
Abstract
Alzheimer’s disease (AD) is a highly age-related cognitive decline frequently attacking the elderly. Senescence-accelerated mouse-prone 8 (SAMP8) is an ideal model to study AD, displaying age-related learning and memory disorders. SAMP8 mice exhibit most features of pathogenesis of AD, including an abnormal expression of anti-aging factors, oxidative stress, inflammation, amyloid-β (Aβ) deposits, tau hyperphosphorylation, endoplasmic reticulum stress, abnormal autophagy activity, and disruption of intestinal flora. SAMP8 mice, therefore, have visualized the understanding of AD, and also provided effective ways to find new therapeutic targets. This review focused on the age-related pathogenesis in SAMP8 mice, to advance the understanding of age-related learning and memory decline and clarify the mechanisms. Furthermore, this review will provide extensive foundations for SAMP8 mice used in therapeutics for AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disorder mainly affecting people aged 65 and over [1]. At present, China has more than 200 million people over the age of 60. The speed of aging process and elderly population in China are far beyond the developed countries. By 2040, more than 20% of the population will be aged 65 and over, and the number of people aged 80 and over is rising by 5% a year to more than 74 million [2]. The prevalence of dementia in China, Hong Kong, and Taiwan is estimated 9.5 million [3]. The onset of AD is hidden and gradually worsens, involving multiple systems of the whole body, and the AD patients gradually lose the ability to take care of themselves. Few effective treatments are available for aging-related neurodegenerative diseases, causing a serious decline in the patient’s quality of life, and bringing about heavy economic burden to society and family [4].
To study the pathogenesis of AD and therapeutic interventions, a variety of animal models of AD have been created, from fruit fly, mouse, rat, rabbit, dog, to non-human primate, but the mouse models are the most popular [5]. AD transgenic mouse models include: APP transgenic, APP/PS1 double transgenic, and Tau/APP/PS1 triple transgenic, APP/NOS2–/–, PDAPP, TASD-41, Tau Tg, TauP301L, JNPL3, Tg 2576, Presenilin mutations (PSEN1), APOE ɛ4, and ob/ob (leptin), etc. [5–7]. Chemical and lesion-induced rodent models of AD include: Aβ-induced type (e.g., Aβ25–35), cholinergic nerve injury type (e.g., scopolamine), inflammation type (e.g., LPS), glucose/energy metabolism impairment (e.g., D-galactose), and heavy metal poisoning (e.g., aluminum) as well as lesion of the cholinergic neurons of the basal forebrain [8, 9]. Among mouse models, the spontaneous senescence-accelerated mouse-prone (SAMP) model is widely used and recognized. SAMP and anti-senescence mouse-prone (SAMR) mice are selected from AKR/J mice by professor Toshio Takeda, which contain a total of 12 lines of inbred strains, including SAMP1, SAMP2, SAMP3, SAMP6, SAMP7, SAMP8, SAMP9, SAMP10, SAMP11, and SAMR1, SAMR4, and SAMR5 [10]. Each SAMP strain exhibits specific disease phenotypes associated with age, which is similar to the symptoms of aging in humans such as senile amyloidosis (SAMP1), senile osteoporosis (SAMP6), and age-dependent deficits in learning and memory (SAMP8) [11]. The SAMP8 mouse model is characterized by a significant decline in learning and memory, but is also associated with skin aging [12], heart aging [13], anxiety [14], and hearing loss [15], and is the most widely used in dementia and senility research. Therefore, SAMP8 mice play an important role in anti-aging research, especially in AD research. This paper reviewed the application of SAMP8 mice in AD research, focusing on advances of molecular mechanisms leading to aging and AD, as well as potential therapeutic targets for drug development.
AGE-DEPENDENT RECOGNITION DECLINE AND AD-LIKE SYMPTOMS IN SAMP8 MICE
AD patients showed age-dependent progression in cognitive decline, behavioral impairments, and brain pathologies [16]. Animal models are essential for identifying AD disease mechanisms and developing effective therapeutic strategies [5]. Chemical- and lesion-induced models only focus on one of the specific pathways and the extent of the lesion is variable [8]. The most common transgenic mouse models, however, mimic familial AD, accounting for only a small percentage of cases, and are restricted to the expression of AD-related pathology associated with specific gene mutations, more relevant to study the mechanism of mutation, rather than AD itself [6, 17].
SAMP8 mice is a spontaneous animal model of accelerated aging. Brains of SAMP8 mice have shown age-associated pathologies in the hippocampus and other brain regions [10], including early age-related Aβ deposition, age-related aberrant hyperphosphorylation of tau-like neurofibrillary tangles (NFTs), and others. All these changes mimic AD patients’ pathologies, making SAMP8 mice a valuable model to study AD and other cognitive disorders [10, 18].
Taking advantage of age-related pathologies, candidate drugs can be administered along the aging process: some target the early onset of AD (6–8 weeks of age), while the majority of studies initiate at the adulthood (4–6 months of age), and some target the late stages of AD (10–12 months). The therapeutic candidates were administered not for days, but for months to better evaluate the therapeutic effects on age-related changes. For example, the long-term (10 months) administration of melatonin to 6-week-old SAMP8 and SAMR1 mice was performed to examine oxidative stress, inflammation, and vasoprotection [19]; green tea extracts were administered in diet starting at 2 months of age for 4 consecutive months, and AD pathologies were examined at 6 months of age [20]. Icariin was administered to 5-month-old SAMP8 mice for 3 consecutive months to examine AD pathologies and autophagy [21]. Liuwei Dihuang decoction (LW-AFC) was administered at 6 months of age for 5.5 months, and the effects on brain and microbiota was examined [22, 23]. In some cases, drugs were administered when AD pathology is well-formed, for example, metformin was administered to 11-month-old SAMP8 mice for 8 weeks to examine the endpoint improvements [24]. 1-Deoxynojirimycin from mulberry leaves was administered to 10-month-old SAMP8 mice for two months to examine cognitive decline, microglia activation, inflammation, Aβ deposition, and neurotrophic factors BDNF for repair [25]. Therefore, it is important to know age-associated abnormalities and pathologies for better experimental design.
The lifespan of SAMP8 mice is about 10–12 months, while SAMR1 mice is about 19–21 months [26–29]. The median survival time of SAMP8 mice is 9.7–10.4 months, while that of control SAMR1 mice is 16.3–17.8 months [10]. However, the mice in the US and Europe have a similar phenotype but at an older age, possibly due to animal maintenance conditions. The age-related cognitive decline and brain pathological features in SAMP8 mice in comparison with age-matched SAMR1 mice or younger SAMP8 mice are listed in Table 1. The major age-related pathologies include aberrant gene expression and spongy degeneration [10]; oxidative stress [18]; tau hyperphosphorylation [17, 30]; cognitive impairment [17, 31]; glial degeneration and inflammation [10, 32]; Aβ deposition [7, 33]; and Aβ plaques and neuron loss [10, 34].
The relationship between first occurred AD-like changes and monthly age in the brain of SAMP8 mice (M: month)
THE PATHOGENESIS OF LEARNING AND MEMORY DISORDER IN SAMP8 MICE
Abnormal expression of anti-aging factors and oxidative stress injury
Genetic defects and damage are the main causes for the rapid aging of SAMP8 mice. Previous studies have shown that compared with age-matched SAMR1 mice, the expression and activity of anti-aging factors such as silent information regulator type (sirtuin/Sirt), Klothos, and Forkhead box class O (FoxOs) are significantly decreased in SAMP8 mice [35–37].
Sirts are NAD+ dependent histone deacetylases involved in the regulation of aging and longevity. Sirts are closely related to the occurrence and development of AD [38]. Clinical studies have shown that enhancing the activity of Sirt 1 and Sirt 3 in the brain of AD patients can significantly improve their neurological impairment and cognitive impairment [38–40]. Similarly, in the SAMP8 mice, the content of Sirt 1 in the pancreatic tissue was also significantly decreased compared with age-matched SAMR1 mice, while increased expression of Sirt 1 significantly improved the age-related changes in the pancreas of SAMP8 mice [41, 42]. Previous studies including our study (unpublished) have found that the protein expression of Sirt 1 in the hippocampus of SAMP8 mice was significantly decreased [43], while the cognitive impairment of SAMP8 could be improved by drug intervention to increase the protein expression of Sirt 1 in the hippocampus [44]. The expression of Sirt 1 and Sirt 3 is also closely related to oxidative stress injury [45]. The expression of other subtypes of the Sirt family in SAMP8 mice in relation to AD-like changes deserve further study.
Klotho, a type I transmembrane protein, encoded by the senescence suppressor gene klotho. According to the function of the protein, Klotho can be divided into membrane binding type and secretory type, and the expression level of secretory type is significantly higher than that of membrane binding type. Klotho protein is mainly expressed in renal tubular epithelium and cerebral choroid plexus, and the extracellular domain of Klotho can be released into the blood and cerebrospinal fluid by proteases from the plasma membrane. Klotho protein is proposed to play a neuroprotective role in AD patients, improve mild cognitive impairment and late-life depression [47]. When SAMP8 mice get older, the expression of Klotho in brain is gradually decreased, and increased Klotho expression could improve the age-related cognitive impairment in SAMP8 mice [35, 48]. Klotho protein has a strong anti-aging activity, interacts with Sirt 1 to play roles in aging vasculature and age-related diseases [49, 50]. Moreover, increased Klotho expression can also promote the clearance of Aβ and induce autophagy activity [46].
Activation of FoxOs can increase the expression of Mn-superoxide dismutase (MnSOD), promote the clearance of ROS, and enhance the ability to resist oxidative stress [51]. FoxOs is also involved in a variety of biological processes, such as repairing DNA damage and extending lifespan. Previous studies have shown that the protein levels of FoxO 1 in the liver of SAMP8 mice decreased with age [52]. Another study found that the mRNA expression of FoxO 1 and FoxO 3A decreased with age, leading to diminished anti-oxidative stress ability and rapid aging in SAMP8 mice [36, 53]. Therefore, with the increase of age, the expression of some subtypes of anti-aging factors such as Sirt, Klothos, and FoxOs decreased, leading to the decline of anti-oxidative stress ability of SAMP8 mice.
Oxidative stress is a risk factor for many diseases, including aging, cancer, atherosclerosis, and neurodegenerative diseases such as Parkinson’s disease and AD [54]. Due to congenital genetic defects and damage, SAMP8 mice showed impaired antioxidant defense at 8 weeks of age compared to age-matched SAMR1, the contents of glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), catalase (CAT), and glutathione reductase (GSSG-Rd, GR) decreased, and malondialdehyde (MDA) levels increased in the brain of SAMP8 mice with aging, which eventually lead to increase oxidative damage in the brain of SAMP8 mice [55]. It has been reported that the use of antioxidants can protect brain proteins against reactive oxygen species and prolong the life of SAMP8 [56]. Dietary supplements of the antioxidant preparation (spray-dried porcine plasma) for 4–6 months improved learning and memory impairment in SAMP8 mice [57]. Testosterone given to 10-month-old SAMP8 mice for 60 days increased hippocampal CA1 neurons and improved cognitive deficits, and its mechanism is related to its antioxidant ability to improve synaptic plasticity [58]. Meanwhile, intensified oxidative stress will lead to irreversible DNA damage and further worsen the SAMP8 learning and memory impairments [59].
Aβ deposition and tau hyperphosphorylation
The two major pathological features of AD are the abnormal deposition of Aβ and tau hyperphosphorylation.
Amyloid-β protein precursor (AβPP) is a macromolecular transmembrane glycoprotein with important functions, which can generate many protein fragments with different functions for the nervous system after being cleaved by enzymes [60]. Under normal conditions, AβPP goes through an alternative non-amyloid protein metabolism pathway, mediated by α-secretases (three types of enzymes involved in the activity of α-secretases: ADAM9, ADAM10, and ADAM17) [61] and β-secretase (BACE1), eventually generates soluble amyloid-β protein precursor α (sAβPPα). The other pathway is the Aβ pathway. By the cleavage of BACE1 and γ-secretases, an enzyme complex consists of presenilin (PS), nicastrin (NCT), anterior pharynx defective (APH-1), and presenilin enhancer-2 (PEN-2). Soluble sAβPPβ, Aβ1–40, and Aβ1–42 are produced, and Aβ1–42 fragment aggregates in the extracellular to form senile plaques in the brain. The increased level of Aβ1–42 is closely related to AD [62]. Excessive production of AβPP and Aβ levels were detected in the cerebral cortex and hippocampus of SAMP8 mice [63]. SAMP8 mice showed Aβ deposition in the hippocampus as early as 6 months of age, which is increased with age, eventually leading to neuronal damage and cognitive dysfunction [7, 10]. Drug intervention could reduce the production and increase the degradation of Aβ, effectively reduce the Aβ deposition in the brain, then improve learning and memory impairment in SAMP8 mice and delay brain aging [64–66]. The proteins related to the production and degradation of Aβ in SAMP8 mice are shown in Table 2.
The changes of proteins associated with Aβ deposition and tau hyperphosphorylation in the brains of SAMP8 mice
The other feature of AD is the formation of NFTs, which is the result of Ttau hyperphosphorylation. Tau hyperphosphorylation is another major pathological feature in the brain of SAMP8 mice similar to AD patients [43]. Tau hyperphosphorylation was detected in the cortex, striatum, and hippocampus of SAMP8 mice as early as 3 months of age compared to age-matched SAMR1 mice [30]. The hyperphosphorylation sites of tau in SAMP8 mice include Ser396 and Ser404 [67–69], Ser202/Thr205 [70], Ser262 [71], Ser199 [72], Ser212 [30], Ser422 [17], and Thr668 [73]. The hyperphosphorylation of tau is related to a variety of enzymes in the brain of SAMP8 mice (Table 2), which are closely correlated with each other (Fig. 2). Studies have shown that the regulation of the expression or activity of these enzymes can effectively reduce the hyperphosphorylation level of tau, improve the cognitive impairment and brain aging process of SAMP8 mice. Glycogen synthase kinase-3 (GSK3) and cyclin-dependent kinase 5 (CDK5) are two important enzymes involved in tau phosphorylation. GSK3β is a subtype of GSK3, which is the molecular link between Aβ and tau. Activation of GSK3β is dependent upon the phosphorylation of Tyr216, and inactivation of GSK3β is dependent upon phosphorylation GSK3β at Ser9. Activation of GSK3β is responsible for 88% of tau phosphorylation [74]. Previous studies showed that there was no difference in GSK3β activity in the brain of SAMP8 mice compared with age-matched SAMR1 [75]. However, our unpublished data and some recent studies found that in addition to increased levels of Cdk5 in the brain of SAMP8 mice, the activity of GSK3β also increased [25, 77], eventually leading to tau hyperphosphorylation and NFTs. Drugs, which can interference with the expression and activities of these enzymes or proteins, can effectively reduce the hyperphosphorylation of tau. Additionally, proteins such as p35, p25 [73], calpain [78], calmodulin-dependent protein kinase (CaMKII) [66], protein kinase C (PKC) [79], and NMDA receptor (NMDAR) [68, 80] were increased in SAMP8 mouse brain. Signaling pathways such as PI3K/Akt/mTOR, ERK, JNK, and P38 MAPK are also involved in regulating the phosphorylation of tau in SAMP8 mice (see Table 2 and Fig. 2).
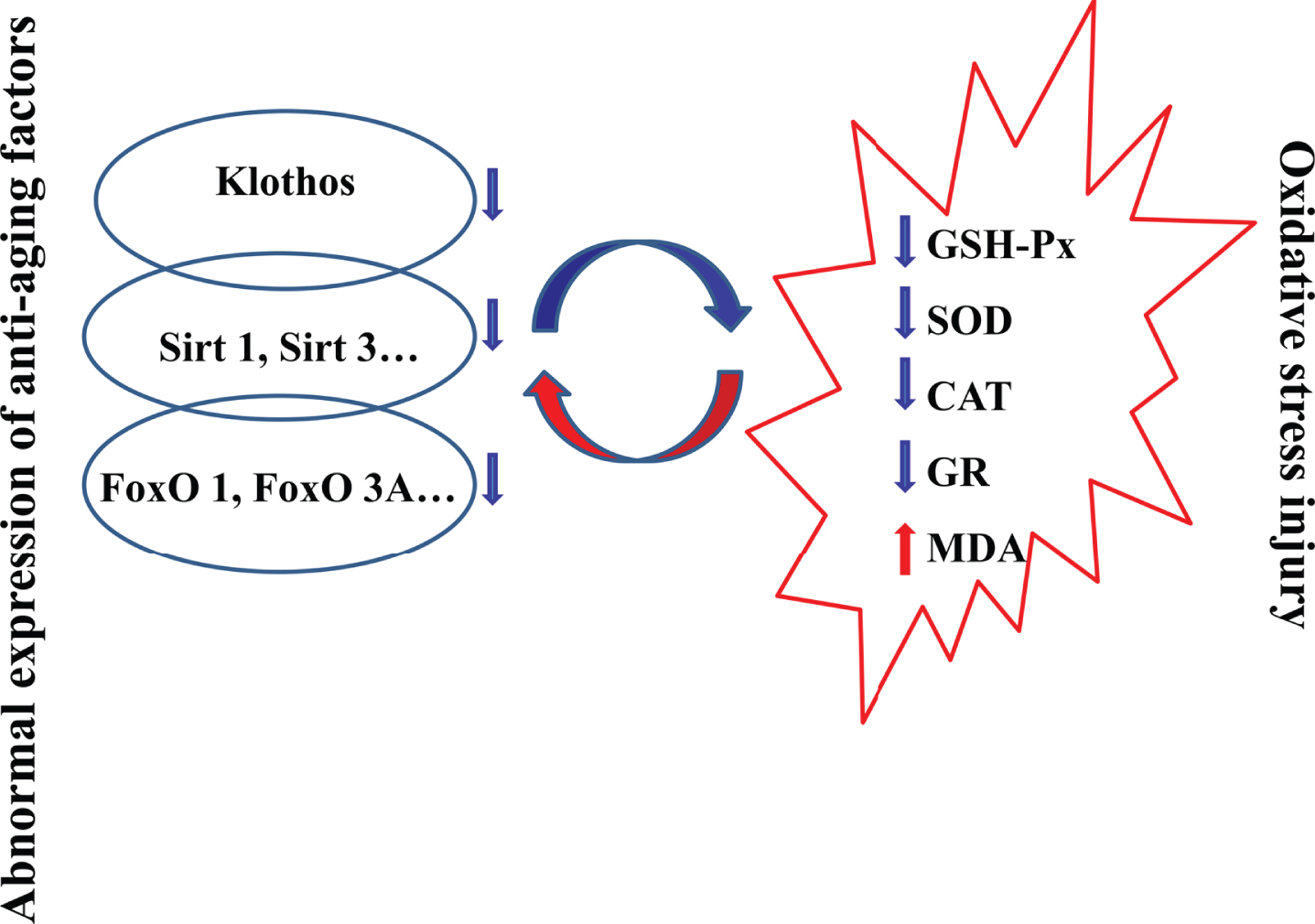
Interaction between aging-inhibiting factors and oxidative stress injury in brain of SAMP8 mice. GSH-Px, glutathione peroxidase; SOD, superoxide dismutase; CAT, catalase; GR, glutathione reductase; MDA, malondialdehyde; Sirt, silent information regulator type; FoxOs, Forkhead box class O.

Protein expression and related signaling pathways of tau hyperphosphorylation in SAMP8 mice compared with age-matched SAMR1 mice or younger SAMP8 mice. PP2A, protein phosphatase-2A; CaMKII, Calcium/calmodulin dependent protein kinase II; PKC, protein kinase C; N-NMDAR, N-methyl-D-aspartate receptor; GSK3β, glycogen synthase kinase-3β; Cdk5, cyclin-dependent kinase 5; PI3K, phosphatidyl inositol 3-kinase; Akt, protein kinase B; mTOR, mammalian target of rapamycin; ERK, extracellular regulated protein kinases; JNK, c-Jun N-terminal kinase; P38 MAPK, p38 mitogen-activated protein kinase.
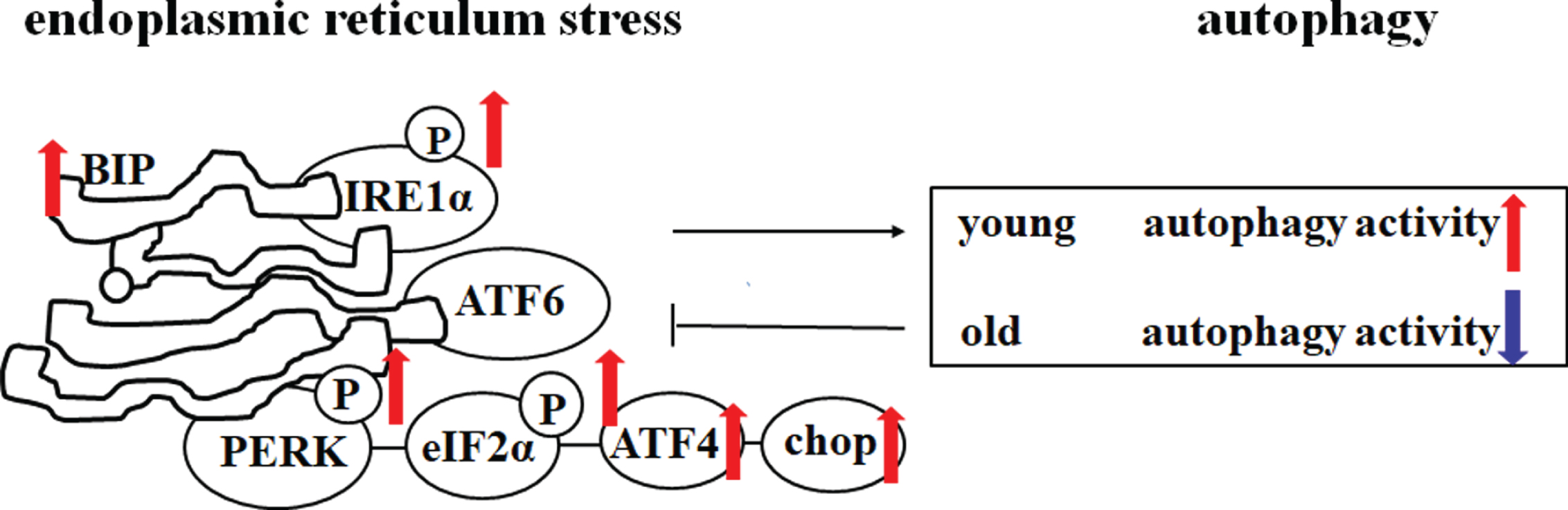
Changes in endoplasmic reticulum stress and autophagy activity in SAMP8 mice compared with age-matched SAMR1 mice or younger SAMP8 mice. PERK, phosphoprotein kinase RNA-like endoplasmic reticulum kinase; eIF2α, eukaryotic initiation factor 2α; ATF4, activating transcription factor-4; CHOP, C/EBP homologous protein; ATF6, activating transcription factor 6; BIP, immunoglobulin heavy chain binding protein; IRE1α, inositol requiring enzyme 1α.
Endoplasmic reticulum stress and autophagy
Endoplasmic reticulum (ER) is the main organelle for protein biosynthesis and transport [90, 91]. When the unfolded or misfolded protein load exceeds the capacity of the ER to properly fold protein, the sensor protein on the ER membrane is activated to trigger the downstream signal transduction. This activation of a specific cellular process called endoplasmic reticulum stress (ERS) or unfolded protein response (UPR). ERS mainly includes three signaling pathways, namely PECK/eIFa/ATF4, ATF6, and IRE1a/XBP1s. ERS is known to induce apoptosis and cell death; moreover, ERS is closely related to the aging process [92]. In SAMP8 mice, ERS plays a key role in many age-related metabolic dysfunctions and is a significant cause of cardiac dysfunction, hepatic dysfunction, and adipose tissue dysfunction [13, 94]. Meanwhile, ERS also plays an important role in brain aging in SAMP8 mice [95]. ERS in neurons can be caused by oxidative stress, calcium imbalance, Aβ deposits, and tau hyperphosphorylation. What is worse, ERS and tau hyperphosphorylation can create a vicious cycle that leads to the death of neurons [96]. In SAMP8 mice, immunoglobulin heavy chain-binding protein (BIP), an ER stress marker, was increased in the brain. ERS-related proteins PERK, eIF-2α, and IRE-1α were increased compared with age-matched SAMR1 mice [97]. Our research group found that, under electron microscopy, the ER in the hippocampal neurons of SAMP8 mice was significantly swollen, further supporting that ERS plays an important role in the brain aging of SAMP8 mice. Increasing evidence suggests that in chronic neurodegenerative diseases, persistent ERS often induces autophagy, which may be a compensatory mechanism to reduce ERS [98].
Autophagy is a widely studied pathway to degrade or eliminate damaged cell organelles and proteins. However, autophagy is a double-edged sword, excessive induction of autophagy may lead to neuronal cell death [99]. Numerous autophagic vacuoles (AVs) accumulate within the hippocampus of AD patients [100]. Dysregulation of the autophagy lysosomal pathway was observed in the brain of SAMP8 mice as early as 5 months of age [101]. Autophagy markers were also increased in the brain of SAMP8 mice at 6, 7, and 10 months of age [41, 103]. Consistent with previous studies, our recent study found that a large number of autophagosomes were formed in the hippocampus of SAMP8 mice at the age of 8 months under electron microscopy, and the expression of autophagy markers p62 and LC3-II in the hippocampus and cortex of SAMP8 mice was also significantly increased [21]. However, autophagy activity was changed in the brain of SAMP8 mice with aging. Our research group found that autophagy markers Beclin-1 and LC3-II were significantly decreased in the hippocampus and cortex of SAMP8 mice at 12 months of age, compared with age-matched SAMR1 mice, indicating that autophagy was decreased in SAMP8 mice at 12 months of age [102]. Thus, it should be noted that autophagy in the brain of SAMP8 mice changes with the age, autophagy activity increased in the early stage, and decreased in the late stage of AD.
Taken together, ERS is enhanced in the brain of young and mature SAMP8 mice, which then induces the increase of autophagosomes. At this time, effective inhibition of ERS significantly improved cognitive disorder in SAMP8 mice [97]. Increased autophagy in early aging of SAMP8 mice suggests that autophagy can act as a compensatory mechanism to reduce ERS, and activation of autophagy can improve learning and memory disorders [21, 105]. However, with the progress of the aging, autophagy activity in the brains of 12-month-old SAMP8 mice decreased, which further exacerbated ERS [67]. At this time, enhancing autophagy could effectively delay the brain aging process. Nevertheless, the changes of autophagy activity with the age in SAMP8 mice need to be further studied.
Intestinal microbiota
In recent years, the role of intestinal microbiota in AD has received extensive attention. Gut microbes are parasitic in the intestinal tract in a relatively constant proportion for a long time and participate in the metabolism of nutrients and immune regulation. Dynamic balance and stability of intestinal microbiota play an important role in maintaining body health [106]. During the aging process, the composition and function of intestinal microbiota change, and beneficial bacteria gradually decrease, thus affecting the health of the host. Imbalance of intestinal flora can lead to high blood glucose, and increase intestinal permeability and inflammation [107]. More notably, there is a brain-gut axis between the intestine and the brain, and the homeostasis of the intestinal flora is closely related to neurodegenerative disease such as AD [108]. Further studies of intestinal flora in AD pathogenesis become a hot topic and help anti-AD drug development [109].
The behavioral differences between SAMP8 and SAMR1 mice were closely related to composition, diversity, and abundance of intestinal flora. For example, α-diversity (Chao 1, Shannon, and Simpson indices), β-diversity (Principal coordinates analysis, Bray-Curtis dissimilarity), and principle component analysis (PCA) revealed quite different patterns between SAMP8 and SAMR1 mice, and at the Phylum, Class, Order, Family, and Genus levels, at least 27 significant differences were detected between SAMP8 and SAMR1 mice [110]. Treatment of 6-month-old SAMP8 mice with a Chinese herb medicine preparation (LW-AFC) for 5.5 months significantly improved cognitive decline of 11.5-month-old SAMP8 mice, together with microbiome modifications, including restored 15 altered bacteria in SAMP8 mice to the levels similar to SAMR1 mice [23]. Baicalein, a flavonoid from Chinese medicine Scutellariae baicalensis, was administered to 8-month-old SAMP8 mice for 8 weeks, significantly improved cognitive decline and reduced neuroinflammation, but importantly, altered the abundance of 6 gerena in SAMP8 mice, which correlated with cognitive impairments (e.g., Chritensenellaceae), and neuroinflammation (e.g., Prevotella) [111]. In elderly patients, a moderate-fat diet enriched with fish oil showed beneficial effects, and such dietary supplements given to SAMP8 mice from 6 to 15 months of age improved intestinal bacteria disruption and brought the increased bacteria in association to fatty liver and inflammation to baseline [112]. Regulation of intestinal flora by drugs, diets, or fecal microbiota transplantation can improve cognitive dysfunction in SAMP8 mice, indicating the importance in targeting gut microbiota as a novel therapeutic strategy [110, 113].
PROSPECT
In summary, age-related learning and memory impairment in SAMP8 mice are closely related to abnormal expression of anti-aging factors, oxidative stress injury, inflammation, Aβ deposition, tau hyperphosphorylation, ERS, autophagy, and disruption of metabolism and intestinal microbiota. These mechanisms act together in an integrated manner to promote rapid aging of SAMP8 mice. These age-dependent pathologies resemble those found in AD patients, either at early onset, or at the late stage of AD. The molecular mechanisms and gut microbiota in this review could be novel therapeutic targets for anti-aging therapy development to protect elderly human health.
Footnotes
ACKNOWLEDGMENTS
This study was financially supported by the National Natural Science Foundation of China, Grant Number: 81773739; Guizhou Province Synergy Innovation Center, Grant Number: CJ-926; Guizhou Provincial Department of Education “125” Major Science and Technology Projects, Grant Number: 2012012; Guizhou Provincial Department of Education Tutor Studio, Grant Number: 99-030.