
Abstract
Background:
Women are reported more likely to develop depression and dementia. However, the involved mechanism is poorly understood.
Objective:
Here, we clarified the role of estrogen receptor α (ERα) in depression and cognitive deficit in young female rats.
Methods:
After being exposed to 7-weeks’ chronic unpredicted mild stress (CUMS), the depression resilient rats (Res rats) and depressed rats (Dep rats) were selected according to their records in sucrose preference test, forced swimming test, and open field test. Their cognition abilities were tested by Morris water maze. Proteomic assay, immunoprecipitation, western blotting, immunohistochemical, and Nissl staining were also used to understand the involved mechanism.
Results:
Compared with control rats and Res rats, Dep rats showed cognitive deficits and hippocampal impairments revealed by proteomic data, neuron losses, increased cleaved caspase-3, β-catenin phosphorylation, and glycogen synthase kinase3β (GSK3β) activation. As ERα, but not ERβ, was found declined in hippocampi of Dep rats, 4,4k,4a-(4-propyl-[1H]-pyrazole-1,3,5-triyl) trisphenol (PPT, an ERα agonist, 1 mg/kg/day), was used to treat Dep rats (Dep + PPT). Twenty days later, the depressive behaviors, cognition deficits, and hippocampal neuron loss were rescued in Dep + PPT rats. Furthermore, Res and Dep + PPT rats had higher levels of β-catenin combined with ERα and lower levels of β-catenin combined with GSK3β than Dep rats in hippocampi.
Conclusion:
These results demonstrated hippocampal ERα is an important pro-resilient factor in CUMS-induced depressive behaviors and cognitive deficits. It was also given that the neuroprotection afforded by hippocampal ERα/Wnt interactions have significant implications for cognition and emotion in young females.
INTRODUCTION
Depression is prevalent neuropsychiatric disorders and epidemiological studies have revealed that depression will be the foremost contributor to the worldwide burden of disease by 2020 [1, 2]. Women are 2.5 times more likely to experience depression than men, and depression mainly occurs in the stages of hormonal changing, such as puberty, pregnancy, postpartum, and perimenopause [3–8]. Depression complicated with dementia in women is common and more women have dementia than men [9]. Early life depression acts as a risk factor for later life dementia and exacerbates cognitive impairments [10–16]. Therefore, to study the actions of the psychopathological progress in stress with female is very meaningful and important.
Hippocampus is a brain region sensitive to stresses [17]. Most autopsy results have shown that a dramatic drop of the hippocampal volume in the individuals with depression compared to the normal volunteers [17–19]. And it was mentioned that the girls or female with small hippocampal volumes were more likely to suffer from depression [17–22]. Estrogen signaling has been implicated in the pathophysiology of depression, while the previous studies demonstrated estrogen replacement therapy (ERT) could reduce depression and mood-related symptomology in menopausal patients [23, 24]. ERT-mediated antidepressant affects mainly through ERα and ERβ in the brain [7, 27]. However, the specific roles of estrogen receptor remain controversial. Activating ERα could significantly improve cognitive impairment and depressive-like behavior in ovariectomized (OVX) rats [28–30]. The postpartum rats treated with an ERα agonist, 4,4k,4a-(4-propyl-[1H]-pyrazole-1,3,5-triyl) trisphenol (PPT), could decline depressive-like behavior also, suggesting a predominant role for ERα in depression [28–31]. In contrast, some other studies have found that ERT improved anxiety and depressive-like behavior in OVX rats via ERβ, but not ERα [30–34]. These disparities require further investigations.
Wnt signaling is a major player in both nervous system development and adult synaptic plasticity and is critical for the functioning of hippocampus [35]. Glycogen synthase kinase3β (GSK3β) and β-catenin are two important molecules in Wnt signaling. Previous studies supported that ERα, GSK3β, and β-catenin form a functional complex [33–38]. Abundant research mentioned the cord action of GSK3β and β-catenin in mood and memory, like GSK3β is a new drug target in depression and β-catenin regulates the formation of memory [32–37]. So, to study the interactions between ERα and Wnt signaling may be important to understand the mechanism underlying hippocampal impairments of females under stress.
Chronic unpredicted mild stress (CUMS) was widely used in reproducing depressive-like animal model [39–42]. In this study, all 125 rats had normal mental status evaluated by depression-related behavioral tests, e.g., sucrose preference test (SPT), forced swimming test (FST), and open field test (OFT). Then 25 rats were randomly chosen as control (Con), and 100 rats underwent CUMS. Seven weeks later, 44 resilient rats (Res) and 32 depression rats (Dep) were determined according to the records from SPT, FST, and OFT. We also detected the spatial cognitive impairments in Dep rats by Morris water maze (MWM). The hippocampal impairments of Dep rats were furtherly revealed by the differentially expressed proteins from the proteomic analysis, the obvious neuron losses in CA1 and CA3 regions, increased cleaved caspase-3, activated GSK3β, and increased phosphorylation of β-catenin. ERα, but not ERβ, were specifically decreased in the hippocampi of Dep rats compared with Con rats and Res rats. Then, we used PPT to treat Dep rats (Dep + PPT, 1 mg/kg/day, n = 10) for 20 days. We found PPT could improve depressive behavior, cognitive deficits, and neuron loss in Dep + PPT rats. And we also found Res rats had a higher level of β-catenin combined with ERα than the vehicle treated Dep (Dep + Veh, n = 9) rats in hippocampi. Dep + Veh rats had the lowest level of β-catenin combined with ERα and the highest level of β-catenin combined with GSK3β. In contrast, Dep + PPT rats had the highest level of β-catenin combined with ERα and the lowest level of β-catenin combined with GSK3β, these results indicating the increased binding of β-catenin with ERα in the hippocampus is beneficial. Thus, the study prompted that ERα is an important pro-resilient factor in CUMS-induced depression and dementia in young female rats.
MATERIALS AND METHODS
Antibodies and chemicals
The primary antibodies used in this study are listed in Table 1. PPT was from Tocris Bioscience (Ellisville, MO, USA), Bicinchoninic acid (BCA) protein detection kit, and phosphocellulose units were from Pierce Chemical Company (Rockford, IL, USA). Anti-rabbit or anti-mouse IgG conjugated to IRDye@ (800CW) (1:10,000) was from Lincoln (USA). Toluidine blue and dimethyl sulfoxide (DMSO) were from Sigma (St. Louis, MO, USA). Diaminobenzidine (DAB) tetrachloride system was from Beijing Zhongshan Jinqiao Biotechnology Co., Ltd. (Beijing, China).
Antibodies employed in the study
WB, western blotting; IHC, immunohistochemistry; IP, immunoprecipitation; p, phosphorylated; M, mouse; R, rabbit.
Animals and treatments
Seven-week-old female SD rats (n = 125) were supplied by Experimental Animal Central of Tongji Medical College, Huazhong University of Science and Technology. All experimental procedures in this research have been approved by the Animal Care and Use Committee of Huazhong University of Science and Technology.
CUMS was used to reproduce depression model in rats as previously [39, 40]. 125 rats with normal mental status evaluated by SPT, FST, and OFT were used in this study. 25 rats were randomly chosen as control (Con), and 100 rats accepted CUMS. Seven weeks later, 44 resilient rats (Res) and 32 depressed rats (Dep) were selected according to the records from SPT, FST, and OFT. Then, 10 Dep rats underwent 20 days of PPT treatment (Dep + PPT, subcutaneous injection, 1 mg/kg/day) and 9 Dep rats underwent 20-days of vehicle treatment (Dep + Veh, daily subcutaneous injection with an equal volume of solvent, 50% DMSO/50% Dulbecco’s PBS). We also detected the spatial cognitive abilities of rats by MWM. All behavioral tests were performed during the light cycle in a dedicated sound-proof behavioral facility by experimenters blind to treatment information.
Chronic unpredicted mild stress (CUMS)
CUMS paradigm was performed as shown in Table 2. Briefly, the rats were exposed to a variety of stressors, by applying four kinds of stressors daily. The stressors included water or food deprivation (24 h), water and food deprivation (24 h), empty water bottles (2 h), cold room (4°C, 2 h), hot room (45°C, 15 min), cage tilt (16 h), continuous lighting (12 h), soiled cage (200 ml of water spilled onto the bedding, 12 h), grouped housing in one cage (4–5 per cage, 12 h), strobe lighting (200 flashes/min, 4 h), and intermittent white noise (85 dB, 6 h).
Procedure of chronic unpredicted mild stress (CUMS) in this research
ND, next day.
Sucrose preference test (SPT)
In SPT, there was a 7-day training phase, 24 h food and water deprivation, and 1 h testing phase as previously described [40, 42]. In the training phase, rats were trained to consume 1.5% water sucrose solution for 1 h (9:30 am–10:30 am) every day. In the testing phase, after 24 h of food and water deprivation, rats were allowed free access to two pre-weighed bottles of solution, e.g., 100 mL of 1.5% sucrose solution and 100 mL of top water, for 1 h. To prevent a possible behavioral interference caused by location, the left/right location of the bottles was switched. The score is the result of sucrose consumption volume/(water consumption volume + sucrose consumption volume)×100%. The second SPT was begun at the 7th week of CUMS (Fig. 1a).
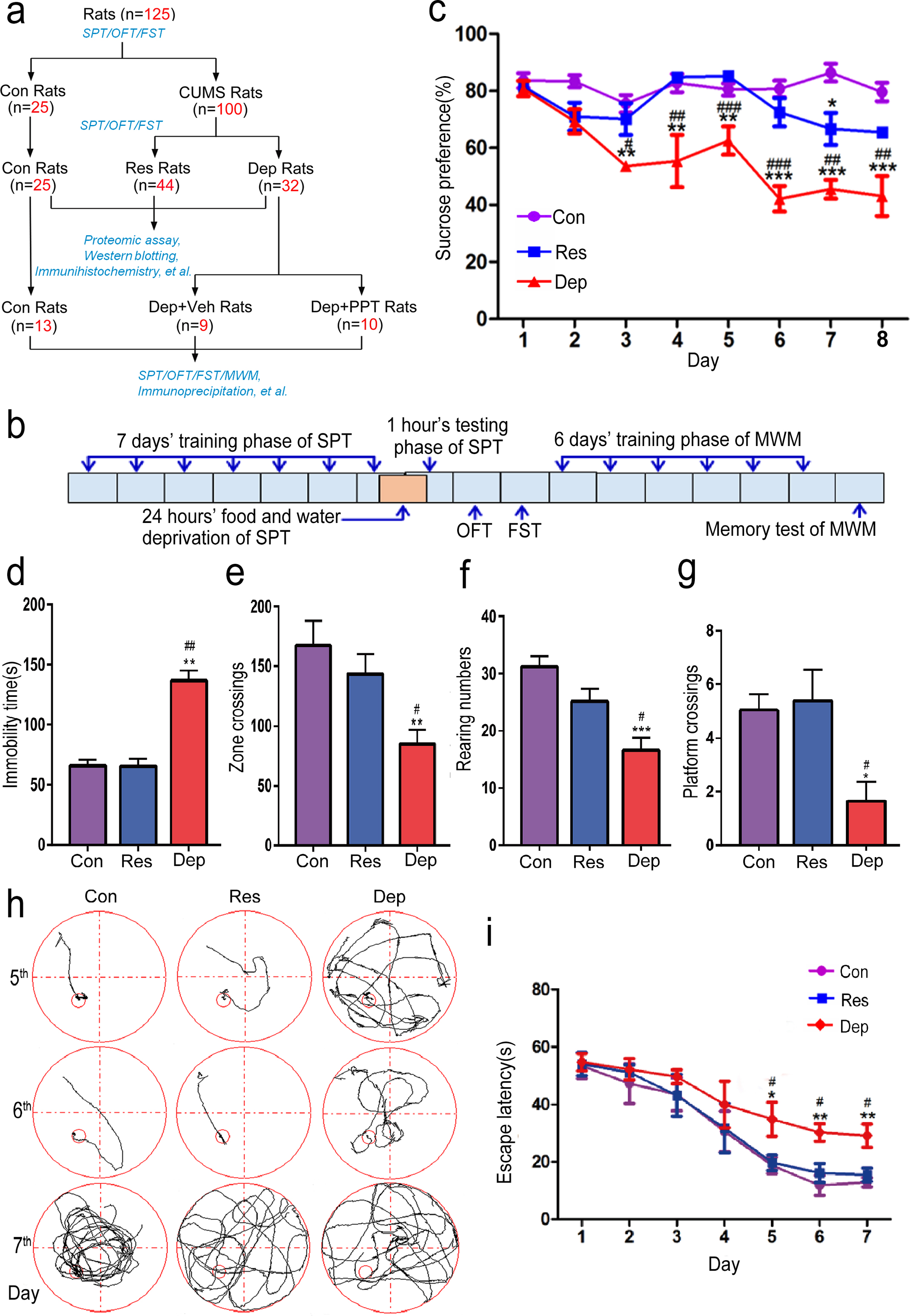
Depressed female rats also showed spatial memory deficit. A diagram of this study (a). Young female rats with normal mental status evaluated by sucrose preference test (SPT), forced swimming test (FST), and open field test (OFT). 25 rats were randomly chosen as control (Con), and 100 rats were exposed to 7 weeks of chronic unpredicted mild stress (CUMS). Depression resilient rats (Res rats) and depressed rats (Dep rats) were selected according to their records in SPT, FST, and OPT (a). Then, we used 4,4k,4a-(4-propyl-[1H]-pyrazole-1,3,5-triyl) trisphenol (PPT) to treat Dep rats (Dep + PPT, 1 mg/kg/day, n = 10) for 20 days (a). Behavior tests including SPT, FST, OPT, and Morris water maze (MWM), immunoprecipitation and western blotting analysis, and morphological studies were performed (a). The timing and order for the behavioral tests were shown in (b). The percentages of sucrose consumption in SPT (c), the immobility times in FST (d), the numbers of zone crossing (e), and the rearing numbers (f) in OFT of control rats (Con rats, n = 25), Res rats (n = 44), and Dep rats (n = 32) were recorded. In the training phase of MWM, the times crossing the hidden platform (g) from 2nd quadrant in the memory test of rats, the swimming pathways (h) and the latencies to find the platform from 2nd quadrant (i) were recorded. Data were presented as means±S.E.M. *p < 0.05, **p < 0.01, ***p < 0.001 versus Con rats. #p < 0.05, # #p < 0.01, # # #p < 0.001 versus Res rats.
Open field test (OFT)
In this test, a bare square box with 100 cm (length)×100 cm (width)×40 cm (height) was used as previously described [40]. Each rat was placed in the center of a black floor with 25 equal squares (20 cm×20 cm per square) including 16 peripheral squares (along with the wall of the box) and 9 central squares. The behaviors of the rat were recorded for 5 min by an overhanging camera linking to a computer. The box was cleaned with 75% alcohol before the next test.
Force swimming test (FST)
In FST, the rat was tested in transparent glass cylinders (height = 50 cm, diameter = 20 cm) containing water at 23–25°C and depth of 14 cm as previously [40]. Briefly, the rat was habituated for the first 1 min and its behaviors were monitored in the next 5 min. A 6 min test session was videotaped, and immobility scores were recorded.
Morris water maze (MWM)
As previously [28], the rats were acclimatized in room and temperature and water was kept at 26±2°C. The water in the pool was made opaque with ink to hide the escape platform. The plexiglass platform was 40 cm high, and 10 cm in diameter and its surface was scarred to help rats climb. The water surface was 18 cm from the rim of the pool and inner wall was consistently carefully wiped to eliminate any local cues. The rim of the pool was 1.0 m from the nearest visual cue of red and blue marks. The behaviors of rats were recorded by an overhanging camera linking to a computer. The submerged platform (2 cm below the surface of the water) was located at a fixed position throughout training. A training session consisted of 3 trials altogether (one trial per quadrant) with a 30 s interval, lasting for 6 days. The first quadrant was included as the first trial every day. On each trial, the rat started from the middle of the quadrant facing the wall of the pool and the trial ended when it climbed the platform. The rats were not allowed to search for the platform more than 60 s, after which they were guided to the platform. In memory testing, the platform was removed and the behaviors of rats were recorded.
Immunoprecipitation and western blotting analysis
The rats were anaesthetized with isoflurane and sacrificed. The hippocampi were quickly dissected out of brain and homogenized at 4°C using a Teflon glass homogenizer in 50.0 mmol/L Tris-HCl (pH 7.4), 150.0 mmol/L NaCl, 10.0 mmol/L NaF, 1.0 mmol/L Na3VO4, 10.0 mmol/L β-mercaptoethanol, 5.0 mmol/L EDTA, 2.0 mmol/L benzamidine, 1.0 mmol/L phenylmethylsulfonyl fluoride, 5.0μg/mL leupeptin, 5.0μg/mL aprotinin, and 2.0μg/mL pepstatin. For immunoprecipitation, the hippocampal extracts (about 500μg total proteins) were immunoprecipitated with anti-ERα or anti-GSK3β at 4°C and shaken overnight, and then protein G agarose (Pierce Chemical Company, Rockford, IL) was added and incubated at 4°C for 2 h. The agarose beads were collected, washed, and resuspended in 60μL of sample buffer containing 50 mmol/L Tris-HCl, pH 7.6, 2% SDS, 10% glycerol, 10 mmol/L dithiothreitol, and 0.2% bromophenol blue and boiled for 5 min and analyzed by western blotting. The western blotting analysis was performed as previously [40, 42]. Briefly, the homogenate was mixed in 2:1 (v/v) ratio with lysis buffer containing 200.0 mmol/L Tris-HCl (pH 7.6), 8% SDS, 40% glycerol and boiled for 10 min in a water bath, stored at –80°C for western blotting analysis. The protein concentrations of the extracts were measured by a BCA kit according to the manufacturer’s instruction. The protein was separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. The membranes were blocked with 2% nonfat milk dissolved in TBS (50.0 mmol/L Tris-HCl, pH 7.6, 150.0 mmol/L NaCl) for 1 h and probed with the primary antibodies for overnight. The blots were incubated with anti-rabbit or anti-mouse IgG conjugated to IRDye™ (800 CW) (1:10,000) for 1 h at room temperature. The intensities of immunoblotting strips were automatically recognized by the Odyssey system (Li-Cor Bioscience, Lincoln, NE). All intensities of strips were normalized by the average intensity of DM1A. Then take the average value of Con group as 1, and calculate the relative intensities of each strip.
Immunohistochemical staining and Nissl staining
The brain slices were prepared and stained as previously [40–42]. The rats were anesthetized with isoflurane and transcardially perfused with 100 mL normal saline and then perfused with 400 mL 4% paraformaldehyde solution. The brains were dissected and post-fixed in perfusate overnight at 4°C, and then cut into coronal slices (30 microns) with a freezing microtome (Leica, Nussloch, Germany; S100, TPI) after being dehydrated twice in 30% sucrose in PBS. The brain slices were collected consecutively in PBS. For DAB based immunohistochemical staining, the free-floating sections were incubated with 3% normal serum in PBS containing 0.1% Triton X-100 and 0.3% H2O2 for 30 min at room temperature to block the nonspecific surfaces. Then the slices were incubated overnight at 4°C with primary antibodies. After washing with PBS, the slices were subsequently incubated with biotin-labeled secondary antibodies for 1 h at 37°C. The immunoreaction was detected using horseradish peroxidase-labelled antibodies and visualized with the DAB tetrachloride system. To test the validities and specificities of the antibodies, PBS and normal serum were used as control to substitute for the primary antibodies. For each primary antibody, 3 to 5 consecutive slices from each brain were used. For Nissl staining studies, the selected slices were washed twice for 5 min in 0.01 mol/L PBS and incubated in 0.1% toluidine blue staining solution for 10 min at room temperature. Then they were rinsed in distilled water, soaked in 95% ethanol for 10 min, and dehydrated in 100% ethanol. After dehydration, the slices were cleared in xylene and covered with neutralized resin. The images were acquired with the same parameters by a microscope (Olympus SV120, Tokyo, Japan) and analyzed with Image-Pro Plus 4.5 system (Media Cybernetics Inc., USA).
Proteomic assay
The rats were anesthetized with isoflurane and sacrificed immediately by decapitation, then the hippocampus was immediately removed from the brain and stored at –80°C after being frozen in liquid nitrogen for 1 min. The proteomic analysis procedure was conducted as previously [40–44]. Briefly, the hippocampus proteins were extracted, digested and labeled by isobaric tags for relative and absolute quantification (iTRAQ)-6plex reagents in accordance with the manufacturer’s protocol. After labeled, the peptides were fractionated by high pH reverse-phase high performance liquid chromatography (HPLC). The resulting fractions were dissolved, loaded onto a reversed-phase pre-column (Acclaim PepMap 100, Thermo Scientific), and then separated using a reversed-phase analytical column (Acclaim PepMap RSLC, Thermo Scientific). The peptides were accepted to NSI source followed by tandem mass spectrometry (MS/MS) in Q Exactive TM Plus (Thermo Scientific) coupled online to the ultra-performance liquid chromatography (UPLC). Intact peptides were detected in the Orbitrap at a resolution of 70,000. Peptides were selected for MS/MS using NCE setting as 28, 32, ion fragments were detected in the Orbitrap with a resolution of 17,500. To identify the proteins, we analyzed the resulting MS/MS data using the Mascot search engine (v.2.3.0) and searched against Uniprot_rat database (32,983 sequences). As reportedly, we defined observed proteins with iTRAQ ratios of >1.2 and <0.83 as differentially expressed. Furthermore, the protein-protein interaction (PPI) network of the differentially expressed proteins were analyzed by The Search Tool for the Retrieval of Interacting Genes (STRING) database (http://string-db.org/), and these data were visualized by Cytoscape Software (https://cytoscape.org/).
Statistical analysis
The data were analyzed using SPSS 12.0 (SPSS Inc., Chicago, IL, USA), and the statistical graphs were produced by GraphPad Prism (GraphPad Software, Inc., La Jolla, CA). Data were expressed as means±SEM. The differences among Con, Res, and Dep group and Con, Dep + Veh, and Dep + PPT group were tested with the one-way analysis of variance procedure followed by Turkey’s multiple comparisons test. The differences between Dep + Veh and Dep + PPT group in the last experiment were tested with Student’s t tests. The level of significance was set at p < 0.05.
RESULTS
Depressed female rats induced by CUMS also showed spatial memory deficit
As shown in Fig. 1a and 1b, after CUMS the Dep rat was defined as the rat whose preference to sucrose in SPT was less than 43%, immobility time in FST was more than 104 s, and rearing number was less than 20 and zone crossing was less than 100 times in OFT. In contrast, the rats having higher preference to sucrose (>60%) in SPT, less immobility time in FST (<80 s), and higher rearing number (>21) and zone crossing times (>139) were defined as Res rats (Fig. 1c, F2,98 = 33.78, p < 0.0001; 1d, F2,98 = 33.78, p < 0.01; 1e, F2,98 = 5.596, p < 0.01; 1f, F2,98 = 9.418, p < 0.01). From 125 rats with normal mental status, 25 rats were randomly chosen as control (Con), and 100 rats underwent CUMS. Seven weeks later, there were 44 resilient rats (Res) and 32 depression rats (Dep) according to their records from SPT, FST, and OFT. We measured the spatial learning and memory abilities of rats by MWM. In the learning testing phase, no difference was shown between Con rats, Dep rats, and Res rats from the 1st to 4th day (Fig. 1i). On the 5th and 6th days of training, Dep rats had obviously increased latencies to find the platform as compared with Con and Res rats (Fig. 1h, i, F2,98 = 27.73, p < 0.01). In the memory testing phase, the times of crossing platform is 1.25±1.02 times in Dep rats, much less than those in Con rats (4.88±1.12 times) and Res rats (5.0±1.3 times) (F2,98 = 4.276, p < 0.05) (Fig. 1g). These results demonstrate that 7 weeks of CUMS induced depressive behaviors and cognition deficits in young female rats.
Hippocampal impairments in Dep rats
Given the key role in depression and dementia, the hippocampal proteomics has been extensively studied by proteomic analysis. In total, 4,690 proteins were identified and 3,645 proteins were quantified. When setting quantification ratio of >1.2 coupled with p < 0.05 as an upregulated threshold and <0.83 coupled with p < 0.05 as a downregulated threshold, 262 differentially expressed proteins were obtained. In the comparison between Dep and Con rats (Dep/Con), there were 77 differentially expressed proteins, 42 upregulated and 35 downregulated proteins (Fig. 2a). There were 19 upregulated and 14 downregulated proteins in the comparison between Res rats and Con rats (Res/Con) (Fig. 2b). In the comparison between Dep rats with Res rats (Dep/Res), there were 20 upregulated and 17 downregulated proteins (Fig. 2c). Ten differentially expressed proteins with high ratio (Fig. 2d) and 9 proteins with low ratio (Fig. 2e) in both Dep/Con and Dep/Res were normal in Res/Con, indicating the proteins with depression related alterations. The changes of 19 proteins were stress related, among which 5 proteins with high ratio and 14 proteins with a low ratio in both Dep/Con and Res/Con (Fig. 2f) were normal in Dep/Res.

Hippocampal impairments in Dep rats. By proteomic analysis of hippocampi, 262 differentially expressed proteins were obtained. In the comparison between Dep and Con rats (Dep/Con), there were 77 differentially expressed proteins, 42 upregulated and 35 downregulated proteins (a). There were 19 upregulated and 14 downregulated proteins in the comparison between Res rats and Con rats (Res/Con) (b). In the comparison between Dep rats with Res rats (Dep/Res), there were 20 upregulated and 17 downregulated proteins (c). 10 differentially expressed proteins with high ratio (d) and 9 proteins with low ratio (e) in both Dep/Con and Dep/Res were normal in Res/Con. The changes of 19 proteins were stress related, among which 5 proteins with high ratio and 14 proteins with a low ratio in both Dep/Con and Res/Con (f) were normal in Dep/Res. The numbers of hippocampal neurons were shown by Nissl staining on brain slices (g) and quantitatively analyzed (h) (n = 3 rats/group, scale bar = 100μm). The levels of hippocampal caspase-3 and cleaved caspase-3 (C-caspase 3) were tested by western blotting (i) and quantitative analysis (j) (n = 5 rats/group, Data were presented as means±S.E.M.). *p < 0.05, versus Con rats. #p < 0.05 versus Res rats.
Hippocampal neuron loss in Dep rats was indicated by increased tenascin-C (TNC) and pleiotrophin from the proteomic assay. TNC was reported to cause post-SAH neuronal apoptosis [47]. It was significantly increased in Dep rats (1.484 in Dep/Con, p < 0.001, and 1.443 in Dep/Res, p < 0.001) (Fig. 2d–f). Pleiotrophin, a cytokine highly upregulated in the brain in different disorders characterized by overt neuroinflammation [48], increased to 1.232 times (p < 0.05) in Dep/Con and 1.297 times (p < 0.05) in Dep/Res. NLR family member X1 (NLRX1), a mitochondrial NOD-like receptor that amplifies the nuclear factor-κ B (NF-κB) and c-Jun N-terminal kinase (JNK) pathways [49], increased to 1.209 times (p < 0.05) in Dep/Res. Overexpression of NLRX1 resulted in the increased reactive oxygen species (ROS) production [49]. As an important negative modulator of Nav1.2 and Nav1.6 channels [50], proline-rich transmembrane protein 2 (PRRT2) increased to 1.242 times (p < 0.001) in Dep/Con and 1.312 times (p < 0.01) in Res/Con. PRRT2 was reported to impair the cell viability and promote apoptosis when overexpressed [51]. Then, we tested the neuron number in the hippocampus by Nissl staining and found that Dep rats showed significantly neuron losses in the CA1 (F2,6 = 9.287, p < 0.05) and CA3 region (F2,6 = 21.65, p < 0.05) (Fig. 2g, h). And the level of cleaved caspase-3 was much higher in the hippocampus of Dep rats by western blotting (Fig. 2i, j, p < 0.05). Additionally, hippocampal synaptic impairments in Dep rats was also suggested. Sushi domain containing-2 (Susd2), a transmembrane protein regulating neurite growth and excitatory synaptic density in hippocampal cultures, was decreased in Dep rats [52], as evidenced by 0.7122 (p < 0.05) in Dep/Con and 0.744 (p < 0.01) in Dep/Res. Whereas, Res rats had a higher level of OTU deubiquitinase 7A (OTUD7A) (0.806 in Dep/Res, p < 0.05), reduced OTUD7A was found to contribute to the dendritic spine and dendrite outgrowth deficits [53].
ERα decreased in Dep rats, but not in Res rats
In the proteomic data, Dep rats had a lower level of leukemia-related protein 16 (LRP16), a coactivator of ERα [54], which was 0.825 (p < 0.05) in Dep/Res. ERα and ERβ are expressed in the pituitary and many brain regions [54–60]. Then we tested levels of hippocampal ERα and ERβ by western blotting. Compared with Con rats, Dep rats had a significantly decreased level of ERα (p < 0.05) while Res rats had no change (Fig. 3a, b). However, levels of ERβ decreased both in Dep rats (p < 0.05) and Res rats (p < 0.05) (Fig. 3a, b). By immunohistochemical analysis, decreased hippocampal ERα was shown in the CA1 (F2,6 = 70.23, p < 0.05) and CA3 (F2,6 = 49.75, p < 0.05) regions of Dep rats (Fig. 3c, d). Additionally, ERα was also decreased in the PFC of Dep rats (data not shown). GSK3β plays an important role in ERα nuclear translocation and 17β-estradiol-dependent activation of ERα [33, 36]. Previously, we reported hippocampal ERα deficit in young OVX rats with GSK3β activation [61]. Here, the phosphorylation level of GSK3β at Ser9 was significantly decreased (indicating its activating) in hippocampus of Dep rats. By immunohistochemical analysis, decreased phosphorylation level of GSK3β at Ser9 was shown in CA1 and CA3 regions of Dep rats (data not shown). Furtherly, we assayed the hippocampal levels of β-catenin, a well-recognized cell survival factor in Wnt signaling and a substrate of GSK3β, and its phosphorylation. As shown (Fig. 3g, h), the level of β-catenin was decreased and its phosphorylation at Ser33/37 was increased in Dep rats. GSK3β and β-catenin was no change in the hippocampus of Res rats (Fig. 3g, h). These results suggested that ERα declined as one of key aspects of the occurrence of depression and cognitive deficits in Dep rats.
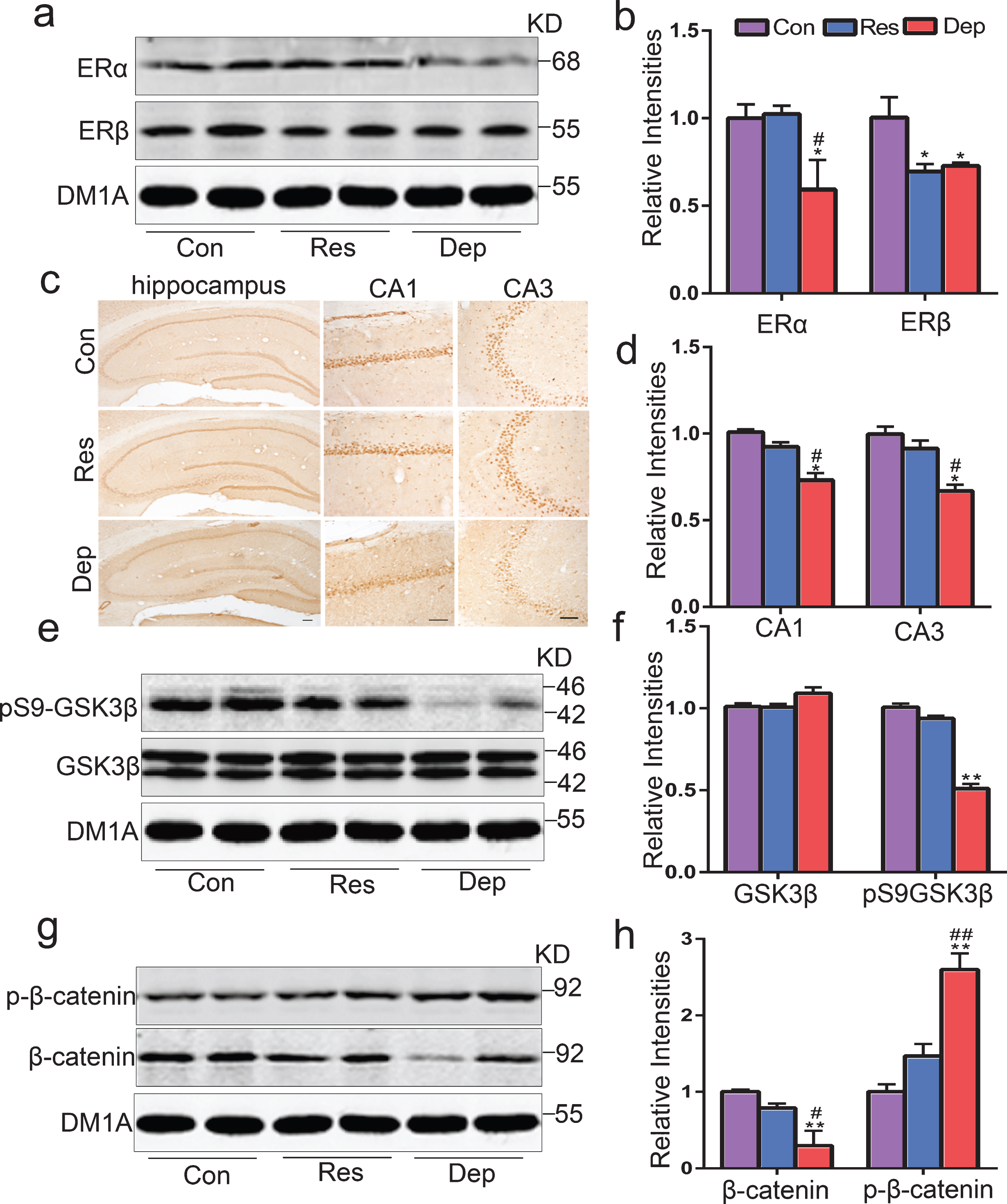
Hippocampal estrogen receptor α (ERα) loss in Dep rats, but not in Res rats. The levels of hippocampal ERα and ERβ were tested by western blotting (a) and quantitative analysis (b) (n = 5 rats/group). The distribution and expression of ERα in hippocampus were verified by immunohistochemistry staining (c) and quantitative analysis (d) (n = 3 rats/group) (Scale bar = 100μm). The levels of hippocampal glycogen synthase kinase3β (GSK3β) and phosphorylated GSK3β at Ser-9 (pS9-GSK3β), β-catenin and phosphorylated β-catenin at Ser33/37 were tested by western blotting (e) and quantitative analysis (f) (n = 5 rats/group). Data were presented as means±S.E.M. *p < 0.05, **p < 0.01 versus Con rats. #p < 0.05, # #p < 0.01 versus Res rats.
ERα agonist PPT rescued depressive behaviors and neuron loss in Dep rats
To further confirm the role of ERα, we treated Dep rats with PPT (1 mg/kg.d, Dep + PPT rats, n = 10) or equal volume of vehicle (Dep + Veh rats, n = 9) for 20 days as shown in Fig. 1a. In SPT, Dep + Veh rats had a lower sucrose preference than Dep + PPT rats (Fig. 4a, F2,29 = 12.33, p < 0.01). In FST, the immobility time of Dep + PPT rats was 54.49±15 s, much shorter than Dep + Veh rats (120.36±12 s, F2,29 = 8.53, p < 0.01) (Fig. 4b). In OFT, the crossing number (68.25±8.5) and the rearing number (24.75±4) of Dep + PPT rats were more than those of Dep + Veh rats (crossing number 35.22±3.2, F2,29 = 16.66, p < 0.01, rearing number 6.62±3.5, F2,29 = 10.9, p < 0.05) (Fig. 4c, d). In addition, we tested the learning and memory abilities of rats in MWM after treatment. In the learning phase, Dep + PPT rats had shorter latencies to find platform on 4th day (22.55±6.3 s, p < 0.05) and 6th day (17.875±2.3 s, p < 0.01) than Dep + Veh rats (39.96±7.8 s on 4th day, 28.96±2.8 s on 6th day). In the memory testing phase, the frequency of crossing platform in Dep + PPT rats was (3.8±0.48) more than Dep + Veh rats (1.25±0.7, p < 0.01) (Fig. 4e, f). Previous studies shown that PPT could revised neuron loss after ovariectomy [27–30]. In this study, NeuroN (a neuron-specific nuclear protein) based immunohistochemical staining was used to mark neuron (Fig. 5a). Compared with Dep + Veh rats, Dep + PPT rats had more neurons in the CA1 (F2,6 = 16.68, p < 0.05) and CA3 (F2,6 = 28.91, p < 0.001) regions of their hippocampi as Con rats (Fig. 5a, b). The studies supported that ERα, GSK3β, and β-catenin form a functional complex [33–36]. So, we used immunoprecipitation and western blotting to detect the levels of β-catenin combined with ERα and GSK3β. In the hippocampus of Res rats, the level of β-catenin combined with ERα was higher than that in Dep + Veh rats. And the Dep + PPT treated rats had the highest level of β-catenin combined with ERα (p < 0.05). Dep + Veh rats had the highest level of β-catenin combined with GSK3β in the hippocampus, which was significantly decreased in Dep + PPT rats (p < 0.01) (Fig. 5c–f). These results demonstrated the benefits of the increased binding of ERα with β-catenin in hippocampi of depressed rats.
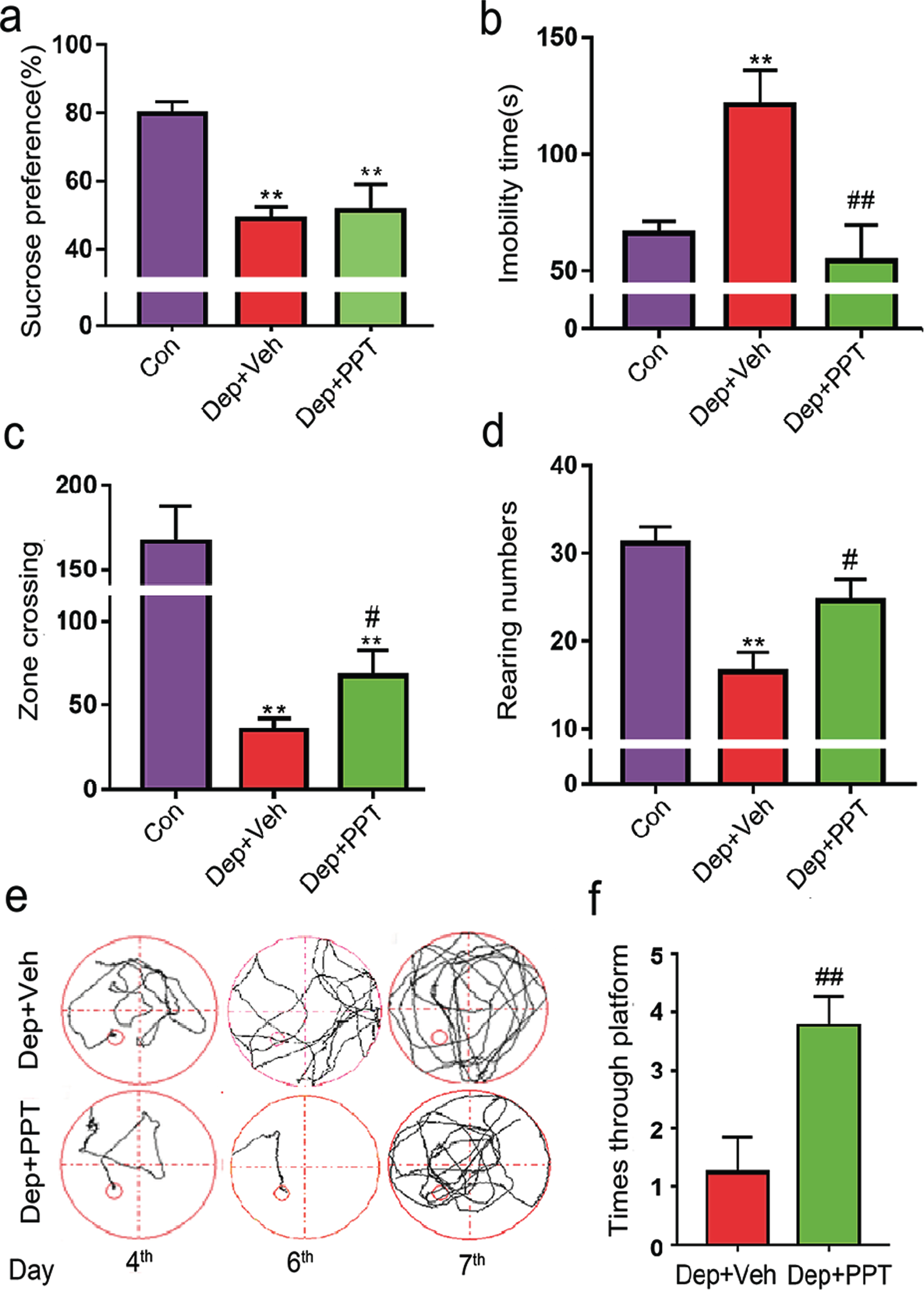
Treatment of PPT, an ERα agonist, rescued depressive and demential behaviors in Dep rats. Dep rats were daily treated with PPT (Dep + PPT, 1 mg/kg/day, n = 10) or vehicle treatment (Dep + Veh, daily subcutaneous injection with an equal volume of solvent, 50% DMSO/50% Dulbecco’s PBS) for 20 days. Then, the percentages of sucrose consumption in SPT (a), the immobility times in FST (b), the numbers of zone crossing (c), and the rearing numbers (d) in OFT of control rats (Con rats, n = 13), Dep + PPT rats (n = 10), and Dep + Veh rats (n = 9) were recorded. In the training phase of MWM, the swimming pathways (e), and the times crossing the hidden platform (f) from 2nd quadrant in the memory test of rats were recorded. The data were expressed as means±S.E.M. *p < 0.05, **p < 0.01 versus Con rats, #p < 0.05, # #p < 0.01 versus Dep + PPT rats.

PPT rescued the hippocampal neuron loss and increased the binding of ERα with β-catenin in Dep rats. The numbers of hippocampal neurons were shown by NeuroN (a neuron-specific nuclear protein) based immunohistochemical staining on brain slices (a) and quantitatively analyzed (b) (n = 3 rats/group, scale bar = 100μm). By immunoprecipitation and western blotting, the levels of β-catenin combined with ERα (c) and GSK3β (e) were tested and quantitatively analyzed (d, f). In the hippocampus of Res rats, the level of β-catenin combined with ERα was higher than that in Dep + Veh rats. And the Dep + PPT treated rats had the highest level of β-catenin combined with ERα (p < 0.05). Dep + Veh rats had the highest level of β-catenin combined with GSK3β in the hippocampus, which was significantly decreased in Dep + PPT rats (p < 0.01). The data were expressed as means±S.E.M (n = 3 rats/group). *p < 0.05, **p < 0.01 versus Con rats, #p < 0.05, # #p < 0.01 versus Res rats, Δ p < 0.05, ΔΔ p < 0.01 versus Dep + Veh rats.
To understand the interactions between the differential proteins in Dep/Con and β-catenin, ERα and GSK3β, we constructed a diagram present the interactions (Fig. 6). 11 upregulated proteins, e.g., versican (VCAN), Collagen Type 1 α chain 1 (Col1a1), Col1a2, superoxide dismutase 1 (Sod1), solute carrier family 1 (glial high affinity glutamate transporter) member 2 (Slc1a2), Tropomyosin 1 (Tpm1), transgelin (Tagln), myosin heavy chain 11 (Myh11), keratin 8 (Krt8), Krt18 and filamin A (Flna), and downregulated calcium/calmodulin kinase IV (Camk4) are β-catenin and ERα associated. The downregulated Axin1, lymphoid enhancer-binding factor 1 (Lef1) and adenomatous polyposis coli (APC) are β-catenin, GSK3β, and ERα associated. These 6 proteins are more closely related to each other (Fig. 6). β-catenin is also associated with the upregulated cluster of differentiation 82 (Cd82) and downregulated E3 ubiquitin-protein ligase BRE1-like protein (Hub), guanosine-5’-monophosphate reductase (Gmpr), myristoylated-alanine rich C-kinase substrate (Marcks), pleiotrophin (Ptn) and Heterogeneous nuclear ribonucleoprotein U-like protein 1 (Hnrnpul1). Caldesmon 1(Cald1), RNA-binding motif protein 39 (Rbm39), ferritin (Fth1), transferrin receptor protein 1 (Tfrc), α2-HS glycoprotein (Ahsg), Ras-related protein 2 (Rras2), tubulin β 2B class IIB (Tubb2b) and hemopexin (Hpx) are ERα associated (Fig. 6).
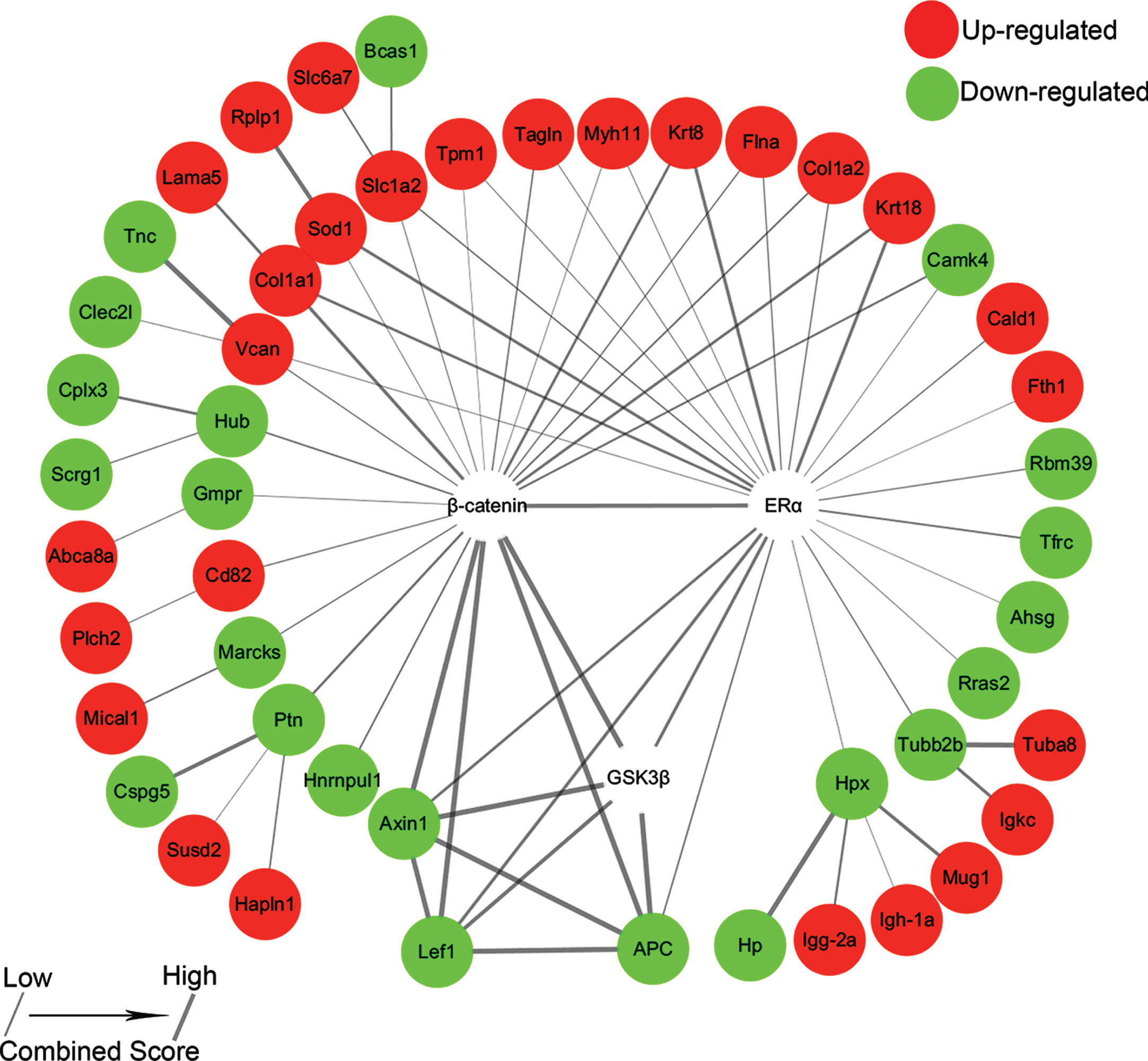
The interaction network of ERα, GSK3β, and β-catenin and their associated differentially expressed proteins in Dep/Con. Red denotes upregulation, while green denotes downregulation. The higher values of the combined score indicate the stronger of protein-protein interactions.
DISSCUSION
This study sought to elucidate the role of ERα in CUMS-induced depression and cognitive deficits in young female rats. We demonstrated that dysregulation of ERα/Wnt signalings in the hippocampus is one key pathological link and ERα is an important pro-resilient factor. During aging, declined ERα in the hippocampi of aged female rats or patients with AD [62, 63] and schizophrenics [62–64], whereas hippocampal ERβ immunoreactivity was increased in AD patients [65]. ERβ deficiency was found impair the brain-derived neurotrophic factor (BDNF) and serotonin (5-HT) signalings in the hippocampus of the female brain, suggesting a possible mechanism for menopausal depression [66]. No differences were found recently in the ERα and ERβ levels in female mice subjected to chronic social instability stress with anxiety and depressive-like behavior [67].
In this study, hippocampal ERβ decreased in Dep rats and Res rats after CUMS, while decreased ERα was only observed in the hippocampi of Dep rats. Some studies in receptor knockout mice indicated that ERα maintains hippocampal function, particularly when estrogen levels are low, and ERβ might have an inhibitory effect on ERα-mediated effects [68]. All these data suggested the complicated roles of hippocampal ERα and ERβ in depression and dementia.
In hippocampus ERα and ERβ are both predominately expressed. However, ERα is reported transcriptionally more active than ERβ [68], and low estrogen level will preferentially activate ERα-dependent transcription [69]. Women tend to be depressed with low estrogen level [70–72], and the depressed patients show reduction in hippocampal volumes, which could be revised by anti-depressant treatments [5, 73]. Recent studies found there is a 50% reduction of synapses that contain ERα in the CA1 region of rat hippocampus, and the ability of ERT to induce spine formation declines with age and is associated with ERα loss [74, 75]. Like ERα, the level of synaptic ERβ decreases with age, but increases following ERT [76]. In this study, hippocampal ERβ decreased both in Dep rats and Res rats after CUMS. Hippocampal ERα decreased in Dep rats accompanied with neuron loss, and ERα agonist revised this neuron loss. Thus, ERα maybe a key target in hippocampal neuronal loss of depression.
GSK3β and β-catenin are two important molecules in Wnt signaling, which is critical for normal functioning of hippocampus [33–36]. Cytosolic β-catenin protein is the principal mediator of Wnt signaling. Axin acts as a scaffold, in the absence of Wnt signaling, for the cytosolic β-catenin destruction complex, including β-catenin, APC, and the serine/threonine kinase GSK3β. APC is an essential negative regulator of Wnt signaling. In the absence of ligands, β-catenin is recruited into a complex comprising Axin, APC, and GSK-3β, facilitating GSK-3β to phosphorylate β-catenin and target it for ubiquitination and degradation to keep the low level of cytosolic β-catenin. Lef1 is a downstream mediator of the Wnt/β-catenin signaling pathway. If the formation of Axin-APC-GSK-3β complex is disturbed, GSK-3β-mediated phosphorylation of β-catenin is inhibited and the non-phosphorylated β-catenin translocates into the nucleus to binds to the N terminus of Lef1, and recruits co-factors to promote the transcription of Wnt target genes. Many of the Wnt target genes are responsible for several cellular processes such as anti-apoptosis, neurogeneration, and synaptic plasticity. In this study, the increased phosphorylated β-catenin and decreased total β-catenin were observed in the hippocampi of Dep + Veh rats, in keeping with the neuron loss and hippocampal dysfunction in Dep rats and also indicating the dysregulation of Wnt signaling. There are close connections between Wnt and estrogen signaling pathways. Estrogen signaling associated neuronal development is mediated through the activation of the Wnt signaling. Recent reports showed ERα forms a protein complex with GSK3β and β-catenin in the neurons of hippocampus from OVX rats. And estradiol addition dissociated β-catenin from this complex, which was correlated with GSK3 inhibition [33–36]. By activating phosphoinositide 3-kinase (PI3K)/Akt signaling pathway through ERs, estradiol activated the transcriptional activity of Wnt/β-catenin signaling by inhibiting GSK-3β in neuroblastoma cells and primary cortical neurons [36]. ASPP 049, a phytoestrogen, induced the proliferation and differentiation of osteoblastic cells through ERα-, Akt-, and GSK-3β-dependent activation of β-catenin [76]. In this research, Dep + Veh rats had the lowest level of β-catenin combined with ERα and the highest level of β-catenin combined with GSK3β in their hippocampi. Conversely, PPT treated rats had the highest level of β-catenin combined with ERα and the lowest level of β-catenin combined with GSK3β. Thus, it was given that the neuroprotection afforded by hippocampal ERα/Wnt interactions have significant implications for cognition and emotion in females. Previously, we found the phosphorylation of ERα at Ser118 could be regulated by GSK3β [61]. In this research, although decreased hippocampal ERα was shown in Dep rats, the slight decrease in electrophoretic mobility of ERα in Dep rats (Fig. 3a) indicated that ERα might be phosphorylated by active GSK3β. In our previous research [40], CUMS-induced female Dep rats were found have 107% increase of hippocampal tau phosphorylation at Ser396, 24% increase at Ser404, and 608% increase at Thr205, and a 41% decrease of the dephosphorylated tau. Furthermore, these female Dep rats also had activated GSK-3β and JNK in their hippocampi. If the tau hyperphosphorylation in female Dep rats is GSK3β-dependent needs further investigation.
In conclusion, CUMS induced depressed female rats showed obvious cognitive deficits with the declined level of ERα, neuron loss, upregulated cleaved caspase-3, activation of GSK-3β and increased phosphorylation of β-catenin in the hippocampus. It was further demonstrated that ERα is an important pro-resilient factor in CUMS-induced depression and dementia in young female rats. The findings provide evidence for a possible mechanism underlying the increased susceptibility of depression in young female, and a possible target for the prevention of depression and dementia in women.
Footnotes
ACKNOWLEDGMENTS
This work was supported by grants from the National Natural Science Foundation of China (81960260, 91539112, 8140086), Health and Family Planning Scientific Research Project of Hubei Province (WJ2015Q033), Population and Family Planning Commission of Wuhan (WX14B34), the Science and Technology Department of Jiangxi Province (20192BAB215022), Award and Fellowships from Wuhan Young & Middle-Aged Talents, Health and Family Planning Commission of Wuhan Municipality and China Scholarship Council File (NO.201608420019).