
Abstract
Many Alzheimer’s disease (AD) patients suffer from persistent neuropathic pain (NP), which is mediated, at least partially, but microglia. Nevertheless, the exact underlying mechanism is unknown. Moreover, a clinically translatable approach through modulating microglia for treating AD-associated NP is not available. Here, in a doxycycline-induced mouse model (rTg4510) for AD, we showed development of NP. We found that the total number of microglia in the CA3 region was not increased, but polarized to pro-inflammatory M1-like phenotype, with concomitant increases in production and secretion of pro-inflammatory cytokines. To examine whether this microglia polarization plays an essential role in the AD-associated NP, we generated an adeno-associated virus (AAV) serotype PHP.B (capable of crossing the blood-brain barrier) carrying shRNA for DNA methyltransferase 1 (DNMT1) under a microglia-specific TMEM119 promoter (AAV-pTMEM119-shDNMT1), which specifically targeted microglia and induced a M2-like polarization in vitro and in vivo in doxycycline-treated rTg4510 mice. Intravenous infusion of AAV-pTMEM119-shDNMT1 induced M2-polarization of microglia and attenuated both AD-associated behavior impairment but also NP in the doxycycline-treated rTg4510 mice. Thus, our data suggest that AD-associated NP may be treated through M2-polarization of microglia.
INTRODUCTION
The prevalence of neurodegenerative diseases like Alzheimer’s disease (AD) and other dementias continuously increases with the elongation of the lifespan of humans, and these disease are regarded as aging-associated disorders [1]. AD is the most common form of dementia, in which patients suffer from cognitive and behavioral impairments, and nearly half experience chronic pain [2]. Although the correlation between AD and chronic pain has been faithfully detected [3], a causal or mechanistic link is not established.
Microglia are the primary innate immune cells in the central nervous system (CNS), constituting the immune protective barrier against various insults to CNS [4]. It is now known that microglia play critical roles in inflammatory response and tissue recover from traumatic brain injury [5], in development of neurodegenerative diseases [6–8], and in chronic pains [9]. It is traditionally thought that macrophages/microglia are mainly phagocytotic, while they are later found to have much more functions [10–13]. Compared to the classical phagocytotic “M1” macrophages/microglia, the other ones are called alternatively polarized macrophages or “M2” macrophages/microglia, which function as a modulator for cellular and humoral immunity and as a promotor and mediator of tissue repair and remodeling [10–13]. M2 macrophages/microglia highly express specific markers like CD206, CD163, arginase, and CD301, and cytokines like IL-4, IL-10, and IL-13. On the other hand, M1 macrophages/microglia highly express specific markers like CD86, nitric oxide synthase (iNOS), and reactive oxygen species (ROS), and cytokines like tumor necrosis factor alpha (TNFα), IL-1β, and IL-6 [10–13]. Microglial activation could be either pro-inflammatory or anti-inflammatory activation, which promotes neuropsychiatric diseases through pro-inflammatory cytokine and inhibits them, respectively [14]. Very recently, microglia, as a type of tissue-resident macrophages in the CNS, were shown to affect various diseases, including ischemic hemorrhage [15], traumatic brain injury [5], and neuropathic pain [16], through their polarization. Of note, Chitinase1 has been shown to exert protective effects against AD by polarizing microglia to an M2 phenotype [17]. However, whether microglia polarization can influence AD-associated NP is unknown and thus addressed in the current study.
MATERIALS AND METHODS
Animals
All experiments were performed in strict accordance with the Care and Use of Laboratory Animal Guideline, issued by Lanzhou University. The rTg4510 transgenic mice [18] were purchased from Jackson Labs (Bar Harbor, ME, USA). Doxycycline 200 ppm in chow was fed to rTg4510 mouse ad libitum to suppress transgene expression in order to induce the AD phenotype. Mice without receiving doxycycline (DOX off) were used as controls for doxycycline-treated (DOX on) rTg4510 mice. The mice were kept in cages with free access to fresh water and a standard diet, and housed under a 12-h light-dark cycle (temperature: 24±2°C; humidity: 46±4%).
Virus generation, production, and injection
Cell lines HMC3 (microglia cell line), KG-1 (macrophage cell line), HCN-2 (neuronal cell line), and 3T3 (fibroblast cell line) were all purchased from American Type Culture Collection (ATCC, Rockville, MD, USA) and were cultured in Dulbecco’s modified Eagle medium (DMEM, Gibco; Life Technologies, Carlsbad, CA, USA) with 10% fetal bovine serum (FBS, Sigma-Aldrich, St Louis, MO, USA), penicillin (100 μg/ml), and streptomycin (250 ng/ml) at 37°C, in a 5% CO2 atmosphere. The sequence for shRNA for DNA Methyltransferase 1 (shDNMT1) is 5’-CCGGCCCGAGTATGCGCCCATATTTCT CGAGAAATATGGGCGCATACTCGGGTTTTTG-3’, which targets sequence CCCGAGTATGCG CCCATATTT on DNMT1). Scramble sequence is 5’-GGTATCTACTAGATGTACT-3’. Microglia-specific TMEM119 promoter was purchased from GeneCopoeia (#34837, GeneCopoeia, Rockville, MD, USA). AAV serotype PHP.B vector (capable of crossing the blood-brain barrier) selected pUCmini-iCAP-PHP.eB plasmid (Addgene #103005, Addgene, Watertown, MA, USA). AAV serotype PHP.B carrying shDNMT1 or scramble under TMEM119 promoter (AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble) also contained a GFP reporter to be co-expressed with the shDNMT1 or scramble. AAVs were generated by transfection of human embryonic kidney 293 cells with all necessary plasmid by Lipofectamine 3000 reagent (Invitrogen, CA, Carlsbad, USA), according to the instructions of the manufacturer. Titration of viral vectors was determined using a dot-blot assay. Prepared virus was stored at –80°C. For tail vein injection, 1012 viral particles were injected in 100 μl volume.
Behavioral and pain assessments
A Morris water-maze test was used to assess spatial reference memory of the mice. Briefly, all mice were assessed at 5 months of age, regardless of viral administration at 2 months of age. Immediate before testing, mice were trained in the testing pool with a 20-s water exposure daily for 10 days. At testing, mice received visible platform training for 18 trials in 3 days, and then hidden platform training for 18 trials in the subsequent 4 days. The mean target quadrant occupancy was analyzed. A motor assessment was done for testing balance and coordination ability. First, all mice received a grip test for holding a bar with its front paws in 3 consecutive days. The grip test measures the strength with which a mouse holds a bar with its front paws against an experimenter. The force at the give-up point of the mice was recorded. A beam crossing test was applied to evaluate the limb coordination, accuracy of limb placement, and motor coordination by handling a mouse to walk across a relative wide or a narrow beam to reach a home cage.
A von Frey filament test was used to measure pain in mice. Briefly, the mice were put in a test box fitted with a wire mesh backing. Von Frey microfilaments were applied through the grid floor to the ventral surface of the hind paw of the injured hind limb of the mice. The filament was pressed till bent and wait for the mice to withdraw the hind leg without moving. During each test, the filament series was presented after the incremental procedure and the 50% response threshold for each mouse was calculated [19].
Conventional PCR, RT-qPCR, and ELISA
RNA extraction and cDNA synthesis were routine performed. For conventional PCR to check GFP expression in different tissue after viral infusion, we used GFP primers (Forward: 5’-GTCCAGGAGCGCACCATCT-3’, reverse: 5’-GTTCTTCTGCTTGTCGGCCA-3’) and GAPDH primers (Forward: 5’-CATCACTGCCACCCAGAAGACTG-3’, reverse: 5’-ATGCCAGTGAGCTTCCCGTTCAG-3’). RT-qPCR primers were all purchased from Qiagen. Values were normalized against GAPDH, which proved to be stable across the samples, and then compared to experimental controls. ELISA was performed using specific kits for iNOS, arginase (ARG), TNFα, IL-1β, IL-6, and IFNγ (R&D System, Los Angeles, CA, USA).
Flow cytometry
Flow cytometry data was analyzed by FlowJo software (Flowjo LLC, Ashland, OR, USA). Flow cytometry-based macrophage sorting was done using PE-cy5-conjugated CD11b, APC-conjugated CD45, and pacific blue-conjugated CD206 antibodies (Becton-Dickinson Biosciences, Shanghai, China).
Statistics
Statistical analysis was performed using GraphPad Prism 7 (GraphPad Software, San Diego, CA, USA). Analysis was performed using one-way ANOVA with a Bonferroni correction, followed by Fisher’s Exact Test upon necessity. All values are depicted as mean±standard deviation from 5 individuals and are considered significant if p < 0.05.
RESULTS
Development of NP in rTg4510 AD-mice
Due to the recent failure of clinical trials with novels medicine addressing amyloid-β peptide aggregates (Aβ), we chose an AD model with increased phosphorylated tau for formation of intraneuronal neurofibrillary tangles, rTg4510 mice [20]. Doxycycline was administrated to induce AD phenotype (Dox on) and mice without doxycycline treatment were used as control (Dox off). All mice were analyzed at 5 months of age. In Morris water maze testing for assessing spatial reference memory, the mice showed no difference in the path length in the visible platform phase between the Dox-on or Dox-off treated groups (Fig. 1A). However, the Dox-on mice used a significantly longer path length in the hidden platform phase (Fig. 1B) and had significantly more target quadrant occupancy (Fig. 1C), compared to Dox-off mice. For motor assessment, the Dox-on mice exhibited no difference in grip power with the Dox-off mice (Fig. 1D), but required significantly longer time to cross a 2.5 cm-wide beam (Fig. 1E), and fell more frequently when they crossed a 2 cm-wide beam (Fig. 1F), compared to Dox-off mice. Thus, Dox-on rTg4510 mice exhibited impaired memory and locomotor capability, the features for AD. Von Frey filament test was then applied to evaluate the mechanical and thermal pain in 4 consecutive days. Dox-on mice displayed a significant decrease in mechanically-induced withdrawal threshold (Fig. 1G) and a significant decrease in thermally-induced withdrawal latency of the paw (Fig. 1H). Together, these data suggest that Dox-on rTg4510 mice successfully develop AD and AD-associated NP.
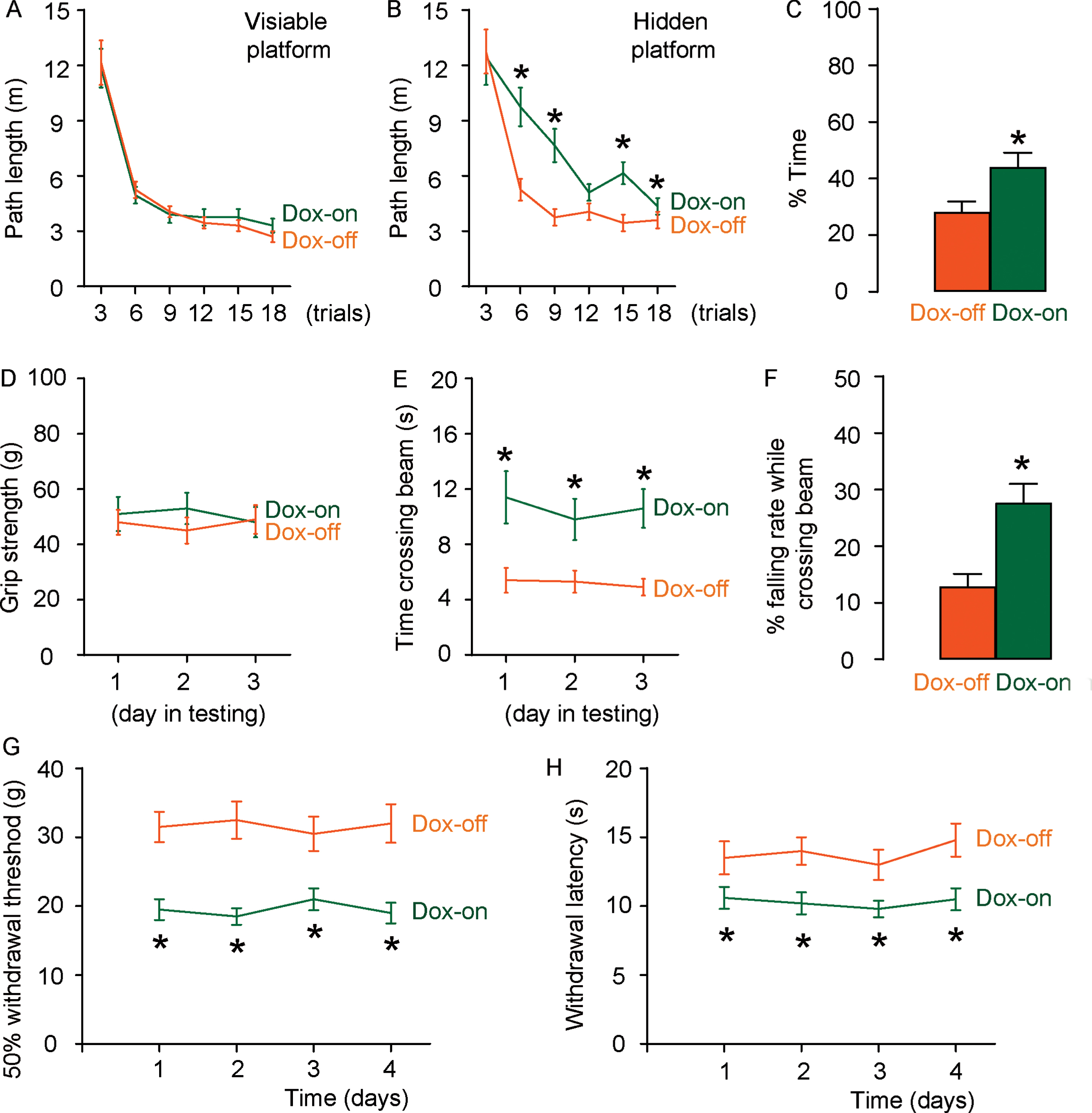
Development of NP in rTg4510 AD-mice. A–C) A Morris Water Maze test for assessment of path length in the visible platform (A), path length in the hidden platform (B), and mean target quadrant occupancy (C) in 5-month-old rTg4510 mice that received doxycycline (Dox-on) or not (Dox-off). (D–F) A motor assessment test of grip strength (D), time crossing a 2.5 cm-wide beam (E), and falling rate during crossing a 2 cm-wide beam (F). *p < 0.05. N = 5.
Increase in pro-inflammatory polarization of microglia in Dox-on rTg4510 mice
Next, we isolated microglia from the CA3 region of the 5-month-old Dox-on and Dox-off rTg4510 mice by flow cytometry based on CD11b+CD45l∘w. The isolated microglia were further analyzed for CD206, a marker for M2 subtype (Fig. 2A). We found that the total number of microglia did not differ between Dox-on and Dox-off rTg4510 mice (Fig. 2B). However, the number and ratio of CD206 + M2 microglia were significantly reduced in Dox-on rTg4510 mice, compared to Dox-off rTg4510 mice (Fig. 2B). Moreover, M1-associated iNOS, TNFα, IL-1β, IL-6, and IFNγ significantly increased, while M2-associated arginase (ARG) significantly reduced in microglia from Dox-on rTg4510 mice, compared to Dox-off rTg4510 mice (Fig. 2C). Together, these data suggest increased pro-inflammatory polarization of microglia in Dox-on rTg4510 mice.
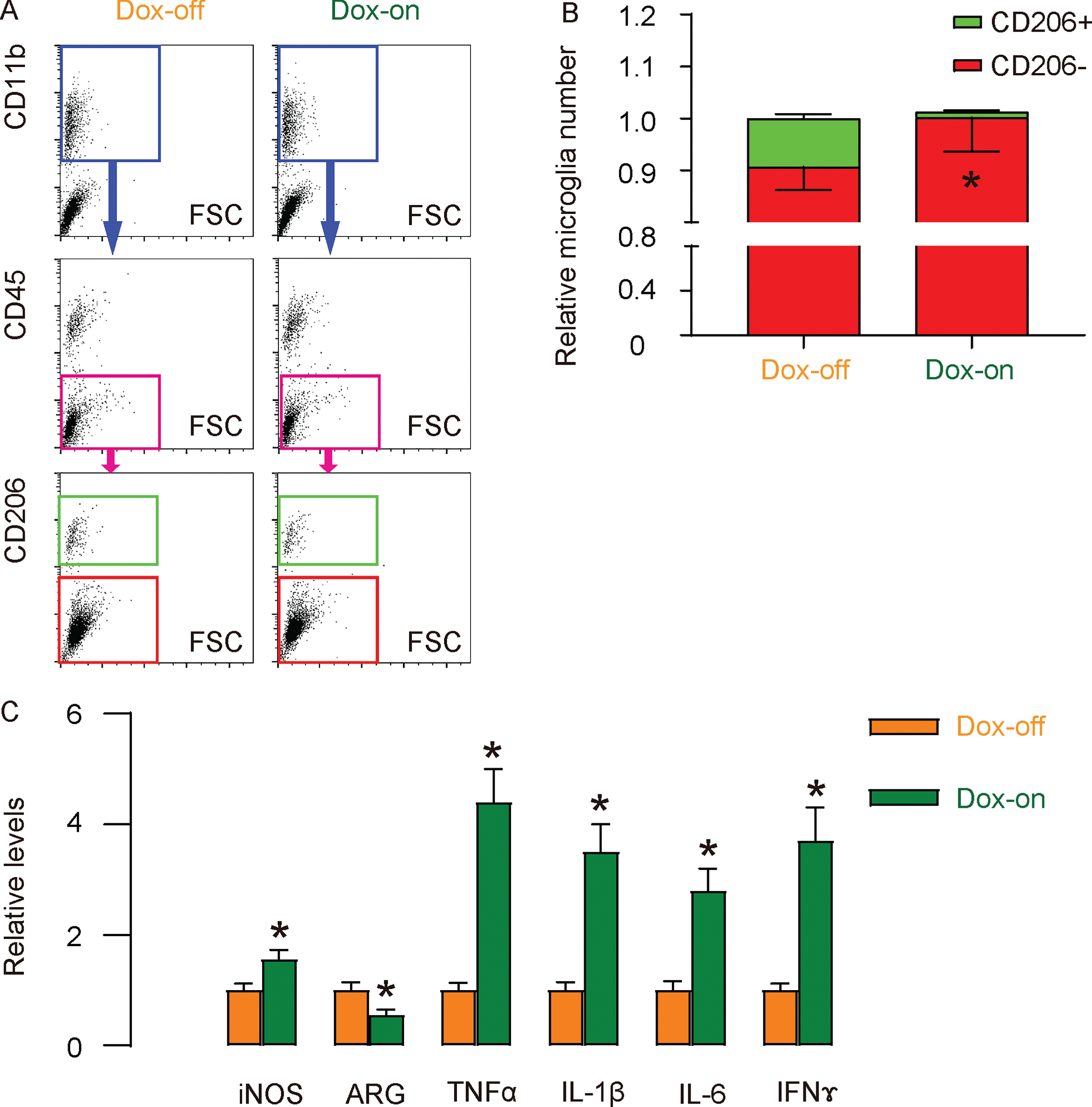
Increase in pro-inflammatory polarization of microglia in Dox-on rTg4510 mice. A) Representative flow charts for isolation of microglia from the CA3 region of the 5-month-old Dox-on and Dox-off rTg4510 mice by flow cytometry based on CD11b+CD45l ∘w. The isolated microglia were further analyzed and sorted by CD206 into CD206-M1 microglia and CD206 + M2 microglia. B) Quantification of CD206 + versus CD206- microglia in CA3 region. C) ELISA for iNOS, ARG, TNFα, IL-1β, IL-6, and IFNγ in isolated microglia from Dox-on and Dox-off rTg4510 mice. *p < 0.05. N = 5.
Generation of AAVs that target microglia and induce M2-like polarization
Then we aimed to examine whether this increased pro-inflammatory polarization of microglia plays a role in the development of AD-associated NP. In order to convert microglia into M2-like in vivo, we generated AAV serotype PHP.B (the serotype capable of crossing blood-brain barrier [21–23]) carrying shDNMT1 or scramble under a microglia-specific TMEM119 promoter [24] (AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble). The viruses also contained a GFP reporter to be co-expressed with the shDNMT1 or scramble. DNMT1 is known to regulate macrophage polarization, while depletion of DNMT1 can induce M2-like polarization [25–27]. First, we examined the specificity for microglia and the potential of inducing M2-polarization by these AAV-pTMEM119-shDNMT1 and AAV-pTMEM119-scramble. AAV-pTMEM119-scramble was found to transduce microglia cell line HMC3, but not macrophage cell line KG-1, or neuronal cell line HCN-2, or fibroblast cell line 3T3 based on viral GFP by flow cytometry (Fig. 3A), confirming the specificity of TMEM119 promoter for microglia. Seven days after tail vein injection of AAV-pTMEM119-scramble, GFP signals were exclusively detected in the mouse brain, but not in the mouse heart, lung, spleen, stomach, intestine, liver, pancreas, kidney, and muscle (Fig. 3B), confirming both the capability of crossing blood-brain barrier by AAV serotype PHP.B, but also the specificity of TMEM119 promoter. Moreover, we detected significantly reduced M1-associated iNOS, TNFα, IL-1β, IL-6, and IFNγ, and significantly increased M2-associated ARG in AAV-pTMEM119-shDNMT1-transduced HMC3 cells, compared to AAV-pTMEM119-scramble-transduced HMC3 cells (Fig. 3C). Together, these data suggest that the generated AAV (serotype PHP.B)-pTMEM119-shDNMT1 is able to cross blood-brain barrier and induce M2-like polarization of microglia specifically.

Generation of AAVs that target microglia and induce M2-like polarization. AAV serotype PHP.B is the serotype capable of crossing blood-brain barrier. Depletion of DNMT1 can induce M2-like polarization. TMEM119 is a microglia-specific promoter. AAV-pTMEM119-shDNMT1 and AAV-pTMEM119-scramble were generated using serotype PHP.B. The viruses also contained a GFP reporter to be co-expressed with the shDNMT1 or scramble. A) The capability of transduction of microglia cell line HMC3, macrophage cell line KG-1, neuronal cell line HCN-2, and fibroblast cell line 3T3 with AAV-pTMEM119-scramble was evaluated by flow cytometry based on viral GFP. B) Viral GFP was detected by PCR in mouse brain, heart, lung, spleen, stomach, intestine, liver, pancreas, kidney, and muscle 7 days after tail vein injection of AAV-pTMEM119-scramble. C) ELISA for iNOS, ARG, TNFα, IL-1β, IL-6, and IFNγ in HMC3 cells treated with either AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble. *p < 0.05. N = 5.
Administration of AAV-pTMEM119-shDNMT1 to Dox-on rTg4510 mice induces M2-polarization of microglia
Next, we did tail vein injection of AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble to Dox-on rTg4510 mice at 2 months of age and analyzed the mice 3 months later. First, we isolated microglia from the CA3 region of these 5-month-old AAV-treated Dox-on rTg4510 mice by flow cytometry based on CD11b+CD45l ∘w. The isolated microglia were further analyzed for CD206, a marker for M2 subtype (Fig. 4A). We found that the total number of microglia did not differ between Dox-on rTg4510 mice treated with either AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble (Fig. 4B). However, the number and ratio of CD206 + M2 microglia were significantly increased in AAV-pTMEM119-shDNMT1-treated Dox-on rTg4510 mice, compared to AAV-pTMEM119-scramble-treated Dox-on rTg4510 mice (Fig. 4B). Moreover, M1-associated iNOS, TNFα, IL-1β, IL-6, and IFNγ significantly decreased, while M2-associated ARG significantly increased in microglia from AAV-pTMEM119-shDNMT1-treated Dox-on rTg4510 mice, compared to AAV-pTMEM119-scramble-treated Dox-on rTg4510 mice (Fig. 4C). Together, these data suggest that administration of AAV-pTMEM119-shDNMT1 to Dox-on rTg4510 mice induces M2-polarization of microglia.

Administration of AAV-pTMEM119-shDNMT1 to Dox-on rTg4510 mice induces M2-polarization of microglia. A) Representative flow charts for isolation of microglia from the CA3 region of the 5-month-old Dox-on rTg4510 mice that had received tail vein injection of either AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble at 2 months of age by flow cytometry based on CD11b+CD45l ∘w. The isolated microglia were further analyzed and sorted by CD206 into CD206- M1 microglia and CD206 + M2 microglia. B) Quantification of CD206 + versus CD206-microglia in CA3 region. () ELISA for iNOS, ARG, TNFα, IL-1β, IL-6, and IFNγ in isolated microglia from the 5-month-old Dox-on rTg4510 mice that had received tail vein injection of either AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble at 2 months of age. *p < 0.05. N = 5.
Administration of AAV-pTMEM119-shDNMT1 to Dox-on rTg4510 mice attenuates NP
Finally, we examined the effects of AAV-pTMEM119-shDNMT1-induced M2 polarization of microglia on AD and AD-associated NP in Dox-on rTg4510 mice. In Morris water-maze testing for assessing spatial reference memory, the mice showed no difference in the path length in the visible platform phase between the AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble treated Dox-on mice (Fig. 5A). However, the AAV-pTMEM119-shDNMT1-treated Dox-on mice used significantly shorter path length in the hidden platform phase (Fig. 5B) and had significantly less target quadrant occupancy (Fig. 5C), compared to AAV-pTMEM119-scramble-treated Dox-on mice. For motor assessment, the AAV-pTMEM119-shDNMT1-treated Dox-on mice exhibited no difference in grip power with the AAV-pTMEM119-scramble-treated Dox-on mice (Fig. 5D), but required significantly shorter time to cross a 2.5 cm-wide beam (Fig. 5E), and fell much less frequently when they crossed a 2 cm-wide beam mean (Fig. 5F), compared to AAV-pTMEM119-scramble-treated Dox-on mice. Thus, AAV-pTMEM119-shDNMT1-treated Dox-on rTg4510 mice established attenuated impairment in memory and locomotor capability. Von Frey filament test was then applied to evaluate the mechanical and thermal pain in 4 consecutive days. AAV-pTMEM119-shDNMT1-treated Dox-on mice displayed a significant increase in mechanically-induced withdrawal threshold (Fig. 5G) and a significant increase in thermally-induced withdrawal latency of the paw (Fig. 5H). Together, these data suggest that administration of AAV-pTMEM119-shDNMT1 to Dox-on rTg4510 mice attenuates AD and AD-associated NP.

Administration of AAV-pTMEM119-shDNMT1 to Dox-on rTg4510 mice attenuates NP. A–C) A Morris Water Maze test for assessment of path length in the visible platform (A), path length in the hidden platform (B), and mean target quadrant occupancy (C) in 5-month-old rTg4510 Dox-on mice that had received tail vein injection of either AAV-pTMEM119-shDNMT1 or AAV-pTMEM119-scramble at 2 months of age. D–F) A motor assessment test of grip strength (D), time crossing a 2.5 cm-wide beam (E), and falling rate during crossing a 2 cm-wide beam (F). *p < 0.05. N = 5.
DISCUSSION
The rTg4510 mouse is a well-defined model for increased NFLs by pTau [20, 28], and we chose it since recent declined evidence on Aβ as a primary pathological lesion in AD and target for therapeutic medicine [29–31]. The determined analyzing time point was based on published data, showing that these mice started to develop cognitive and behavioral impairment as early as about 3 months of age, and obtained severe phenotype at 5 months of age [20].
Epigenetics and biomedical technologies based on epigenetic mechanisms have recently become an important part of the field of life science and technology in the post-genomic era [32]. In-depth research on epigenetic changes such as DNA methylation modification, post-translational modification of histones, and non-coding RNA in the processes of tumors, cardiovascular and cerebrovascular diseases, diabetes, and middle-aged and elderly neurodegenerative diseases help us to understand related diseases. A new branch of modern molecular biology characterizes the gene expression patterns required for “environment-gene interaction” [33]. Various types of epigenetic modifications such as methylation modifications at specific sites in DNA, post-translational modifications of histones in nucleosomes, non-coding RNA regulation, histone variants, and catalyzed chromatin remodeling have been identified [32]. Recent research suggests that microglial/macrophage polarization is a complex epigenetic controlled process. The 5 ‘position of cytosine at certain sites in eukaryotic DNA molecules is a common methylation modification (5-Methylcytosine, 5-meC) site [34–38]. Cytosine methylation modifications appearing at corresponding sites are one of the important reasons for epigenetic modification to promote gene silencing and chromatin to become “heterochromatin” [34–38]. In mammalian cells, methylation modification of DNA is common at the C bases that are scattered in the “CpG” dinucleotide site upstream of the promoter. The 5th position of the “C” base in the “CpG” at these sites can accept the methyl group from S-adenosylmethionine under the catalysis of DNA methyltransferase [34–38]. Unmethylated CpG is recognized by CXXC domain-containing proteins, while methylated CpGs are recognized by methyl binding domain proteins [34–38]. DNA methylation modulation regulates polarization in macrophages, for example, using 5-aza-2’-deoxycytidine to inhibit DNA methylation, or removing DNMT1 through the genome can promote M2 activation of macrophages and inhibit the pro-inflammatory effects of macrophages [39]. Here, we depleted DNMT1 in microglia and successfully reversed the M1 pro-inflammatory phenotype to an anti-inflammatory M2 phenotype.
In order to direct shDNMT1 specifically and exclusively to microglia, we applied several strategies. First, we used a unique AAV serotype, PHP.B, which has been shown to cross the blood-brain barrier to enter the brain. This method was first published by Deverman in 2016 and has been proven by multiple laboratories [40]. Second, we used a microglia-specific TMEM119 promoter and we showed that this promoter did not drive transgene expression in macrophages, neuronal cells, and fibroblasts, the major cell types in the brain besides microglia. These approaches guaranteed the unique targeting of microglia with this gene therapeutic strategy.
To summarize, here using a novel gene therapeutic strategy, we showed a clinic-translational method to reduce AD-associated NP through in vivo modulation of microglia polarization.
DISCLOSURE STATEMENT
Authors’ disclosures available online (https://www.j-alz.com/manuscript-disclosures/20-0099r1).