 U/mL; Hyclone). For in vitro experiments, SH-SY5Y cells were plated at a density of 2.0×105 cells/well and MSCs were co-cultured without direct contact using a Costar Transwell apparatus (Corning, Big Flats, NY, USA). MSCs were cultured on the permeable membrane of a Transwell insert. One day after plating, the SH-SY5Y cells were incubated in DMEM with 20μM Aβ for 24 h. After a change of the medium, the MSCs were cultured on a Transwell insert and SH-SY5Y cells were maintained on the bottom of a plate for 12 h and 24 h.
MSC characterization
Characterization of BM-MSCs was performed as described previously [28]. ES-MSC characterization was performed using modified minimal criteria of MSCs suggested by ISCT (2006). Cell surface markers were analyzed by flow cytometry using a FACS Verse™ device (BD Bioscience, San Jose, CA, USA) using human monoclonal antibodies. MSC surface markers (positive markers) were analyzed using anti-29-phycoerythrin (PE)-conjugated, anti-CD44-PE-conjugated, anti-CD73-PE-conjugated, and anti-CD105-PE-conjugated antibodies. Hematopoietic lineage marker (negative marker) was analyzed using the anti-CD45-fluorescein isothiocyanate (FITC)-conjugated antibodies. MHC class type II marker (negative marker) was analyzed using the anti-human leukocyte antigen-(HLA)-DR-FITC-conjugated antibody. Pluripotent stem cells markers (negative markers) were analyzed using the anti-stage-specific embryonic antigen 3 (SSEA3)-PE-conjugated, anti-Tra-1–60-FITC-conjugated, and anti-Tra-1–81-FITC-conjugated antibodies (all from BD Bioscience). The antibodies were compared with fluorescence and IgG type matched-isotype controls.
Differentiation potential of ES-MSCs was also analyzed using the tri-lineage differentiation of osteocytes, chondrocytes, and adipocytes. Induction of tri-lineage differentiation was performed using commercially available differentiation media (Gibco) according to the manufacturer’s protocols. Differentiated cells were confirmed by Alizarin red S (osteocytes), Alcian blue (chondrocytes), and Oil-red-O (adipocytes) staining.
Cell viability
To investigate the protective effect of MSCs on Aβ-induced cell death, the viability of co-cultured SH-SY5Y cells was assessed by Trypan blue exclusion test. The proportion of viable cells was calculated as the ratio of the number of dead (stained) cells to the number of viable (unstained) cells that were counted using a hemocytometer.
Aβ ELISA
Aβ ELISA was performed using a High Sensitivity Human Amyloid β ELISA kit (EZHS42; Millipore, Billerica, MA, USA). Diluted samples and standards included in the kit were applied (50μL each) to wells of microtiter plates precoated with antibody that specifically recognized the N-terminus of Aβ peptides. The color reaction was measured with a model ELX 800 automatic ELISA reader (Bio Tek Instruments, Winooski, VT, USA) at a wavelength of 450 nm and a reference wavelength of 590 nm. A standard curve was used to calculate the concentration of the samples were performed using the commercially available software (Microplate Manager; Bio Rad, Hercules, CA, USA).
Western blot
Cells and brain tissues were dissolved in ice-cold RIPA buffer (50 mM Tris-HCl, pH 7.5, with 150 mM sodium chloride, 1% Triton X-100, 1% sodium deoxycholate, 2 mM EDTA, and 0.1% sodium dodecyl sulfate) (Biosesang, Korea) plus protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA). The lysates were centrifuged at 4°C for 20 min (14,000 rpm) and supernatants were transferred to fresh tubes. An equal amount of total protein (10μg) was resolved by 10% or 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to a nitrocellulose membrane (Hybond ECL; Amersham Pharmacia Biotech, Piscataway, NJ, USA). Blots were blocked with 5% nonfat dried milk in PBS containing 0.05% Tween-20 (PBST) for 1 h at room temperature. After briefly washing with PBST, blots were incubated overnight at 4°C with specific antibodies: rabbit anti-LC3 (1 : 1000, Sigma-Aldrich, St. Louis, MO, USA), anti-LAMP2 (1 : 1000, Abcam, Cambridge, MA, USA), and anti-β-actin (1 : 1000, Santa Cruz Biotechnology, Santa Cruz, CA, USA). The blots were then incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit antibodies (1 : 5000, Santa Cruz Biotechnology). Immuno reactive proteins were detected by the enhanced chemiluminescence (ECL) western blotting detection system.
Animal models and ES-MSC administration
All procedures were performed in accordance with the Laboratory Animals Welfare Act, the Guide for the Care and Use of Laboratory Animals, and the Guidelines and Policies for Rodent Experimentation provided by the Institutional Animal Care and Use Committee at the Yonsei University Health System. Adult male or female Sprague-Dawley rats (200–220 g) were used to establish the Aβ-induced animal model for AD. Aβ (5μg/μL) was stereotaxically injected into the lateral ventricle (AP: –0.8 mm, ML: –1.4 mm, DV: –4.0 mm) for 5 min and the needle was left in the same place for further 5 min before being slowly withdrawn. The rats were randomly divided into three group: 1) the control group received normal saline, 2) the Aβ-treated group, 3) the MSC treatment group in Aβ-injected animals. Five days after Aβ injection, ES-MSCs were injected into the internal carotid artery (ICA). Based on previous studies, a concentration of 4×107 cells/60 kg body weight was selected as a standard dose of ES-MSCs [27, 28]. First, we examined neuroprotective effect of ES-MSCs in Aβ-injected animals using a standard dose of ES-MSCs. Then, we evaluated behavioral performance and ischemic safety using a ×0.5, ×1, ×2, ×5, and ×10 the standard dose of ES-MSCs. For ICA injection, animals were anesthetized with 30% isoflurane. As previously described [33], the common carotid artery (CCA) was exposed and the external carotid artery and pterygopalatine artery were ligated. ES-MSCs in 200μL of solvent (Daewoong, Inc., Seoul, Korea) were injected via a 33G microneedle into the CCA with preserved blood flow in the CCA and ICA for 30 s and bleeding control was performed by application of bio absorbable Gelfoam (Pfizer, New York, NY, USA) at the injection site for 5 min. Pressure was maintained at the injection site until bleeding ceased and then fixed external carotid artery and pterygopalatine artery were opened. The protocols for sample isolation and their subsequent use for research purposes were approved by the institutional review board of the Institutional Bioethics Committees, Korea (P01-201407-ES-01). The embryonic stem cells used are registered with the Korea Centers for Disease Control and Prevention (hES12011003). All rats were sacrificed 20 days after ICA injection.
Brain sample preparation
For immunofluorescence, rats were perfused with PBS and fixed with 4% paraformaldehyde (Sigma-Aldrich, St. Louis, MO, USA) dissolved in PBS at 20 days after ICA injection. Brains were removed and post-fixed overnight in buffered 4% paraformaldehyde at 4°C and stored in a sucrose solution (30% in PBS) for 3 days at 4°C until they sank. The brain tissue was embedded in the OCT compound and then frozen at –20°C. Brains were sliced coronally using a freezing microtome at 30μm for light microscopy, and free-floating sections were stored in a cryopreservative solution (3 : 3:4 glycerol: ethyleneglycol: PBS) at 4°C in preparation for immunohistochemistry straining. For western blotting, rats were euthanized at 20 days after ICA injection and brains were rapidly dissected and frozen at –70°C.
Immunohistochemistry
The coronal brain sections were rinsed twice with PBST for 5 min at room temperature and then blocked with 0.5% bovine serum albumin in 1×PBST. After blocking, sections were incubated overnight at 4°C with anti-NeuN as the primary antibody (1 : 500, Abcam). After overnight incubation, the brain sections were rinsed three times with 1×PBST (10 min per rinse) and incubated with the appropriate biotinylated secondary antibody and avidin–biotin complex (Elite Kit; Vector Laboratories, Burlingame, CA, USA) for 1 h at room temperature. Bound antibodies were visualized with 0.05% diaminobenzidine–HCl (DAB, Dako, Carpinteria, CA, USA). Stained sections were mounted on slides, dehydrated in ethanol, cleared in xylene and cover slipped. Parallel sections from the same animals were used across immunohistochemical experiments. The quantification of NeuN was the area in which NeuN was stained, expressed in % values.
Magnetic resonance imaging
Animals were anesthetized with 30% isoflurane and magnetic resonance imaging (MRI) scans were acquired using a BioSpec 94/20 USR (Bruker BioSpin, Ettlingen, Germany) 2 weeks after ES-MSC transplantation. The in vivo MRI data were obtained with the following parameters: 1) two-dimensional T2*-weighted gradient echo sequence with repetition time (TR) 1,000 ms and echo time (TE) 10.5 ms for transverse sections of the brain, and 2) T2-weighted two-dimensional spin echo sequence with TR 2,500 ms and TE 24 ms. MR-angiography (MRA) scans were obtained using a time of flight sequence technique (TR 18 ms, TE 4 ms, flip angle 80 degrees, slice thickness 0.35 mm, field of view 40×40 mm, and matrix size 256×256).
Ultrasonic measurement of blood flow
For the surgical procedures, animals were anesthetized with 10% isoflurane in a mixture of 70% N2O and 30% O2. Anesthesia was maintained with 2% isoflurane. During operative procedures, body temperature was monitored continuously with a rectal probe and maintained at 37°C using a homeothermic blanket control unit and a heating pad (Harvard Apparatus, Holliston, MA, USA). A midline cervical incision was made, and the right CCA was carefully dissected. An ultrasonic Doppler flow probe (MA0.7PSB; Transonic Instruments, Ithaca, NY, USA) was placed around the midportion of the CCA. Carotid blood flow was measured using a TS420 Blood Flow Meter (Transonic Instruments, Ithaca, NY, USA) and an IX-304T data acquisition system (iWorx Systems, Inc., Dover, NH). Computer-based analyses using Labscribe 2 software (version 2.045000; iWorx Systems, Inc.) were performed to minimize the bias when assessing results. The baseline CCA flow was measured for 3 min.
Morris water maze
Morris water maze was carried out in accordance with the procedure described previously [34–36]. The Morris water maze test was carried out in a circular tank (170 cm diameter) filled with water. The temperature of the water was maintained at 23°C to 25°C. A camera hanging above the tank was used to record swimming traces of rats. Prior to spatial learning, rats were subjected to visible platform Morris water maze. The aim of the visible platform test was to exclude mice with visual or motivational impairments, as well as to habituate them to the testing conditions. The pool was divided into four quadrants and a hidden platform was set 1 to 2 cm under the water in the targeted quadrant. Distal visual cues are arrayed on each quadrant, and in general, rats are able to learn the location of the hidden platform based on these cues. Each rat was placed in the water facing the wall of the tank from a different quadrant in all trials. The learning ability of rats was examined using the hidden platform test from day 1 to 5, which allowed rats to swim three times per training day. Memory test was evaluated by one probe trial on day 6 after removing the platform from the pool. The escape latency in the hidden platform test and swimming traces in all experiments were recorded for off-line analysis with Smart 3.0 software (Panlab, S.L.U, Barcelona, Spain).
Statistical analyses
The data of the present study are expressed as mean±SE. Viability, protein expression and behavioral parameter were compared using one-way analysis of variance (ANOVA).Bonferroni correction was used for multiple comparisons after performing thee one-way ANOVA. The statistical analyses were performed with SPSS (version 25.0; IBM Corporation, Armonk, NY, USA), and results with a two-tailed p-value <0.05 were considered statistically significant.
RESULTS
Characterization of ES-MSCs
We first confirmed that the ES-MSCs had gained characteristic MSC surface markers. Flow cytometry analysis revealed that ES-MSCs were positive for expression of the specific mesenchymal markers CD29, CD44, CD73, and CD105. However, cells were negative for the hematopoietic lineage marker (CD45), MHC class type II (HLA-DR) and ES cell markers (SSEM-3, Tra-1–60, and Tra-1–81) (Fig. 1A, B). To investigate the multi-differentiation potential, ES-MSCs were cultured in osteogenic, adipogenic, and chondrogenic induction medium. ES-MSCs successfully differentiated into osteoblasts, chondrocytes, and adipocytes (Fig. 1C).
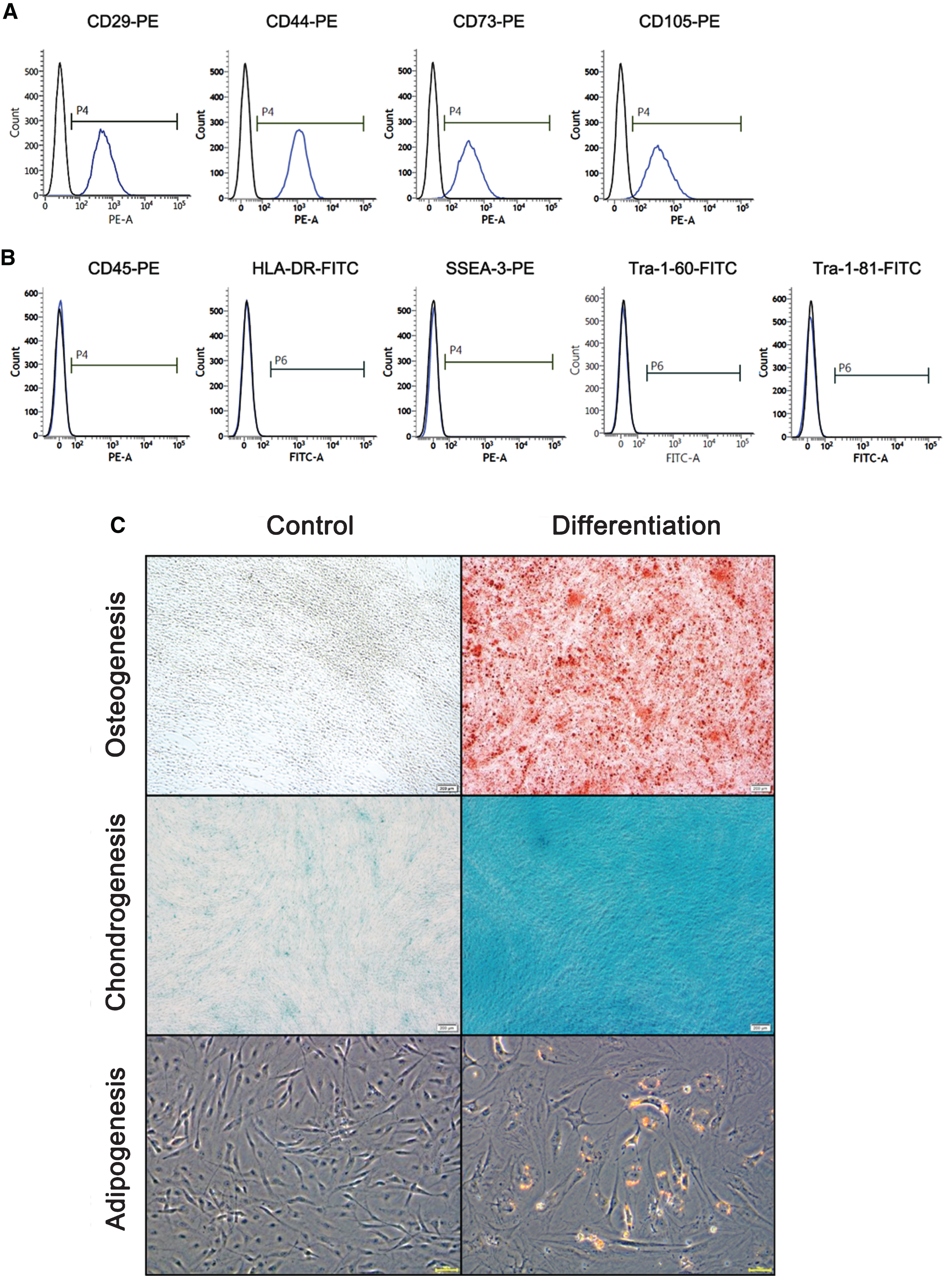
Characterization of ES-MSCs. A) Positive surface markers of ES-MSCs were analyzed using CD29, CD44, CD73, and CD105. B) Negative surface markers of ES-MSCs were analyzed using CD45 (hematopoietic lineage), HLA-DR (MHC class type II), SSEA-3, Tra-1–60, and Tra-1–81 (embryonic stem cell marker). C) Differentiation potentials of ES-MSCs were analyzed using osteocyte-, chondrocyte-, and adipocyte-specific differentiation media. After the induction of differentiation, osteogenesis, chondrogenesis, and apidogenesis were assessed using the Alizarin red staining (upper panel), Alcian blue staining (middle panel), and Oil-red-O staining (lower panel).
Effects of MSCs on Aβ-induced cell death according to different cell types
We identified the effect of MSCs on Aβ-induced cell death depending on different cell types (Fig. 2A). After treatment of Aβ for 24 h in SH-SY5Y cells, BM-MSCs, and EM-MSCs were co-cultured for 12 h and 24 h. There was no difference in Aβ-induced cell death between the Aβ treatment group and the MSC co-culture group at 12 h of co-culture. At 24 h of MSC co-culture, the BM-MSC (p = 0.037) and ES-MSC groups (p < 0.001) displayed significantly higher cell viability than the Aβ treatment group. Among the different types of MSCs, ES-MSCs exhibited higher cell viability relative to BM-MSCs (p = 0.047, Fig. 2A). In a previous study, we reported that BM-MSCs enhanced autophagy and the lysosomal clearance of Aβ [16]. Therefore, we evaluated whether MSCs increased the induction of autophagy in SH-SY5Y cells treated with the Aβ in the different cell types. Compared to the Aβ treatment group, MSC treatment significantly increased the expressions of LC3-II and LAMP2 at 12 h (p < 0.05) or 24 h (each p < 0.01) after MSC co-culture, and the levels of LC3-II and LAMP2 were comparable between BM-MSCs and ES-MSCs (Fig. 2B). Next, we examined whether MSC treatment changed the levels of Aβ in SH-SY5Y cells. Aβ treatment led to increased intracellular Aβ levels, whereas MSC treatment significantly decreased intracellular Aβ levels 12 h (each p < 0.05) or 24 h (each p < 0.001) after MSC co-culture. In addition, intracellular Aβ levels tended to be lower in ES-MSCs relative to BM-MSCs 24 h after MSC co-culture (p = 0.008; Fig. 2C).
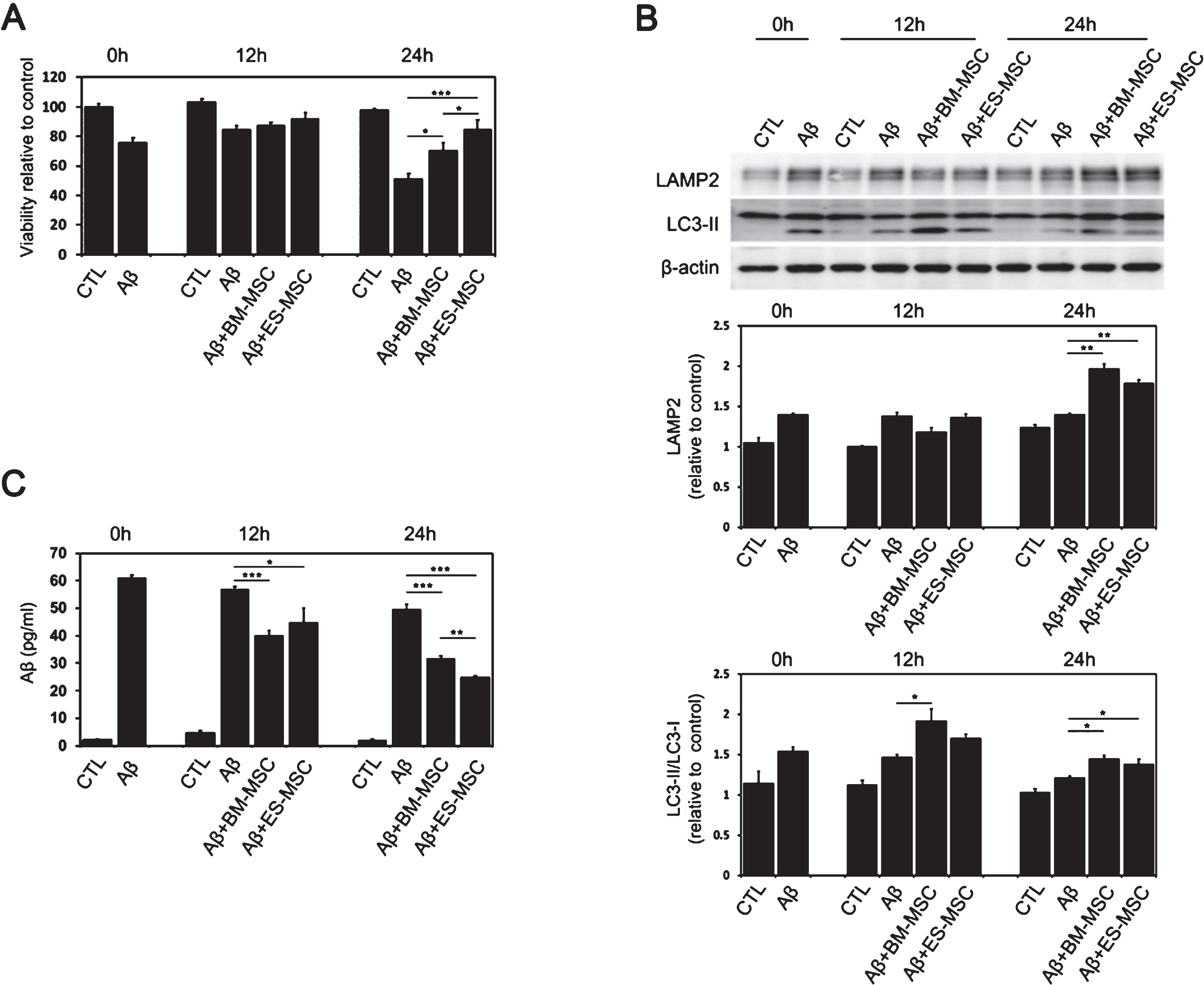
Effects of MSCs on Aβ-induced cell death according to different cell types. Aβ-treated SH-SY5Y cells were co-cultured with BM-MSCs and EM-MSCs for 12 h and 24 h. A) Cell viability using a Trypan blue exclusion assay revealed greater viability of ES-MSC and BM-MSC populations than that of the Aβ treatment group. Cell viability was greater in the ES-MSC group relative to the BM-MSC group (n = 5, per group). B) The expression of LAMP2 and LC3-II was higher in the MSCs-treated group relative to the control group, and the expression levels were comparable between BM-MSCs and ES-MSCs (n = 5, per group). C) MSC treatment significantly decreased intracellular Aβ levels at 12 h or 24 h after MSC co-culture, and the intracellular Aβ levels tended to be lower in the ES-MSC group relative to the BM-MSC group at 24 h after MSC co-culture (n = 5, per group). The data are presented as the mean±SE.*p < 0.05, **p < 0.01, ***p < 0.001.
Behavioral analysis following intra-arterial injection of standard dose of ES-MSCs
The in vitro data indicated a pro-survival effect of ES-MSCs against Aβ that was comparable to BM-MSCs. Thus, we performed an in vivo analysis to identify whether ES-MSCs could modulate memory performance of Aβ-treated animals. In the first week, all rats reduced their escape time during the 5-day training phase (Fig. 3A). In the second week, only Aβ-injected rats had a slower escape time during the 5-day training relative to controls or ES-MSC-treated rats (each p < 0.05, Fig. 3B). During the probe test, Aβ-treated animals who received ES-MSCs spent a significantly longer time in the target quadrant compared to the Aβ-injected group at either 1 week (p < 0.001) or 2 weeks (p < 0.001, Fig. 3B and 3C). Similarly, the number of target crossings for the platform position was significantly increased in the ES-MSC treatment group compared to the Aβ-injected group (each p < 0.05, Fig. 3D). The beneficial effect of ES-MSCs on cognitive performance in female rats was identical (Supplementary Figure 1).

Behavioral analysis following intra-arterial injection of a standard dose of ES-MSCs. After the injection of ES-MSCs, the water maze test was performed on week 1 and week 2. A) In the second week, only Aβ-injected rats had a slower escape time during the 5-day training relative to controls or ES-MSCs-treated rats (n = 10, per group). B, C) ES-MSC treatment in Aβ-treated animals resulted in a significantly higher percentage of time in the target quadrant compared to the Aβ-injected group (n = 10, per group) at either 1 week (B) or 2 weeks (C). D) The number of target crossings for the platform position was significantly increased in the ES-MSC treatment group compared to the Aβ-injected group (n = 10, per group). The data are presented as the mean±SE. *p< 0.05, **p < 0.01, ***p < 0.001.
Standard does of ES-MSCs inhibits Aβ-induced cell death and promotes autophagy for Aβ clearance
Immunohistochemical analysis showed that Aβ treatment led to a significant reduction of NeuN-positive neurons in the CA1, CA3, and dentate gyrus compared to control animals. However, ES-MSC treatment in Aβ-treated animals significantly increased NeuN-positive neurons in the hippocampus relative to the Aβ-treated group (CA1, p = 0.008; CA2, p < 0.001; DG, p = 0.006; Fig. 4A). Next, we investigated the expression of LAMP2 by western blotting. MSC treatment in Aβ-injected rats led to a significant increase in the expression of LAMP2 compared to the Aβ-injected group (p = 0.016, Fig. 4B). Concomitantly, MSC treatment in Aβ-injected rats significantly decreased the levels of Aβ in the hippocampus compared to the Aβ-injected group (p < 0.001, Fig. 4C). The pro-survival effect of ES-MSCs on hippocampal neurons via autophagy modulation was identical in female rats (Supplementary Figure 2).
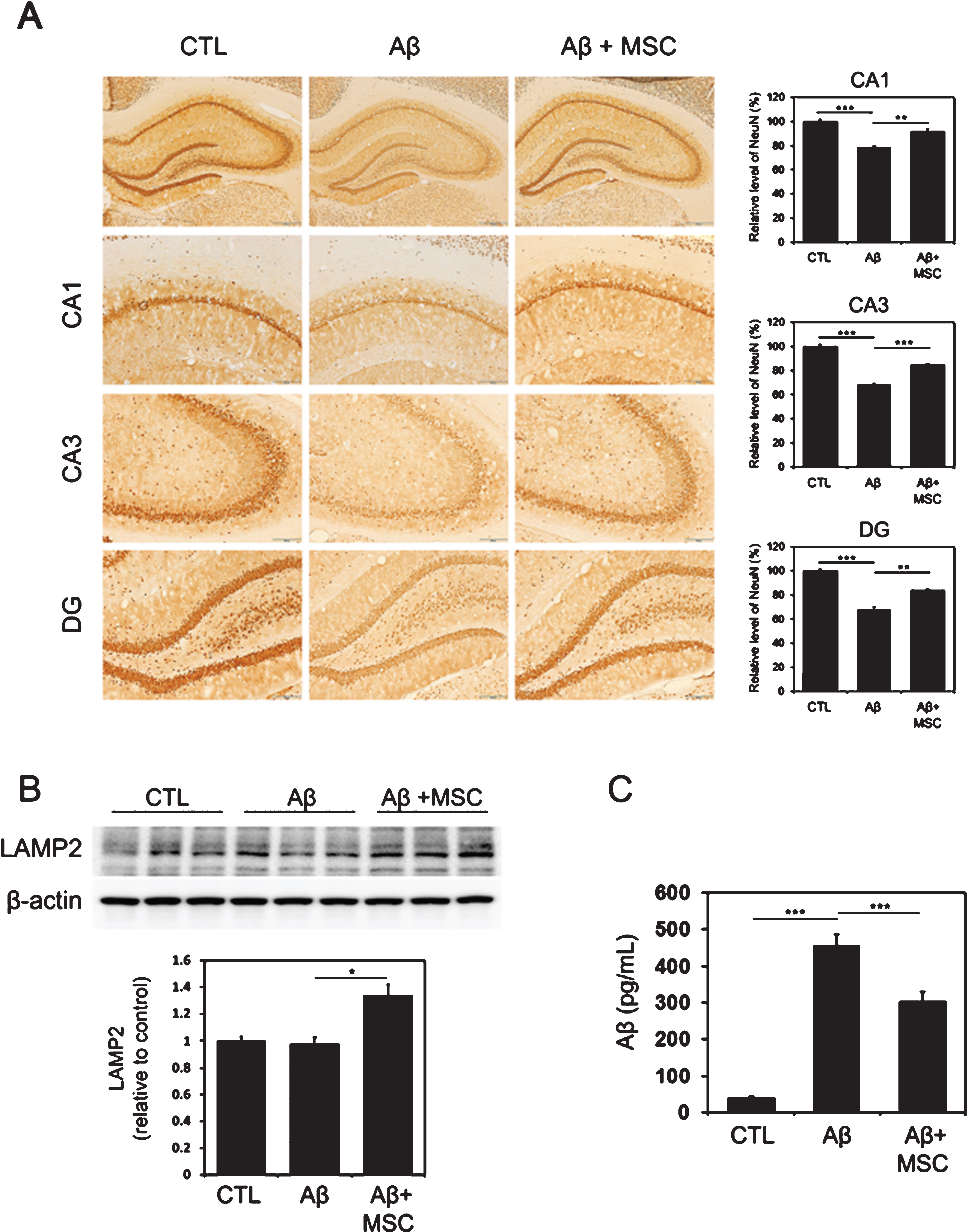
Standard dose of ES-MSCs inhibits Aβ-induced cell death and promotes autophagy for Aβ clearance. A) Immunohistochemical analysis revealed that ES-MSC treatment in Aβ-treated animals significantly increased NeuN-positive neurons in the hippocampus relative to the Aβ-treated group (n = 5, per group). B) ES-MSC treatment in Aβ-injected rats significantly increased the expression of LAMP2 relative to Aβ-injected animals (n = 5, per group). C) Compared to the Aβ-injected group, ES-MSC treatment in Aβ-injected rats significantly decreased the levels of Aβ in the hippocampus (n = 5, per group). The data are presented as the mean±SE. Scale bar: 200μm. *p < 0.05, **p < 0.01, ***p < 0.001.
Behavioral analysis following intra-arterial injection of different doses of ES-MSCs
In the first week, all rats reduced their escape time during the 5-day training phase (Fig. 5A). In the second week, rats injected with different doses of MSCs displayed shortened escape latency during the 5-day training phase compared to the Aβ-injected group (each p < 0.05, Fig. 5B). Compared to the Aβ-injected group, ES-MSC treatment with ×1, ×2, ×5, or ×10 the standard dose resulted in a significantly higher percentage of time in the target quadrant during probe test in both week 1 (each p < 0.05, Fig. 5C, E) and week 2 (each p < 0.001, Fig. 5D, F). However, ES-MSC treatment with ×0.5 the standard produced no significant difference in time in the target quadrant relative to the only Aβ-injected group (Fig. 5E, F). The beneficial effect of ES-MSCs on memory performance measured by the water maze test was superior using doses ×2 and ×5 of the standard dose compared to the ×1 standard dose at either 1 week or 2 weeks after ES-MSC treatment. However, AD animals receiving ×10 of the standard dose did not differ from those receiving a standard dose. Additionally, memory performance did not differ between AD animals receiving ×2 and ×5 of the standard dose (Fig. 5E, F). The number of target crossings did not differ depending on different doses of ES-MSCs at either 1 week (Fig. 5G) or 2 weeks (Fig. 5H) after ES-MSC treatment.

Behavioral analysis following intra-arterial injection of different doses of ES-MSCs. After the injection of ES-MSCs with a different dose, the water maze test was performed on week 1 and week 2 (A and B). In the second week, rats injected with MSCs of different dose displayed shortened escape latency during the 5 days training phase compared to the Aβ-injected group with no difference in the first week (n = 8, per group). C–F) Compared to the Aβ-injected group, ES-MSC treatment with ×1, ×2, ×5, or ×10 the standard dose resulted in a significantly higher percentage of time in the target quadrant during probe test (n = 8, per group) in both week 1 (C and E) and week 2 (D and F). Beneficial effect of ES-MSCs on memory performance was superior in a dose×2 and×5 the standard dose compared to×1 the standard dose. However, AD animals that received ×10 the standard dose did not differ from those receiving a standard dose. G, H) The number of target crossing did not differ depending on different doses of ES-MSCs at either 1 week (G) or 2 weeks (H) after ES-MSC treatment (n = 8, per group). The data are presented as the mean±SE. *p < 0.05, ***p < 0.001.
Evaluation of internal carotid artery flow and cerebral ischemia following intra-arterial injection of different doses of ES-MSCs
To evaluate the efficacy of intra-arterially injected ES-MSCs, we chose ×2 and ×10 of the standard dose of ES-MSCs because these two doses corresponded to the lowest dose exhibiting a comparable effect in terms of better cognitive performance and to the highest dose in terms of concentrations, respectively. The ICA blood flow evaluated by an ultrasonic flow meter was well preserved in AD animals receiving ES-MSCs with ×2 or ×10 the standard dose (Fig. 6A). In addition, no occlusive lesion of the ICA on MR-angiography was observed in AD animals receiving ES-MSCs with ×2 or ×10 the standard dose (Fig. 6B). T2-weighted MRI did not reveal hyperintense signals in the brain parenchymal of AD animals receiving ES-MSCs with ×2 or ×10 the standard dose (Fig. 6C).
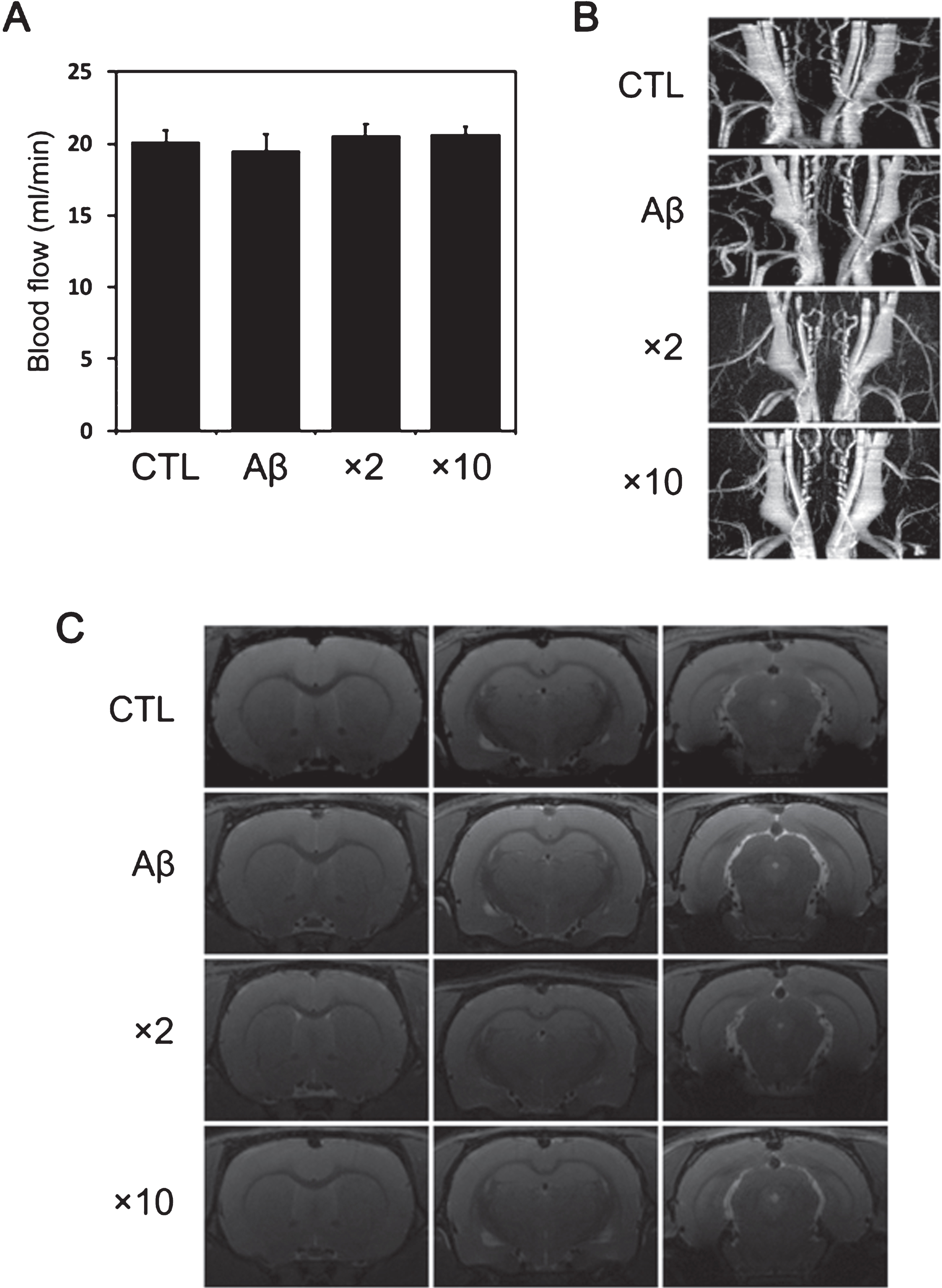
Evaluation of internal carotid artery flow and cerebral ischemia following intra-arterial injection of different doses of ES-MSCs. A) The ICA blood flow on an ultrasonic flow meter was well preserved in AD animals receiving ES-MSCs with ×2 or ×10 the standard dose (n = 5, per group). B) MR-angiography revealed no occlusive lesions of the in AD animals receiving ES-MSCs with ×2 or ×10 the standard dose (n = 5, per group). C) T2-weighted MRI revealed no hyperintense signals in the brain parenchymal of AD animals receiving ES-MSCs (n = 5, per group).
DISCUSSION
The present study revealed that ES-MSCs had a comparable pro-survival effect against Aβ-induced cell death relative to BM-MSCs. In addition, if their concentration was appropriate, intra-arterial injection of ES-MSCs was safe and had a neuroprotective effect in hippocampal neurons and improved cognitive performance in AD models. These data indicate that intra-arterial administration of ES-MSCs is feasible and efficacious with respect to neuronal viability and behavioral performance in Aβ-induced AD models.
In terms of pathogenesis of AD, ample evidence from animal and clinical studies have shown that accumulation of Aβ, hyperphosphorylation and accumulation of tau, neuroinflammation, and dysfunction in cellular clearing systems are the main contributors to AD pathologies [37–39]. Several drugs that focus on pathogenic mechanisms have been developed, and many clinical trials have been performed to uncover the efficacy of various treatments in AD patients. However, these trials have failed to provide ample evidence of neuroprotection or had limited efficacy in AD patients [5]. These results indicate that a certain molecular candidate that affect only one pathogenic target is not effective; rather, it is necessary to have multi targets that control multiple pathogenic steps.
Given that MSCs from various sources have pleiotropic effects against the AD-related microenvironment [8–12], the present study used Aβ-induced AD cellular and animal models to examine the feasibility and efficacy of the intra-arterial administration of various doses of ES-MSCs to extend its clinical application to patients with AD. We found that ES-MSCs increased the viability of SH-SY5Y cells treated with Aβ by modulating autophagolysosomes with an efficacy comparable to BM-MSCs. Autophagy is an essential process in the removal of damaged organs and macromolecules [40, 41]. This degradation pathway depends on the formation of autophagosomes with double membranes, which combine with lysosomes, degrade cell contents, and are associated with various human diseases, including neurodegenerative diseases. Several studies have shown that small molecular compounds that can activate autophagy or lysosomal proteolysis can significantly reduce the Aβ load in AD [42, 43]. Specifically, we found that MSCs from various sources led to an increase in autophagy markers, such as LC3-II and LAMP2, followed by significantly decreased intracellular Aβ levels. Interestingly, the efficacy of autophagy induction in ES-MSCs was comparable to that in BM-MSCs, but intracellular Aβ levels were more significantly reduced in ES-MSCs than in BM-MSCs. These data suggest that, like other MSCs from different sources, ES-MSCs could enhance clearance of Aβ by modulating the autophagy pathway, which benefited survival. In addition, in vivo ES-MSCs in the AD animal models could inhibit Aβ-induced cell death in the hippocampus and promote autophagolysosome-mediated Aβ clearance and concomitant decrease in the levels of Aβ in the hippocampus. Furthermore, ES-MSC treatment in Aβ-treated animals produced better memory performance compared to the Aβ-injected group. The pro-survival effect of ES-MSCs on hippocampal neurons via autophagy modulation was identically observed in male and female rats. These data provide in vivo evidence that ES-MSCs have a neuroprotective property against the AD-related microenvironment.
In the present study, a concentration of ES-MSCs that was used in previous clinical trial and a rodent study (4×107 cells/60 kg body weight) was selected as the standard concentration [27, 28] to evaluate the safety of intra-arterial administration. In a comparative analysis between the groups according to the dose of ES-MSCs, behavioral analyses showed that memory performance was better in AD animals receiving ES-MSCs with ×2 or ×5 the standard dose compared those receiving ×1 or ×10 of the standard dose with no between-group difference for the ×2 and ×5 standard dose groups. However, memory performance in AD animals receiving ×0.5 the standard dose did not differ from the Aβ-injected group. These data suggest that ×2 of the standard dose of ES-MSCs may be the most effective dose for intra-arterial injections in AD animals with respect to cognitive performance. Regarding ischemic complications associated with intra-arterial injection of MSCs, the amount of cells in arterial blood and the rate of arterial flow are important factors in determining the development of ischemic events, even if the rate of complications vary according to experimental methods of the intra-arterial injection [33, 45]. In a previous study, we demonstrated that intra-arterial administration of higher doses of MSCs led to in situ occlusion of the injected artery or ischemic insults in the distal cerebral artery. Specifically, when multiple system atrophy animals were injected with ×50 the standard dose, the cells were regurgitated from the injection site in the ICA probably due to arterial occlusion, which would increase mortality. In addition, intra-arterial injection with ×20 the standard dose was associated with the development of ischemic lesions in the distal MCA territory, which negated the behavioral efficacy of MSCs [28]. Based on these results, we selected×10 the standard dose of ES-MSCs as the highest dose. As well, ×2 the standard dose was also chosen as it was the lowest dose that produced a comparable effect in terms of better cognitive performance. These two doses did not lead to the development of in situ occlusion of the injected artery or ischemic insults in distal arteries. However, it is difficult to explain why ×10 the standard dose of ES-MSCs did not produce better memory performance, since cerebral perfusion to the distal small artery or arteriole that is low and insufficient to cause ischemic insults [46] may negatively impact behavioral performance. Accordingly, the present findings suggest that intra-arterial administration of an appropriate cell density of ES-MSCs is safe and poses no risk of cerebral ischemia. Further research is warranted to address the other safety issues associated with intra-arterial injection of ES-MSCs.
ES-MSC has several advantages over other MSCs [29–31]. 1) ES-MSCs readily proliferate during in vitro expansion, which facilitates mass production. 2) ES-MSC has excellent secretion of various cytokines and small molecules. 2) ES-MSCs have comparable neuroprotective ability in neurological diseases relative to MSCs from various sources. Therefore, ES-MSCs are expected to potentially be applicable to a number of neurological disorders, such as Parkinson’s disease, Huntington’s disease, multiple system atrophy, or amyotrophic lateral sclerosis. In addition, these advantages of ES-MSC are expected to be a promising strategy for treating various major systemic diseases via tissue repair and soluble factor communications [18]. In terms of the x-factors controlling AD microenvironments, a dozen of factors secreted from the MSCs have been suggested to modulate beta-amyloid or tau protein related neurotoxicity as well as its related neuroinflammation. For example, UCB-MSCs have a neuroprotective effect in vitro against Aβ toxicity via gatectin-3 secretion [47]. In addition, soluble intercellular adhesion molecule-1 secreted from UCB-MSCs enhances Aβ clearance [48]. Moreover, CCL5, a chemoattractive factor secreted from BM-MSC, plays an important role in the recruitment of microglia, which leads to the reduction of Aβ deposition and memory impairment in AD mice [49]. Neprilysin secreted from adipose tissue-derived MSC is known to decrease intracellular and secreted Aβ levels [50]. In additionally, we have found other candidates secreted from MSCs to control another toxic protein related microenvironments, such as alpha-synuclein; galectin-1 for cell-to-cell transmission of alpha synuclein [51], eEF1A-2 for axonal transport of alpha synuclein [52], and IL-4 for microglia polarization [53]. However, we think that so called the discovery of x-factors would be beyond the scope of the present study.
In summary, the present study has provided important details that intra-arterial administration of ES-MSCs is feasible and efficient route, resulting in improvement of neuronal survival and behavioral performance in Aβ-induced AD models. The data support the therapeutic potential of ES-MSCs and clinical applications of the intra-arterial route of ES-MSC administration in AD patients.