
Abstract
Background:
Inflammation and oxidative stress are believed to play an important role in the pathogenesis of Alzheimer’s disease (AD). Tenuifolin (TEN) is a natural neuroprotective compound extracted from Polygala tenuifolia Willd, which may improve cognitive symptoms.
Objective:
This study was designed to evaluate the protective effect of TEN on inflammatory and oxidative stress induced by amyloid-β (Aβ)42 oligomers in BV2 cells, and to explore the underlying mechanisms.
Methods:
We conducted cell viability assays to estimate drug toxicity and drug effects on cells. Quantitative real-time polymerase chain reaction and enzyme-linked immunosorbent assays were performed to detect the release of inflammatory factors. Nitric oxide (NO) assays were used to measure the degree of oxidative stress. Western blot and immunofluorescence analysis were used to explore the influence of TEN on the nuclear factor-κB (NF-κB) pathway.
Results:
Pretreatment of BV2 microglial cells with TEN inhibited the release of tumor necrosis factor-α, interleukin-6, and interleukin-1β, alleviated NO-induced oxidative stress by inhibiting the expression of inducible nitric oxide synthase and cyclo-oxygenase-2, and protected SH-SY5Y cells from the toxicity induced by the medium conditioned by BV2 cells previously exposed to Aβ42 oligomers. Moreover, TEN suppressed upstream activators of NF-κB, as well as NF-κB translocation to the nucleus in BV2 microglial cells.
Conclusion:
This study demonstrates that TEN can protect SH-SY5Y cells from Aβ42 oligomer-induced microglia-mediated inflammation, and oxidative stress by downregulating the NF-κB signaling pathway.
INTRODUCTION
Alzheimer’s disease (AD) is one of the most common chronic degenerative diseases and a major health issue in the elderly population. AD pathogenesis is complex and remains poorly understood. To date, the results of international clinical trials have been mostly unsatisfactory. AD pathogenesis is believed to involve the amyloid-β (Aβ) cascade and the hyperphosphorylation of tau protein [1]. Moreover, inflammatory response and glial hyperplasia are common features of AD [2]. Previous studies have suggested that chronic inflammation, which manifests as a continuous activation of glial cells, is a response to Aβ and neurofibrillary tangles [3]. Although the initial inflammatory response may be beneficial, chronic activation of astrocytes and microglia induces injury via releasing cytokines, chemokines, complement factors, pattern-recognition receptors, as well as cellular and molecular immune factors [4]. Activated astrocytes and microglia, surrounded by Aβ plaques, appear in the proximity of neurons and initiate the inflammatory response by producing inflammatory factors and dysregulating cellular signaling pathways [5, 6]. The existence of this mechanism is supported by the fact that several pro-inflammatory cytokines are commonly expressed in the brains of patients with AD but not in the normal brains [7, 8].
Polygala tenuifolia Willd is a traditional Chinese herbal medicine and has a long history of therapeutic use. Its use has been implicated in treatment of various neuropsychiatric disturbances such as memory decline, sleeping disorders, depression, and others [9, 10]. Tenuifolin (TEN) is one of the major active ingredients of Polygala tenuifolia Willd (Fig. 1). Earlier studies showed that TEN protects PC12 and SY5Y cells from apoptosis induced by Aβ25–35, and significantly improves cognitive dysfunction caused by intracellular injection of Aβ in mice [9]. Moreover, it was found that TEN reduces Aβ production by inhibiting β-secretase [11] and improves cognition in aging mice by increasing the levels of norepinephrine and dopamine in the hippocampus, while reducing acetylcholinesterase activity in the cortex [12]. Recently, Yang et al. found that TEN can reverse spatial learning, memory disorders, and neuronal apoptosis in the hippocampus of APP/PS1 transgenic AD mice [13]. However, so far, no studies have clarified the underlying mechanism of beneficial effects exerted by TEN beneficial in AD models.
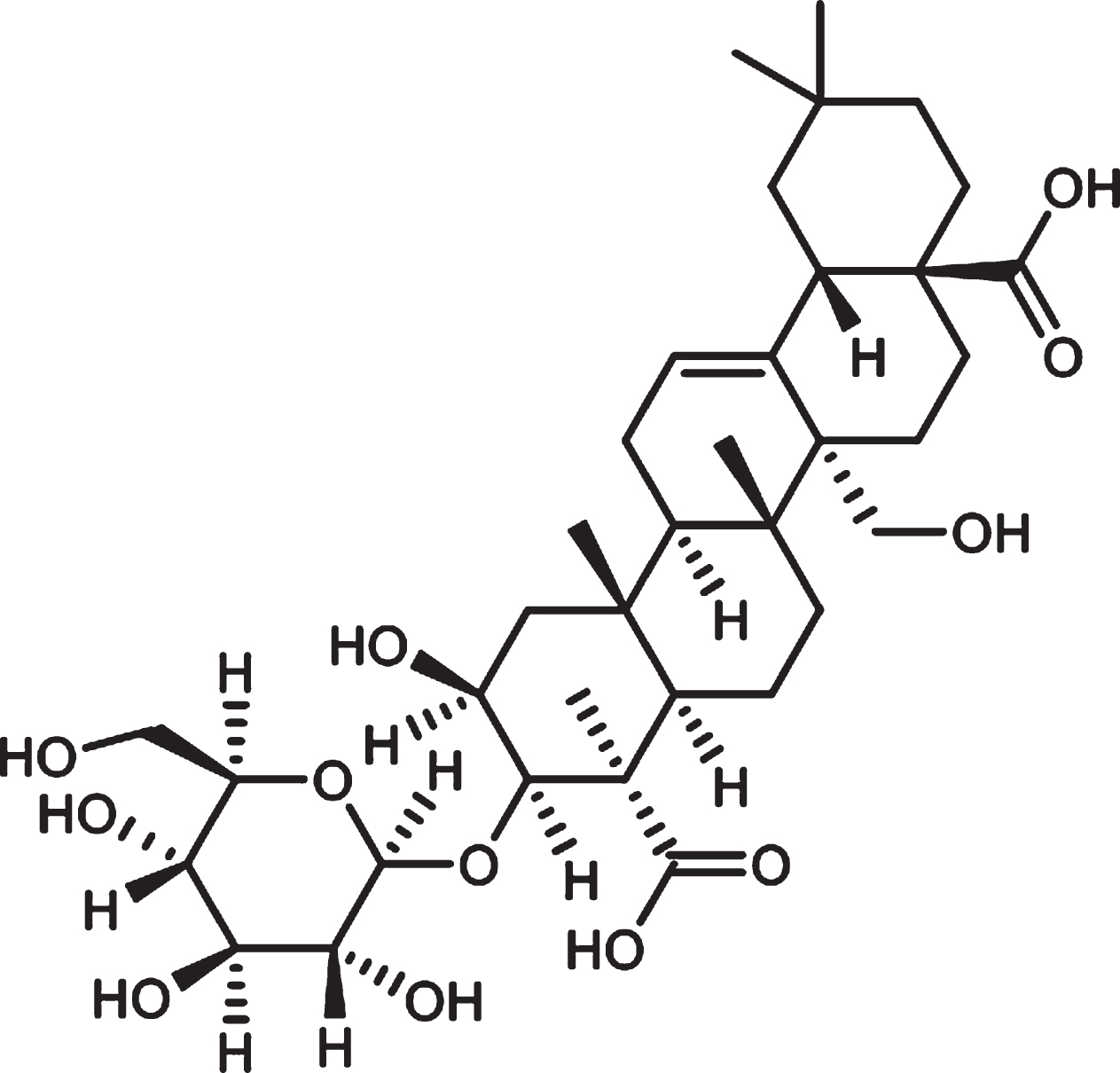
Chemical structure of Tenuifolin.
Aβ42 oligomers are more neurotoxic than Aβ fiber aggregates, and low concentrations of Aβ42 oligomers directly cause neuronal damage [14, 15]. Therefore, in the present research, cellular models of AD were generated by utilizing Aβ42 oligomers. Our data provides evidence for the possible use of TEN as a novel therapeutic modality for patients with AD.
MATERIALS AND METHODS
Cell culture and reagents
Neuron-like SH-SY5Y cells and microglia-like BV2 cells were purchased from the National Infrastructure of Cell Line Resource. Cell lines were cultured in complete medium containing Dulbecco’s Modified Eagle’s Medium (DMEM)/Ham’s F-12 (1 : 1) medium or DMEM (Invitrogen) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin (Sigma-Aldrich). Both The cells types were maintained at 37°C in a humidified 5% CO2 incubator. TEN (purity > 99%) was purchased from the National Institutes for Food and Drug Control. Aβ42 oligomers were procured from ChinaPeptides Co., 3-(4,5-dimethylth iazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) and prostratin (purity≥98%) were purchased from Sigma-Aldrich.
MTT assays
MTT assays were performed to assess the toxicity of Aβ42 oligomers and TEN in BV2 cells. Briefly, BV2 cells were seeded into 96-well culture dishes (5×103 cells per well) and incubated for 24 h. The cells were treated with various concentrations of Aβ42 oligomers (0.1, 1, 5, 10, and 20μM) or TEN (1, 5, and 10μM) for another 24 h. MTT assay was used to test the toxicity of the supernatant from Aβ42 oligomer-stimulated microglia on SH-SY5Y cells. Briefly, BV2 cells were treated with 5μM Aβ42 oligomers for 6 h or pretreated with TEN (1, 5, or 10μM) for 2 h before Aβ exposure. Next, the Aβ-containing medium was replaced with fresh medium and the stimulated BV2 cells were cultured for another 12 h prior to collecting the supernatant. The conditioned medium was added to SH-SY5Y cells previously seeded in 96-well culture dishes (1×104 cells per well) and further incubated for 24 h. Phosphate-buffered saline (PBS) was used to wash the cells twice before adding MTT solution (5 mg/mL) into each well. Cells were incubated with the MTT reagent for 3 h at 37°C. The solution was removed from each well, and blue crystals were dissolved in DMSO. The absorbance was measured at a wavelength of 490 nm by a spectrophotometer (Thermo Fisher, Vantaa, Finland).
CCK-8 assays
The viability of SH-SY5Y cells was also measured with a cell counting kit-8 (CCK-8) assay (Nanjing Jiancheng Bioengineering Institute). The conditioned medium of BV2 was obtained as mentioned above and was added to SH-SY5Y cells previously seeded in 96-well culture dishes (1×104 cells per well). After 24 h of treatments, CCK-8 solution was added to each well to a final concentration of 0.5 mg/mL and the cells were incubated at 37°C for another 3 h. The absorbance of the samples was measured at a wavelength of 450 nm with the spectrophotometer.
qRT-PCR
BV2 cells cultured at 5×105 cells per well in 6-well plates were treated with TEN (5μM or 10μM) for 2 h prior to stimulation with Aβ42 oligomers (5μM) for 6 h. Total RNA was extracted using the Animal Total RNA Isolation kit (Sangon Biotechnology) and reverse-transcribed by using PrimeScript™ RT reagent Kit with gDNA Eraser (TaKaRa Biotechnology). cDNA was amplified with TB Green® Premix Ex Taq™ II (TaKaRa Biotechnology) in a final volume of 20μl. Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using the Step One Plus Real-time PCR System (Applied Biosystems, Foster City, USA). GAPDH was used as a reference gene to normalize the results. PCR amplification was performed using the following program: 95°C for 30 s, followed by 40 cycles of 95°C for 5 s, 60°C for 30 s, and 95°C for 15 s. The results were quantified by using SDS software (Applied Biosystems, Foster City, USA) and expressed as fold-changes. The sequences of oligonucleotide primers are shown in Table 1.
Sequences of primers used for qRT-PCR
NO assay
BV2 cells were seeded into 96-well plates at a density of 1×104 cell per well and cultured for 12 h. After treatment with Aβ42 oligomers, alone or with 5μM or 10μM TEN for another 12 h, the supernatant was collected and the NO content was measured by the Griess method by following the manufacturer’s instructions of the NO assay Kit (Nanjing Jiancheng Bioengineering Institute). The optical density was analyzed at 540 nm after a 10 min incubation at room temperature.
ELISA
BV2 cells were plated in 6-well culture plates (5×105 cells per well) in the culture medium. For the measurement of TNF-α, IL-1β, and IL-6 protein expression, BV2 were pretreated with 5μM or 10μM TEN for 2 h and then stimulated with 5μM Aβ42 oligomers for 24 h. The pro-inflammatory factors in the cell medium were assayed using the enzyme-linked immunosorbent assay (ELISA) Kits (CUSABIO Technology) according to the manufacturer’s instructions.
Western blot
BV2 cells were cultured in 6-well plates (5×105 cells per well) and treated with Aβ42 oligomers in the presence or absence of TEN (5μM or 10μM) for 1 h. The protein concentrations were determined using the BCA Protein Assay Kit (Beyotime Biotech). The nuclear and cytosolic proteins were extracted and isolated on demand according to the manufacturer’s instructions of the Nuclear and Cytoplasmic Protein Extraction Kit (Sangon Biotechnology). Protein samples were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and electro-transferred onto polyvinylidene fluoride (PVDF) membranes. The membranes were incubated overnight at 4°C with the following primary antibodies: IKK (1 : 1000), p-IKK (1 : 1000), IκB (1 : 1000), p-IκB (1 : 1000), NF-κB (p65) (1 : 1000), p-NF-κB (p-p65) (1 : 1000), β-actin (1 : 2000), and Histone H3 (1 : 1000) purchased from Cell Signaling Technology. The membranes were washed three times with Tris-HCl buffer and 0.1% Tween 20 (TBST), and incubated with secondary antibodies at room temperature for 2 h. Finally, the protein expression bands were quantified using Image J (Image J version 1.51e, National Institutes of Health, Maryland, USA).
Immunofluorescence staining
BV2 cells were seeded (1×104 per well) into poly-D-lysine-coated 24-well plates. The cells were pre-treated with or without 5μM TEN for 2 h before adding 5μM Aβ42 oligomers. After incubation for 24 h, the cells were fixed in cool paraformaldehyde for 15 min and then treated with 0.3% Triton X-100 for 20 min. After blocking in goat serum for 30 min, the cells were incubated overnight at 4°C with primary anti-NF-κB p65 antibody (1 : 50) (Cell Signaling Technology). The cells were incubated with Alexa Fluor 594 goat anti-mouse IgG (1 : 500) (Invitrogen Corporation) for 1 h, and 4,6-diamino-2-phenyl indole (DAPI) was used to label the nuclei. The images were acquired by using a fluorescence microscope (Leica DM4 B, Leica Microsystems Inc., Buffalo Grove, USA).
Statistical analysis
All data are expressed as the mean±standard deviation (SD) of three independent experiments. Differences between groups were analyzed by one-way analysis of variance (one-way ANOVA) followed by Bonferroni post hoc tests. Results with p values <0.05 were considered statistically significant. Statistical analysis was performed by SPSS 19.0 (IBM SPSS Statistics for MacOSX, Version 19.0, IBM Corp, NY, USA). GraphPad Prism 7.0 (GraphPad Software, Inc., La Jolla, CA, USA) was used to analyze all data and images.
RESULTS
TEN alleviates Aβ42-induced BV2 neurotoxicity
MTT assays for Aβ42 oligomers showed no significant toxicity was induced by 5μM Aβ42 oligomers, while 10μM and 20μM Aβ42 exerted toxic effects after a 24 h incubation (Fig. 2A, p > 0.05 for 5μM, p = 0.0002 for 10μM, p < 0.0001 for 20μM). In order to make the Aβ42 in the inflammation model produce a stimulating effect on BV2 cells instead of an obvious cytotoxic effect, we used 5μM Aβ42 oligomers in subsequent experiments. MTT assays for TEN indicated that no significant impact on BV2 cell viability was observed within a relatively wide range of TEN concentrations (Fig. 2B, all p > 0.05). Next, the medium of BV2 cells that had been previously treated with 5μM Aβ42 oligomers for 6 h was collected and the conditioned medium was incubated with SH-SY5Y cells for 24 h, and MTT assay was performed to evaluate toxicity. The exposure to the conditioned medium alone significantly decreased the viability of SH-SY5Y cells (Fig. 2C, p = 0.0003). However, neurotoxicity was inhibited when SH-SY5Y cells were treated with the medium of BV2 cells that had been pre-incubated for 2 h with 5μM or 10μM TEN before Aβ42 stimulation (Fig. 2C, p = 0.0093, p = 0.0020, respectively). Moreover, the results of CCK-8 assays further confirmed the cell protection effect of TEN. As shown in Fig. 2D, the conditioned medium which pretreated with 5μM or 10μM TEN reduced the cytotoxicity of BV2 cell release on SH-SY5Y (p = 0.0093, p = 0.0020, respectively).
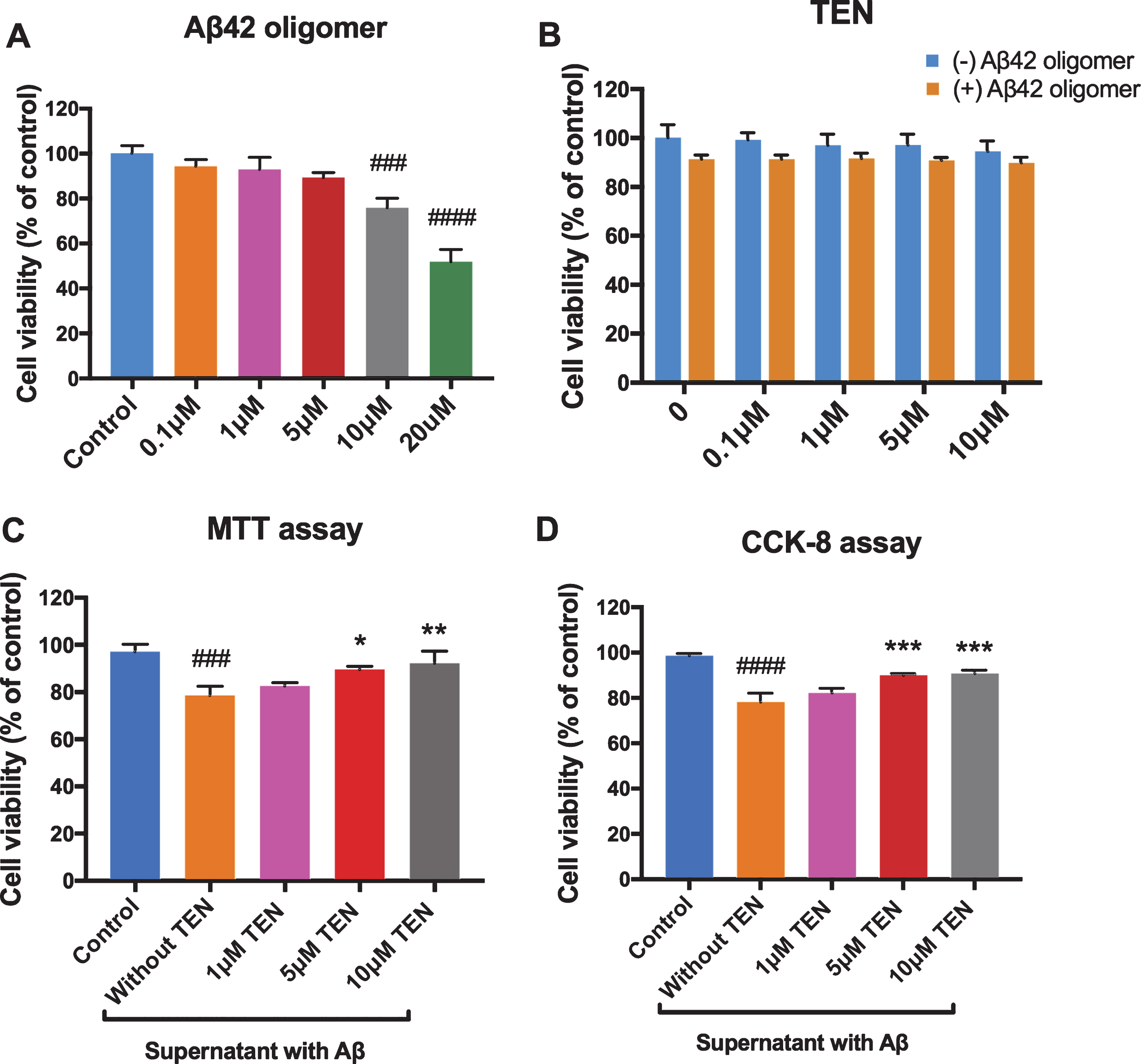
Effect of Aβ42 oligomers and TEN on cell viability. A) Cell viability of BV2 cells analyzed by MTT assays subjected to Aβ42 oligomers stimulation for 24 h. B) Cell viability of BV2 analyzed by MTT assays subjected to TEN treatment with or without 5μM Aβ42 oligomers stimulation for 24 h. C, D) Cell viability of SH-SY5Y analyzed by MTT or CCK-8 assays subjected to the conditioned medium. # # # p < 0.001, # # # # p < 0.0001 versus control, * p < 0.05, ** p < 0.01, *** p < 0.001 versus Aβ or supernatant with Aβ, n = 4 per group.
TEN attenuates Aβ42 oligomer-induced inflammatory reactions in BV2 cells
Notably, when cells were pretreated with 5μM TEN before Aβ42 stimulation, the mRNA levels of TNF-α, IL-1β, and IL-6 were 60.42%, 68.41%, and 60.70%, respectively, of the corresponding levels in cells treated with Aβ alone (Fig. 3A–C, p < 0.0001, p = 0.0028, p = 0.0005, respectively). In contrast, when TEN was used at 10μM, the mRNA levels of TNF-α, IL-1β, and IL-6 were further reduced to 47.09%, 58.60%, and 42.50%, respectively (Fig. 3A–C, p < 0.0001, p = 0.003, p < 0.0001, respectively). ELISA assays showed that while the levels of these cytokines were relatively low and difficult to detect in control cells, they were overexpressed after treatment with Aβ42 oligomers. The treatment with 5μM or 10μM TEN inhibited the upregulation of TNF-α and IL-6 induced by Aβ42 oligomers (Fig. 3D, E). It should be mentioned that no statistically significant differences were observed between TEN-treated cells and the control cells, as determined by qRT-PCR and western blot analyses.
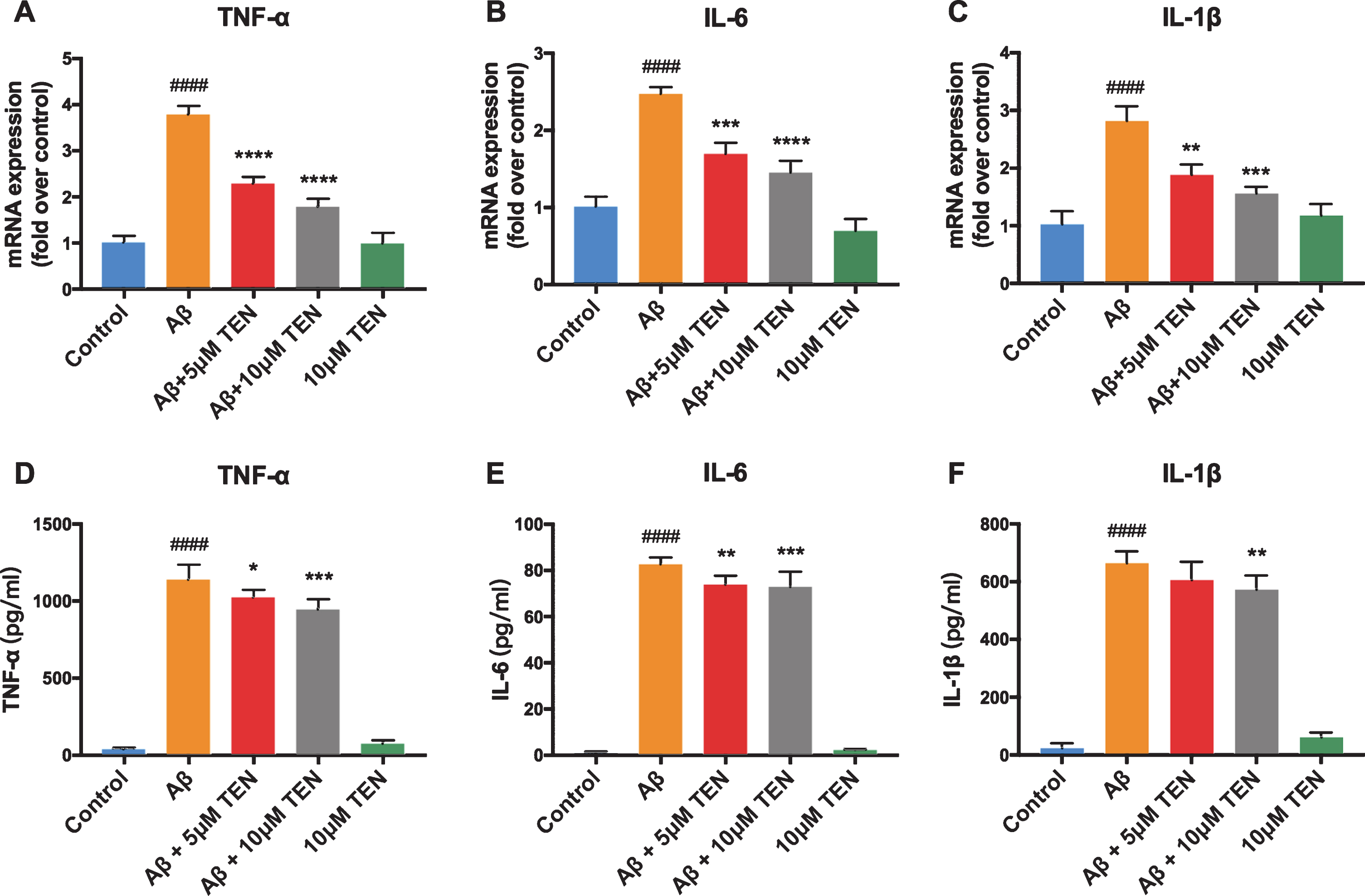
Effect of TEN on the production of inflammatory factors in Aβ42 oligomers-stimulated BV2 cells. A–C) The expression of TNF-α, IL-6, and IL-1β mRNA after treatment with 5μM Aβ42 oligomers alone or with TEN for 6 h. D–F) The production of TNF-α, IL-6 and IL-1β protein after treatment with 5μM Aβ42 oligomers alone or with TEN for 24 h. # # # # p < 0.0001 versus control, * p < 0.05, ** p < 0.01, *** p < 0.001, ****p<0.0001 versus Aβ, n = 3 for qRT-PCR, n = 6 for ELISA.
TEN inhibits Aβ42 oligomer-induced oxidative stress
The pretreatment with 5μM TEN prior to Aβ42 exposure decreased the level of iNOS mRNA by 26.07% (Fig. 4A, p = 0.01), and of COX-2 mRNA by 4.02% (Fig. 4B, p > 0.05). The pretreatment with 10μM TEN reduced iNOS and COX-2 to 34.84% and 20.16%, respectively, compared to the treatment with Aβ42 alone (Fig. 4A, B, p = 0.0016, p = 0.0223, respectively). In addition, no significant difference was found between the two TEN concentration groups, and TEN treated alone did not increase the amount of iNOS and COX-2 mRNA significantly. TEN pretreatments at 5μM or 10μM reduced NO release by 19.59% and 29.42%, respectively (Fig. 4C, p = 0.0498, p = 0.0031, respectively). Treatment with TEN alone did not significantly affect the level of extracellular NO, which was comparable to that of control cells (Fig. 4C, p > 0.05). Finally, no significant differences were observed between cells treated with TEN alone at the two concentrations (Fig. 4C, p > 0.05).
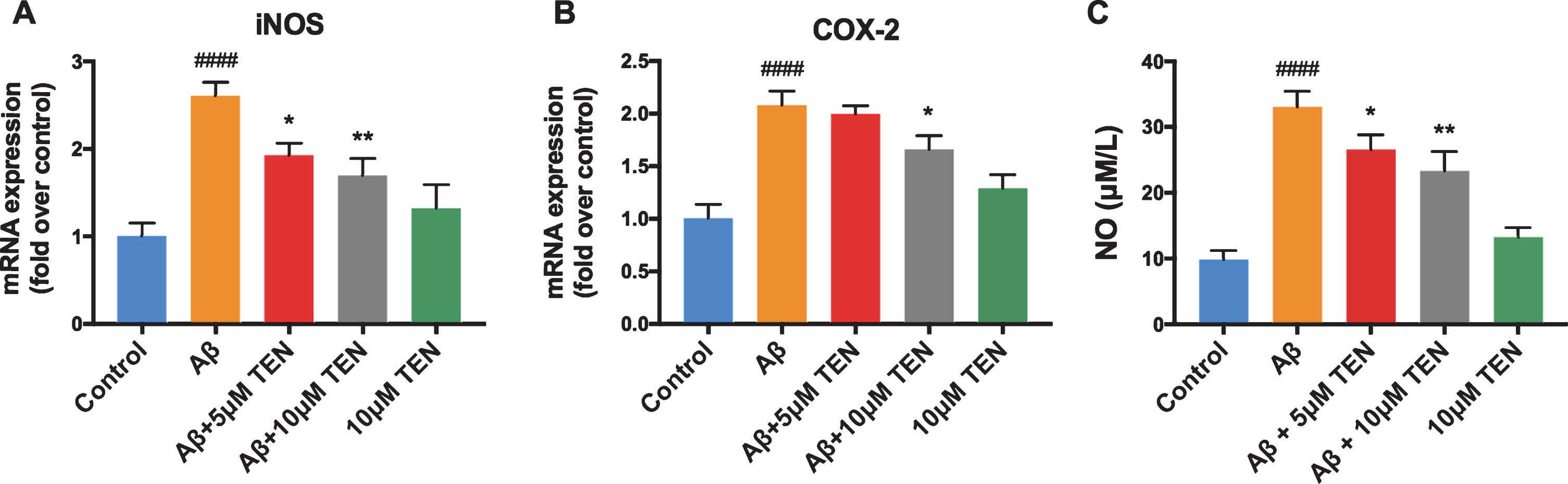
Effect of TEN on the oxidative stress induced by Aβ42 oligomers in BV2 cells. A, B) The expression of iNOS and COX-2 mRNA after treatment with 5μM Aβ42 oligomers alone or with TEN for 6 h. C) The production of NO in supernatants after treatment with 5μM Aβ42 oligomers alone or with TEN for 12 h. # # # # p < 0.0001 versus control, * p < 0.05, ** p < 0.01 versus Aβ, n = 3 per group.
TEN suppresses the activation of the NF-κB signaling pathway by preventing the phosphorylation of IKKβ, IκB, and NF-κB
As shown in Fig. 5, Aβ42 oligomers remarkably increased the phosphorylation of IKKβ, IκB, and NF-κB (p65) compared to controls. However, pretreatment with TEN partially prevented the activation of the two crucial upstream regulators of NF-κB, i.e., IKK and IκB, compared to the Aβ group, thereby suppressing the phosphorylation of NF-κB (p65). TEN could only significantly decrease Aβ42-induced NF-κB upregulation at the 5μM concentration (p = 0.0423), while 10μM TEN inhibited the IKK, IκB, and NF-κB upregulation by 43.08%, 52.46%, and 57.49%, respectively (Fig. 5B, p = 0.0235, p = 0.0311, p = 0.0100, respectively). When adding prostratin (10μg/ml), an activator of NF-κB signaling, the inhibitory effects of 10μM TEN on IKK, IκB, and NF-κB phosphorylation were attenuated.
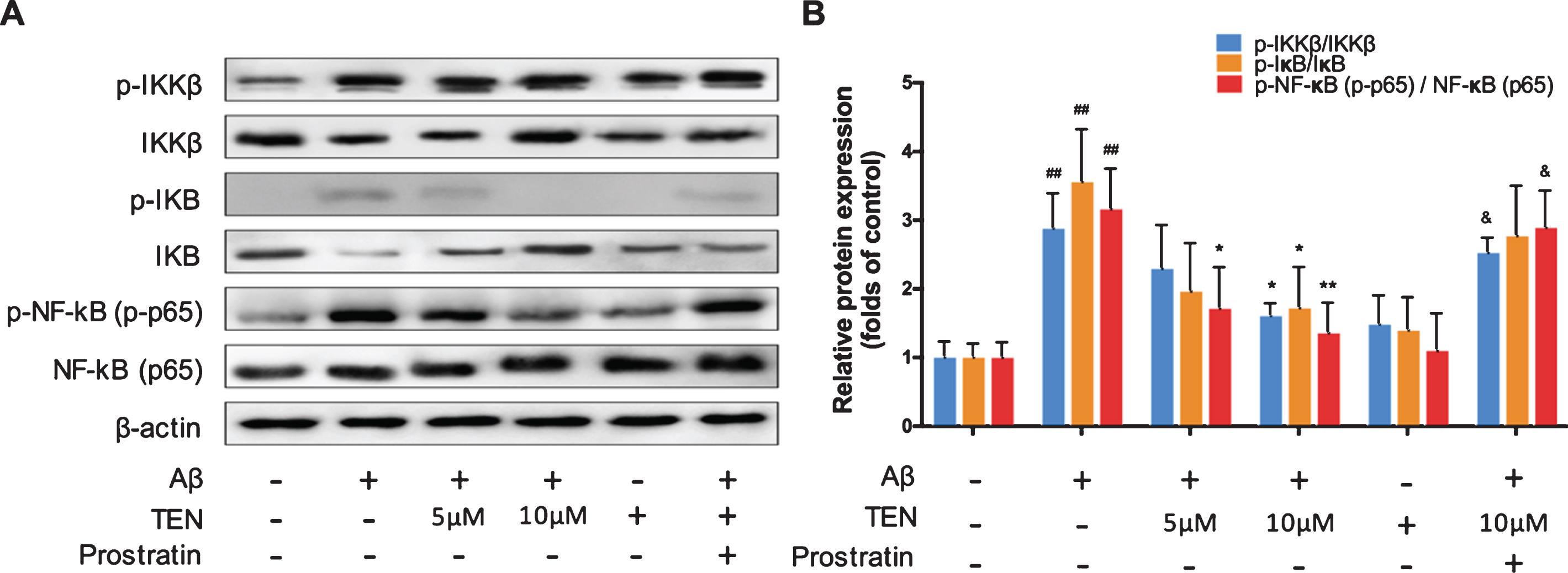
Effects of TEN treatment on Aβ42 oligomers-induced NF-κB pathway activation in BV2 cells. A, B) Western blots and analysis NF-κB signaling pathways proteins after treatment with 5μM Aβ42 oligomers alone or with TEN or with 10μg/ml prostratin for 1 h. # # p < 0.01 versus control, * p < 0.05, ** p < 0.01 versus Aβ, & p < 0.05 in comparison with Aβ+ 10μM TEN, n = 3 per group.
TEN inhibits NF-κB (p65) nuclear translocation in Aβ-activated BV2 microglia
The western blot results showed that after stimulation with 5μM Aβ42 oligomers, the content of NF-κB in the cytoplasm of BV2 cells decreased significantly (Fig. 6A,B, p = 0.0067), while its nuclear level increased (Fig. 6A,B, p = 0.0375); however, 5μM TEN reversed this phenomenon. Immunofluorescence staining confirmed these results (Fig. 6C). In conclusion, TEN inhibited the Aβ42 oligomer-induced activation of NF-κB in BV2 cells and suppressed NF-κB transport to the nucleus, thereby inhibiting its downstream signaling pathway.
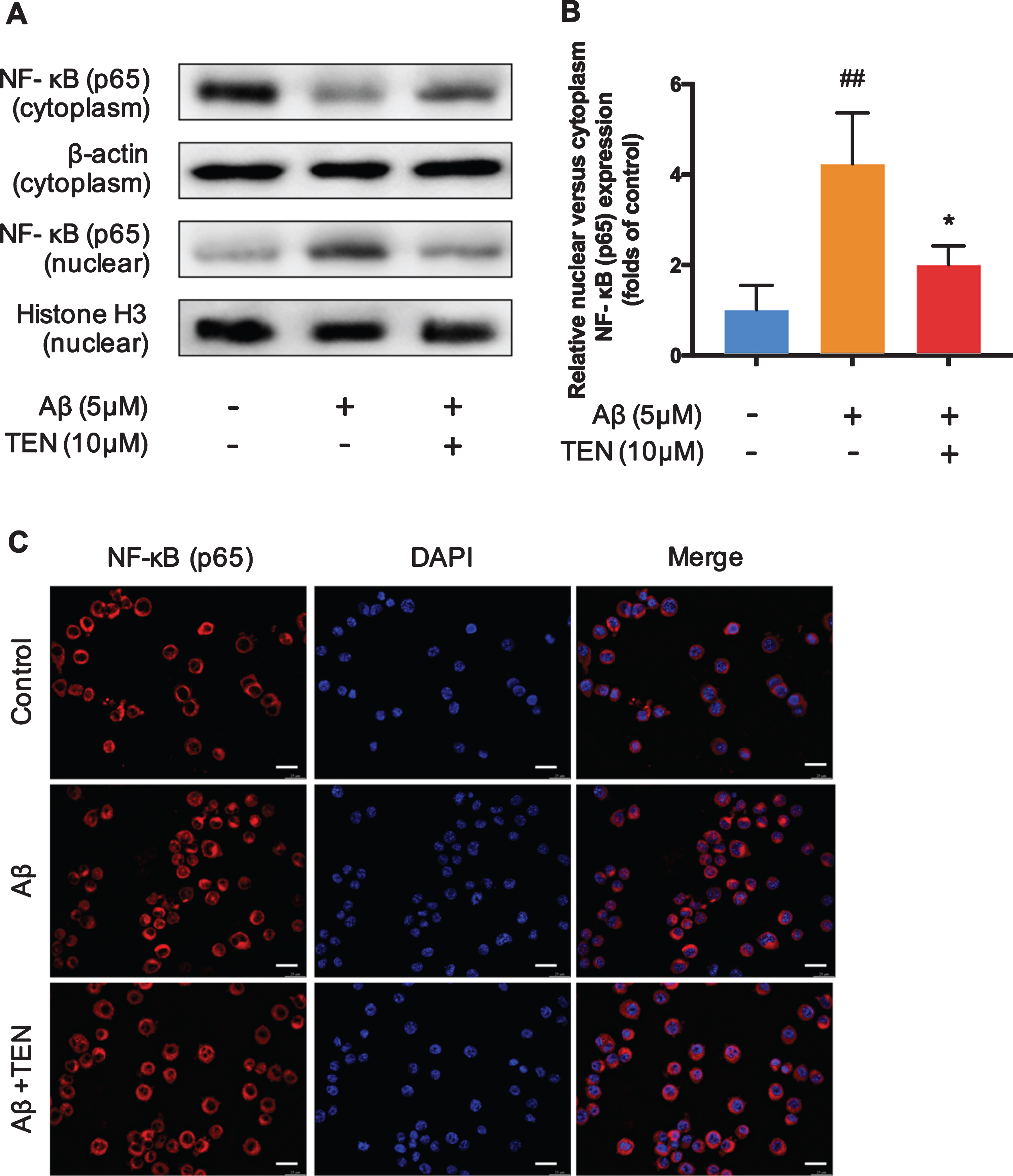
NF-κB changes in the nucleus and cytoplasm. A, B) Western blots and analysis for NF-κB p65 subunit in nuclear and cytosolic fractions after treatment with 5μM Aβ42 oligomers alone or with 10μM TEN for 1 h. C) The representative immunofluorescence images showing translocation of NF-κB (p65, red) to nucleus (blue) after treatment with 5μM Aβ42 oligomers alone or with 10μM TEN for 1 h. Scale bar: 25μm. # # p < 0.01 versus control, * p < 0.05, n = 3 per group.
DISCUSSION
This study indicated that TEN suppresses microglia-mediated neuroinflammation and oxidative stress induced by Aβ42 oligomers. To the best of our knowledge, this is the first study to show that TEN can decrease the level of NO and inhibit the production of the pro-inflammatory cytokines, as well as free radical production, via regulating the NF-κB signaling pathway. Since these effects resulted in the attenuation of microglial cytotoxicity caused by Aβ42 oligomers, TEN could be a potential new therapeutic for the treatment of AD.
The NF-κB protein family consists of various subunits, i.e., p50, p52, p65 (RelA), c-Rel, and RelB. In its resting state, NF-κB dimers are associated with inhibitory IκB proteins or their precursor proteins in the cytoplasm. When the core component of the NF-κB pathway, the IKK complex, is activated, IκB proteins are phosphorylated, ubiquitinated, and degraded. The released NF-κB dimers are then activated and translocate to the nucleus, where they bind to specific DNA sequences and promote the transcription of target genes [16]. Over the last two decades, the role of NF-κB in the nervous system has been characterized in relation to differentiation, apoptosis, inflammation, oxidative stress, insulin resistance, synaptic plasticity, and other processes [17]. In neuronal and microglial cellular models based on the use of Aβ peptides, activation of NF-κB suggests that the NF-κB pathway is related to Aβ neurotoxicity [18]. In turn, NF-κB activation promotes the transcription of amyloid-β protein precursor (AβPP), β-site amyloid precursor protein cleaving enzyme 1 (BACE1), and some of the γ-secretase members, resulting in elevated Aβ levels in vitro. Postmortem studies have shown that NF-κB can also degenerate neurons in the brain of AD patients, particularly in regions that are preferentially affected by the disease. The present study showed that the exposure of BV2 cells to Aβ42 oligomers promoted the phosphorylation of IKKβ, IκB, and NF-κB, as well as caused nuclear translocation of NF-κB (p65). TEN at a concentration of 10μM inhibited the levels of p-IKKβ, p-IκB, and p-NF-κB, and decreased NF-κB translocation to the nucleus. We found that prostratin (10μg/ml), an activator of the NF-κB pathway [19], significantly reduced the inhibitory effect of TEN on this pathway. This further validated that TEN inhibited neuroinflammation by suppressing NF-κB activation in BV2 cells exposed to Aβ42 oligomers.
It is known that levels of TNF-α, IL-1β, and IL-6 are increased in the AD brain [20]. Previous studies have shown that these cytokines may exacerbate the neuropathological alterations of AD, such as the formation and accumulation of Aβ and tau [7, 21]. Analysis of genetic polymorphisms showed that the majority of genetic loci that affect AD are located in the promoter regions of cytokine genes, which further highlights the importance of cytokines in AD pathogenesis [22]. Prior studies have demonstrated that prophylactic and interventional anti-inflammatory strategies reduce the neuropathology and improve cognitive functions in AD animal models. Intriguingly, phase I and phase II clinical trials also showed that inhibitors of inflammation may improve daily activities and cognitive functions in AD patients [23, 24]. The NF-κB pathway is activated by the interaction between a ligand and tumor necrosis factor receptor 1 (TNFR 1) and plays an important role in the activation of the inflammatory process by producing cytokines [25]. Moreover, NF-κB translocation to the nucleus, IKK and p65 phosphorylation, and reduced levels of IκB can be triggered by a broad range of stimuli, such as IL-1β, TNF-α, and LPS through IL-1R, TNFR, and toll-like receptors [26, 27]. Therefore, the inhibition of NF-κB suppresses the induction of cytokines and chemokines. In our study, BV2 microglia stimulation by Aβ42 increased the expression of TNF-α, IL-1β, and IL-6, while TEN partially prevented these effects. Therefore, TEN is capable to break the vicious circle of inflammation between the NF-κB pathway and cytokines.
NO, primarily produced by iNOS, is an important regulatory molecule involved a variety of pathological processes, including inflammation, ischemia, and neurodegeneration [28]. The pathological effects of NO are due to protein nitrotyrosination, which is critical for AD onset. Kummer et al. found that Aβ at tyrosine-10 is regulated by iNOS in APP/PS1 mice, and that senile plaque formation is attenuated by iNOS gene knockout [29]. Stimulated glial cells produce high levels of NO and free radicals, thus promoting oxidative stress and neuronal toxicity in both normal and AD brains [30]. COX-2, constitutively expressed in neurons, is believed to affect Aβ aggregation and deposition, as well as tau phosphorylation [31, 32]. Wang et al. showed that COX-2 dysregulation may cause AD by inducing neuroinflammation, oxidative stress, synaptic plasticity, neurotoxicity, autophagy, and apoptosis [33]. Furthermore, other studies have reported high levels of COX-2 in early-stage AD brains [34, 35]. NF-κB is a primary regulator of various target genes, including iNOS and COX-2 [36]. Consistently, blocking the transcriptional activity of NF-κB in microglial nuclei suppresses the expression of iNOS and COX-2. Notably, in the present study, we found that TEN suppressed Aβ-induced iNOS and COX-2 upregulation at both, mRNA and protein, levels.
As a classic traditional Chinese medicine, Polygala tenuifolia Willd is one of the 10 herbal medicines that have been used for AD treatment worldwide [38]. After druglikeness filtering, 1016 compounds were further analyzed and TEN was found to be one of the typical effective ingredients in Polygala tenuifolia Willd [37]. Although another active ingredient, polygalasaponin, was effective in treating cognitive disorders, it showed high toxicity. In particular, previous studies by Liu et al. have shown that polygalasaponins could cause nosebleeds, gastrointestinal disorders, and even death in laboratory animals [37]. In contrast, TEN, one of the secondary saponins that can be purified and isolated after hydrolysis of polygalasaponins, showed significantly less toxicity [38, 39]. But so far, much lesser studies have been conducted on TEN than other active ingredients, e.g., ginsenoside, curcumin, and huperzine, in AD models. Our results are consistent with the few available studies, which demonstrated that TEN suppresses Aβ42-induced neuroinflammation in vitro [11, 40]. This study provides the first explanation for the anti-inflammatory effects of TEN in AD. Specifically, TEN inhibited Aβ42 oligomer-induced neuroinflammation and oxidative stress by regulating the NF-κB pathway.
In conclusion, the present study showed that TEN may effectively suppress Aβ42-induced production of pro-inflammatory cytokines and free radicals by inhibiting NF-κB signaling. Since NF-κB-mediated neuroinflammation is a key player in AD pathogenesis, TEN may be useful in establishing novel therapeutic strategies for AD.
Footnotes
ACKNOWLEDGMENTS
We gratefully acknowledge Longfei Jia, Heng Zhang, and Bingqiu Li for their contribution to this study.
This study was supported by the Key Project of the National Natural Science Foundation of China (81530036); the National Key Scientific Instrument and Equipment Development Project (31627803); Mission Program of Beijing Municipal Administration of Hospitals (SML20150801); Beijing Scholars Program; Beijing Brain Initiative from Beijing Municipal Science & Technology Commission (Z161100000216137); Project for Outstanding Doctor with Combined Ability of Western and Chinese Medicine; and Beijing Municipal Commission of Health and Family Planning (PXM2019_026283_000003).