
Abstract
Background:
Alzheimer’s disease (AD) is an inflammatory neurodegenerative disease that may be associated with prior bacterial infections. Microbial “old friends” can suppress exaggerated inflammation in response to disease-causing infections or increase clearance of pathogens such as Mycobacterium tuberculosis, which causes tuberculosis (TB). One such “old friend” is Mycobacterium vaccae NCTC 11659, a soil-derived bacterium that has been proposed either as a vaccine for prevention of TB, or as immunotherapy for the treatment of TB when used alongside first line anti-TB drug treatment.
Objective:
The goal of this study was to use a hypothesis generating approach to explore the effects of M. vaccae on physiological changes in the plasma and cerebrospinal fluid (CSF).
Methods:
Liquid chromatography-tandem mass spectrometry-based proteomics were performed in plasma and CSF of adult male rats after immunization with a heat-killed preparation of M. vaccae NCTC 11659 or borate-buffered saline vehicle. Gene enrichment analysis and analysis of protein-protein interactions were performed to integrate physiological network changes in plasma and CSF. We used RT-qPCR to assess immune and metabolic gene expression changes in the hippocampus.
Results:
In both plasma and CSF, immunization with M. vaccae increased proteins associated with immune activation and downregulated proteins corresponding to lipid (including phospholipid and cholesterol) metabolism. Immunization with M. vaccae also increased hippocampal expression of interleukin-4 (IL-4) mRNA, implicating anti-inflammatory effects in the central nervous system.
Conclusion:
M. vaccae alters host immune activity and lipid metabolism. These data are consistent with the hypothesis that microbe-host interactions may protect against possible infection-induced, inflammation-related cognitive impairments.
Keywords
INTRODUCTION
Mycobacterium vaccae NCTC 11659 is a bac-terium that is under development as an anti-tuber-culosis (TB) vaccine that presumably works through a mechanism similar to the Bacillus Calmette-Guérin (BCG) TB vaccine [1]. Treatment with the BCG vaccine, derived from an attenuated strain of Mycobacterium bovis, is associated with the prevention and treatment of Alzheimer’s disease (AD) in human studies and animal models [2, 3]. However, as a vaccine, although BCG protects against the more severe forms of Mycobacterium tuberculosis infection, it falls short otherwise [4]. M. vaccae NCTC 11659 is protective against cognitive decline such as found in AD [5], and numerous trials have indicated that exposure to oral and injectable products derived from M. vaccae bacteria can have positive effects in treating TB [6–8]. M. vaccae NCTC 11659 has been proposed either as a vaccine for prevention of TB [9–12], or as immunotherapy for the treatment of TB when used alongside first line anti-TB drug treatment [6–8, 13].
The relationships between humans and microorganisms that constitute the human microbiome may play an important role in the regulation of inflammatory responses in the host [14–22]. The “old friends” hypothesis proposes that exaggerated or inappropriate inflammation may be due to reduced contact with certain microorganisms that drive anti-inflammatory and immunoregulatory responses, such as some species of bacteria that have been associated with humans throughout evolutionary history [14–22]. “Old friends” are microorganisms that may, in some cases, dampen inflammatory processes of the immune system caused by contact with pathogenic or sterile inflammatory stimuli (which may include “self” antigens, stress-induced damage-associated molecular patterns, or microbe-associated molecular patterns) [23, 24]. It is thought that several mycobacteria species, including the soil-derived bacterium Mycobacterium vaccae NCTC 11659, are “old friends” [5, 25–31].
One way that “old friends” may inhibit inflammation is by suppressing immune responses and promoting the differentiation of regulatory T cells (Tregs) [25, 32]. Previous studies report that immunization with a nonpathogenic, immunologically active, heat-killed preparation of M. vaccae NCTC 11659 induces Treg subsets in the spleen, enhances anti-inflammatory cytokine signaling in the periphery and the brain, and prevents stress-induced microglial priming [5, 31]. Interestingly, these anti-inflammatory and immunoregulatory effects correspond to decreased stress-induced behaviors, implicating microbe-host pathways that promote stress resilience. For example, immunization with a heat-killed preparation of M. vaccae NCTC 11659 consistently reduces stress-induced anxiety- and depressive-like behavioral responses in rodents [26–29, 33], and depletion of Tregs can block the anxiolytic effects of M. vaccae treatment in a chronic subordinate colony housing stress model in mice [26]. As with other “old friends,” the stress-resilience effects of M. vaccae NCTC 11659 may work in part through anti-inflammatory and Treg-dependent, immunoregulatory mechanisms [25, 26].
Due to their anti-inflammatory and immunoregulatory properties, “old friends”, including some commensal microorganisms, have become an exciting new avenue of study in the etiology and treatment of inflammatory disorders, including infection-related neurodegenerative diseases like AD [34–36]. “Old friends” may suppress exaggerated inflammation in response to disease-causing infections [19, 37], or increase clearance of pathogens [8, 38–41], and therefore may be important for the prevention and treatment of disorders like AD. Indeed, decreased exposure to a rich microbial environment has been linked to increased risk of AD development worldwide [35]. Considering AD pathology is inflammatory in nature [42–48], persons with AD exhibit dysregulated Treg phenotypes [49, 50], and Treg-focused therapies may mitigate AD symptoms [51–53], “old friends” might turn out to be promising novel therapeutics. Interestingly, treatment with the BCG vaccine, derived from an attenuated strain of Mycobacterium bovis, is associated with the prevention and treatment of AD in human studies and animal models [2, 3]. It is worth noting that AD is also closely linked to inflammatory metabolic diseases, including type 1 and type 2 diabetes mellitus (T1D and T2D, respectively) [42, 54–58], which also present with impairments in Treg function [59, 60]; thus, it could be that improper metabolic functions help to drive Treg dysfunction in AD and comorbid inflammatory conditions [61]. Of particular relevance, immunization with M. vaccae NCTC 11659 induces anti-inflammatory and immunoregulatory signaling, prevents priming of microglia in the hippocampus, and prevents age-related cognitive impairments in a model of post-operative cognitive dysfunction in aged rats [5]. Although studies of mycobacteria and other “old friends” in the context of age-related cognitive impairments and neurodegenerative diseases are fairly new, the effects of mycobacteria (especially M. tuberculosis infection) on immune function and metabolic signaling are already well described [62–65]. Of course, physiological effects of mycobacterial species differ depending on the strain, live versus heat-killed preparation, mode of administration, and virulence, in the case of live bacteria, as well as the genome, previous environmental exposures, and health of the individual host [66–68].
While the interactions between mycobacterial “old friends” and host physiology and behavior have been characterized, the mechanisms by which M. vaccae NCTC 11659 exerts its effects remain largely unknown. Hypothesis-generating proteomics studies are useful to provide exploratory analyses of proteome profiles across biological systems in a given species [69]. In this study, we use liquid chromatography-tandem mass spectrometry (LC-MS/MS)-based proteomics to identify a number of proteins in plasma and cerebrospinal fluid (CSF) that are directly altered by a series of subcutaneous injections of a heat-killed preparation of M. vaccae NCTC 11659 given to rats, when assessed one week following the final immunization. We follow up our proteomics analysis with assessment of mRNA expression of genes associated with immune and metabolic function in the hippocampus. This is the first study to show long lasting changes in the plasma and CSF proteome of rats treated with M. vaccae NCTC 11659, coinciding with increased anti-inflammatory gene expression in the brain. These results help inform the potential mechanisms through which M. vaccae NCTC 11659 exerts its anti-inflammatory, immunoregulatory, and stress resilience effects and support future studies of heat-killed preparations of mycobacteria as novel approaches to prevention of, and therapeutic strategies for, neurodegenerative disorders, including AD.
MATERIALS AND METHODS
Animals
For an experimental timeline, please see Fig. 1. Adult male Sprague Dawley® rats (Hsd:Sprague Dawley® SD®; Envigo, Indianapolis, IN, USA) weighing 250–265 g upon arrival were pair-housed in Allentown micro-isolator filter-topped caging [259 mm (W)×476 mm (L) ×209 mm (H); cage model #PC10198HT, cage top: #MBT1019HT; Allentown, NJ, USA] containing an approximately 2.5 cm-deep layer of bedding (Cat. No. 7090; Teklad Sani-Chips; Harlan Laboratories, Indianapolis, IN, USA). This species, strain, and supplier were chosen for proteomics analysis due to previous M. vaccae NCTC 11659 studies conducted with these animals [27–29, 31]. All rats were kept under standard laboratory conditions (12 h light/dark cycle, lights on at 0700 h, 22°C) and had free access to bottled reverse-osmosis water and standard rat diet (Harlan Teklad 2918 Irradiated Rodent Chow, Envigo, Huntingdon, United Kingdom). Cages were changed once per week. The research described here was conducted in compliance with The ARRIVE Guidelines for Reporting Animal Research [70], and all studies were consistent with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, Eighth Edition [71]. The Institutional Animal Care and Use Committee at the University of Colorado Boulder approved all procedures. All efforts were made to limit the number of animals used and their suffering.
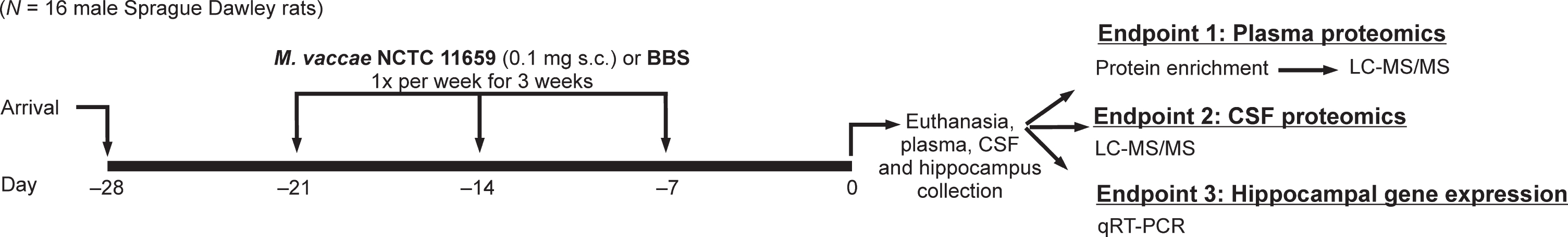
Diagrammatic illustration of experimental design and protocol; sample sizes (N = 16; borate-buffered saline (BBS) vehicle, n = 8; M. vaccae NCTC 11659, n = 8). BBS, borate-buffered saline; CSF, cerebrospinal fluid; LC-MS/MS, liquid chromatography-tandem mass spectrometry; qRT-PCR, quantitative reverse transcription polymerase chain reaction; s.c., subcutaneous.
Reagents
This study used a whole cell heat-killed preparation of M. vaccae NCTC 11659 [IMM-201; alternative designations and different preparations and production processes of M. vaccae NCTC 11659 used in clinical trials or preclinical studies include: DAR-901 [10], DarDar tuberculosis vaccine [11], MV001 [9], MV 007 [12], M. vaccae SRL 172 [11], M. vaccae SRP 299 [72]; V7 [8] is a hydrolyzed version of heat-killed M. vaccae NCTC 11659]; 10 mg/ml solution; strain National Collection of Type Cultures (NCTC) 11659, batch C079-ENG#1, provided by BioElpida (Lyon, France), diluted to 1 mg/ml in 100μl sterile borate-buffered saline (BBS) for injections].
M. vaccae and vehicle immunization
Experimental rats received either subcutaneous (s.c.) immunization with 0.1 mg whole heat-killed M. vaccae NCTC 11659 suspension or received s.c. injections of 100μl of the vehicle, sterile BBS, using 21-gauge needles and injection sites between the scapulae, between the hours of 12 pm and 4 pm. The dose used in these experiments (0.1 mg) was 1/10 of the dose used in human studies (1 mg) [73] and identical to the dose used in previous studies in mice and rats [5, 72].
Euthanasia, tissue, cerebrospinal fluid, and plasma collection
Rats were euthanized one week following the last injection using an overdose of sodium pentobarbital (Fatal Plus®, Vortech Pharmaceuticals Ltd., Dearborn, MI, USA; 200 mg/kg, i.p.). Immediately after euthanasia, CSF was collected as described previously [74]. Briefly, the dorsal aspect of the skull was shaved and swabbed with 70% ethanol. A sterile 26-gauge needle attached via PE50 tubing to a sterile 1 mL syringe was inserted into the cisterna magna, and 0.2 mL of clear CSF was drawn into the syringe. CSF was then spun down at 1000×g for 10 min, and the supernatant was collected and stored at –80°C. Due to loss of samples during CSF collection, final sample sizes for CSF analysis were vehicle, n = 5, M. vaccae NCTC 11659, n = 6. Blood was collected after CSF but prior to perfusion by inserting a 21-gauge needle into the right atrium of the heart and slowly drawing up 1 mL of blood. Blood was gently released into a BD Vacutainer® blood collection tube containing EDTA (Cat. No. 367835; Becton, Dickinson, and Company; Franklin Lakes, NJ, USA) and spun down at 1500×g for 10 min at 4 °C; the plasma supernatant was collected and stored at –80°C. Due to loss of one sample during plasma collection, final sample sizes for plasma analysis were vehicle, n = 7, M. vaccae, n = 8. After blood collection, rats were then perfused with ice-cold 0.9% saline to remove peripheral leukocytes from central nervous system (CNS) vasculature. After perfusion, whole hippocampus was dissected from both left and right hemispheres, stored separately, and immediately placed in liquid nitrogen; hippocampi were stored at –80°C.
Protein enrichment of plasma
Protein enrichment allows for greater resolution of low abundance proteins in downstream analyses of plasma samples. Samples were thawed, and 200 μL was taken for enrichment of low-abundance proteins using ProteoMiner™ Small Capacity Kit (Catalog #163-3006, Bio-Rad Laboratories, Inc., Hercules, CA, USA). Samples were immediately processed for liquid chromatography-tandem mass spectrometry (LC-MS/MS) at the Central Analytical Mass Spectrometry Facility, University of ColoradoBoulder.
Liquid chromatography-tandem mass spectrometry (LC-MS/MS): Plasma sample processing
Plasma samples were processed as described previously [75]. Briefly, 8 M urea in samples was diluted to 2.7 M with 50 mM triethylammonium bicarbonate (TEAB), and samples were reduced and alkylated by incubating with 5 mM tris(2-carboxyethyl)phosphine (TCEP) and 20 mM chloroacetamide at 80°C for 15 min in darkness. Acetonitrile was added to a final concentration of 70% and Sera magnetic beads (GE Healthcare Life Science, Pittsburgh, PA, USA) were added to a final concentration of 0.5 μg/ μL. Beads were washed with 1 mL acetonitrile and 1 mL 70% ethanol and resuspended in 200 μL of 50 mM TEAB. Lys-C was added to 0.5% (w/w) and incubated at 37°C for 1 h, and then trypsin was added to 2% (w/w) and incubated at 37°C overnight. An equal volume of ethyl acetate containing 2% formic acid was added to the sample and mixed vigorously, and the mixture was loaded onto in-house-packed Stop-and-Go-Extraction (STAGE) tips (styrenedivinylbenzene-reverse phase sulfonate (SDB-RPS) membrane) [76]. Samples were washed twice with ethyl acetate containing 2% formic acid and twice with Buffer A (0.1% formic acid in water), and eluted using Buffer X (80% acetonitrile, 5% ammonium hydroxide). The desalted peptides were dried using a vacuum centrifugation. Tandem mass tag (TMT)-labeling (6-plex) of peptides was performed as described in Zecha et al. (2019) [77].
Liquid chromatography-tandem mass spectrometry (LC-MS/MS): Cerebrospinal fluid sample processing
CSF (10 μg protein in 50 μL from each animal) was diluted in 150 μL of 0.1 M ammonium bicarbonate (ABC), 0.01% sodium deoxycholate, 5 mM TCEP, 20 mM chloroacetamide, and incubated at 80°C for 15 min in darkness. Samples were digested with 0.2 μg (2% w/w) of Lys-C/trypsin mixture (Promega, Madison, WI, USA) at 37°C overnight. Desalting was performed the same as for plasma.
LC-MS/MS analysis
The TMT-labeled, tryptic peptides were resolved using a Waters nanoACQUITY UPLC system in a single pump trap mode. The peptides were loaded onto a nanoACQUITY 2G-V/MTrap 5 μm Symmetry C18 column (180 μm×20 mm) with 95% Buffer A and 5% Buffer B (0.1% formic acid in acetonitrile) at 15 μL/min for 7 min. The trapped peptides were eluted and resolved on a BEH C18 column (130 Å, 1.7 μm×75 μm×250 mm) using gradients of 5% to 30% B (0–145 min) and 30% to 60% B (145–150 min) at 0.3 μL/min. MS/MS was performed on a LTQ Orbitrap Velos mass spectrometer, scanning precursor ions between 400 and 1800 m/z (1×106 ions, 60,000 resolution) and selecting the 6 most intense ions for MS/MS with 180 s dynamic exclusion, 10 ppm exclusion width, repeat count = 1, and 30 s repeat duration. Ions with unassigned charge states and MH+1 were excluded from the MS/MS. Higher-energy collisional dissociation (HCD) MS/MS (5×104 ions, 7,500 resolution) was performed with a default charge state = 5, isolation width = 2 m/z, normalized collision energy = 40, and activation time = 1 ms. Maximal ion injection times were 500 ms.
Protein identification analysis
MaxQuant/Andromeda (version 1.6.2.10) was used to process raw files from LTQ-orbitrap with a database consisting of UniProt Rattus norvegicus proteome (total 36,098 entries, downloaded on 2/15/2019) [78–80]. All protein names and gene abbreviations were gathered from the UniProt database [80]. A preset “6-plex TMT” was used. The search allowed trypsin specificity with a maximum of two missed-cleavage, setting carbamidomethyl modification on cysteine as a fixed modification and protein N-terminal acetylation and oxidation on methionine as variable modifications. MaxQuant used 4.5 ppm main search tolerance for precursor ions, 20 ppm HCD MS/MS match tolerance, searching the top 12 peaks per 100 Da. False discovery rates for both protein and peptide were 0.01 with a minimum peptide length of seven amino acids. “Reporter ion corrected” values in proteinGroups.txt were used for downstream statistical analysis.
Real time RT-qPCR semi-quantification of hippocampal mRNA expression
Real time RT-qPCR was performed as described previously [81]. Total RNA was isolated from the left whole hippocampus utilizing a standard method of phenol:chloroform extraction [82]. cDNA sequences were obtained from Genbank at the National Center for Biotechnology Information (NCBI; http://www.ncbi.nlm.nih.gov) [83]. Primer sequences were designed using the Operon Oligo Analysis Tool (http://www.operon.com/technical/toolkit.aspx) or the Integrated DNA Technologies (IDT) PrimerQuest tool (https://www.idtdna.com/pages/tools/primerquest) and tested for sequence specificity using the Basic Local Alignment Search Tool (BLAST) at NCBI [84]. Primers were obtained from Invitrogen (Actb, Cd200, Cd200r1, Il4, Il10, Mrc1 (Cd206), Tgfb1) or IDT (Apoa1, Apob, Apoc2, A1m, Ckm, Igg-2a, Itih1, Ppara, Ppard, Pparg). Primer specificity was verified by melt curve analyses. All primers were designed to span exon/exon boundaries and thus exclude amplification of genomic DNA (see Supplementary Table 1 for primer descriptions and sequences). PCR amplification of cDNA was performed using the QuantiTect SYBR Green PCR Kit (Cat. No. 204056, Qiagen, Hilden, Germany). Formation of PCR product was monitored in real time using the MyiQ Single-Color Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Relative gene expression was determined by taking the expression ratio of the gene of interest to Actb, encoding β-actin.
Statistical analysis
Proteomics analyses were performed separately for plasma and CSF samples using R Statistical Programming (version 3.6.1 for Windows). Abundance of each protein was measured by reporter ion-corrected values. To correct for potential noise in the MS/MS detection, reporter ion-corrected values were removed for any proteins that had a reporter intensity count less than two. Corrected peak intensities for each protein were then normalized to the total of all peak intensities for that sample. For fold change analysis, normalized peak intensities of each protein were averaged for each treatment group, and ratios of treatment to control for each protein were created. To control for batch effect of the 6-plex TMT, generalized linear model (GLM) analysis was performed on log2-transformed normalized peak intensity for each protein, with treatment and batch as factors. Two-tailed significance was set at α= 0.1, and an adjusted p-value was not calculated due to low power [85]. For the full written R code, please see Supplementary Materials. Statistical analysis was not performed on a protein if a group sample size for that protein was less than 50% of the full sample size for that treatment group. Proteins that were found to be significantly different between treatment and control as well as proteins with a fold-change (up or downregulated) greater than two were used in downstream pathway analysis via the Database for Annotation, Visualization and Integrated Discovery (DAVID, version 6.8) using UniProt accession IDs for gene labeling and Rattus norvegicus as species. For protein interaction analysis, STRING database (version 11) was used [86].
Statistical analysis for hippocampal mRNA expression was performed using the software package IBM Statistical Package for the Social Sciences (version 26.0, SPSS Inc., Chicago, IL, USA). Normality was assessed using the Shapiro-Wilks Test for Normality; if the data were found to be non-normal, the data were log-transformed and normality reassessed for confirmation of statistical assumptions. Independent-sample t-tests were run on normalized data, and homogeneity of variance was analyzed by Levene’s Test for Equality of Variance. If variance was unequal, the adjusted p-value (obtained from Welch’s t-test) from the SPSS output was used.
RESULTS
Plasma proteomics
Among 139 proteins identified in plasma, no proteins were found to be upregulated or downregulated by two-fold change after M. vaccae treatment. However, GLM analysis found a significant difference in abundance of three out of 139 proteins. M. vaccae treatment decreased the abundance of apolipoprotein C-II (ApoC-II, p = 0.099), and increased the abundance of serine proteinase inhibitor, clade A, member 4 (Serpin A4, p = 0.039) and Ig gamma-2A chain C region (IgG-2a, p = 0.078) (Fig. 2). The top 15% of proteins of interest in the plasma, determined by p-value, can be found in Supplementary Table 2.
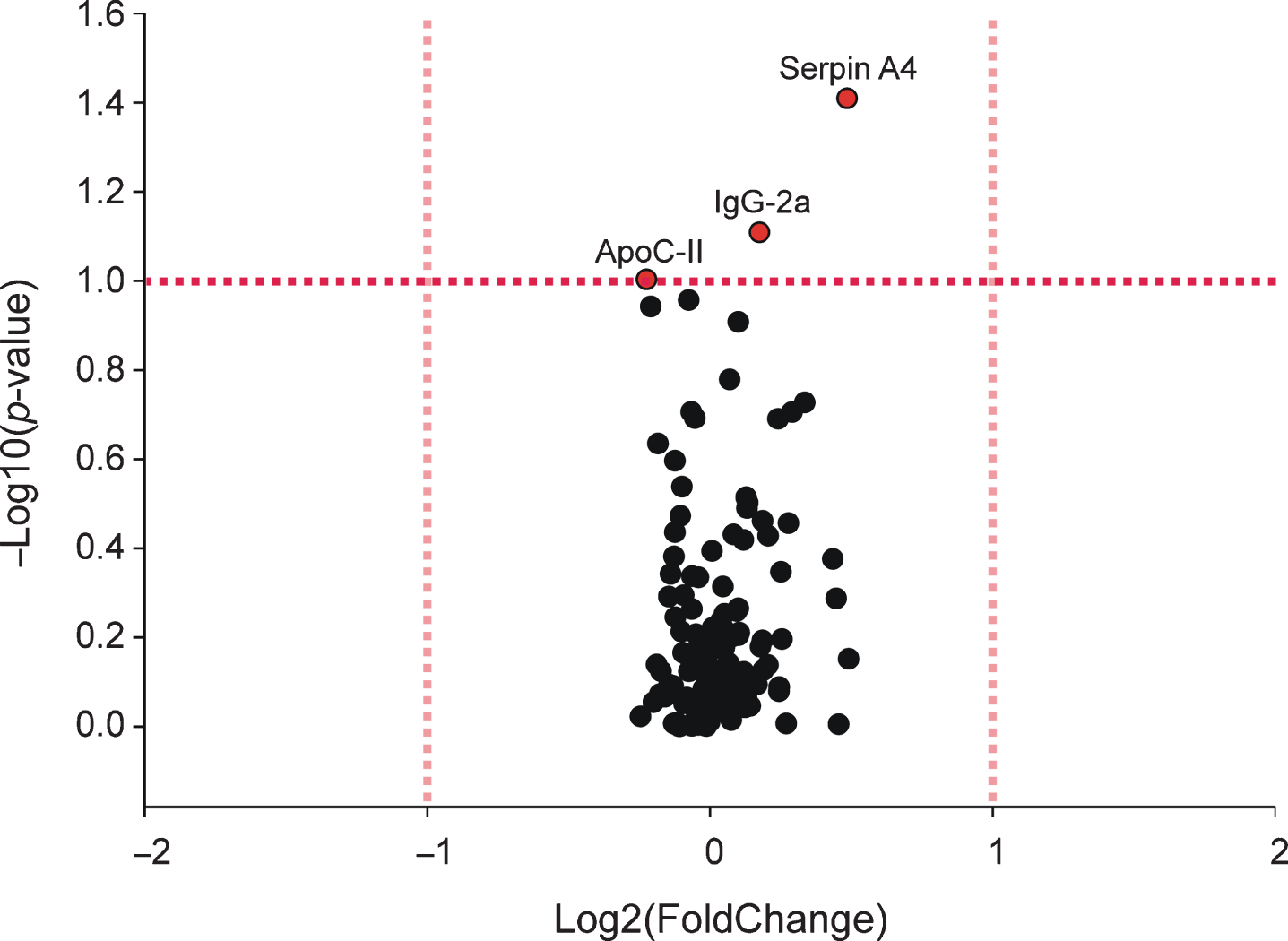
Volcano plot showing effects of Mycobacterium vaccae NCTC 11659 treatment on plasma proteomics as measured by liquid chromatography-tandem mass spectrometry (LC-MS/MS) seven days following the final immunization. The pink vertical dashed lines represent the thresholds for greater than two-fold change in average peak intensity, whereas the red horizontal dashed line represents the threshold for significance at a p-value less than 0.1, corresponding to proteins labeled red. Final sample sizes for plasma analysis were vehicle, n = 7, M. vaccae NCTC 11659, n = 8. ApoC-II, apolipoprotein C-II; IgG-2a, Ig gamma-2A chain C region; Serpin A4, serine proteinase inhibitor, clade A, member 4.
Plasma protein-protein associations and functional enrichment analysis
STRING analysis revealed a significant number of protein interactions, with three nodes and one association edge (PPI enrichment p-value = 0.00789) (Fig. 3), implicating these proteins in biologically related pathways. Text mining and co-expression associations were found between ApoC-II and Serpin A4. However, there were no functional enrichments found among the proteins.
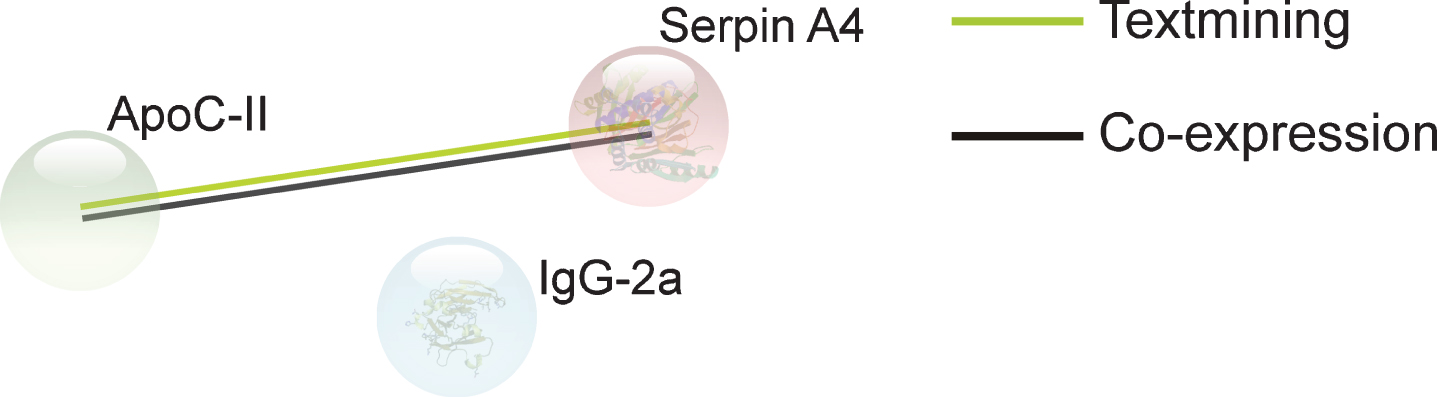
Protein associations in the plasma. Proteins presented are those whose abundance was altered (upregulated or downregulated) by M. vaccae NCTC 11659 treatment, as assessed seven days following the final immunization. Associations were computed by STRING database (v.11). Final sample sizes for plasma analysis were vehicle, n = 7, M. vaccae NCTC 11659, n = 8. ApoC-II, apolipoprotein C-II; IgG-2a, Ig gamma-2A chain C region; Serpin A4, serine proteinase inhibitor, clade A, member 4.
CSF proteomics
We then tested whether the proteomics profile was altered in the CSF of the same rats. Among 199 proteins identified in CSF, our preliminary average fold change analysis revealed that M. vaccae treatment increased the abundance of serine protease inhibitor A3N (Serpin A3N) by two-fold (Fig. 4). Additionally, based on fold change analysis, M. vaccae treatment decreased the abundance of six proteins by two-fold: i.e., alpha-1-macroglobulin (Alpha-1-M, also known as pregnancy-zone protein, or PZP), apolipoprotein A-I (ApoA-I, also ApoAI) [87], apolipoprotein B-100 (ApoB-100), creatine kinase M-type (M-CK), carboxypeptidase N subunit 2 (CPN2), and inter-alpha-trypsin inhibitor, heavy chain 1 (ITIH1) (Fig. 4).

Volcano plot showing effects of Mycobacterium vaccae NCTC 11659 treatment on cerebrospinal fluid (CSF) proteomics as measured by liquid chromatography-tandem mass spectrometry (LC-MS/MS) seven days following the final immunization. The pink vertical dashed lines represent the thresholds for greater than two-fold change in average peak intensity, corresponding to proteins labeled pink, whereas the red horizontal dashed line represents the threshold for significance at a p-value less than 0.1, corresponding to proteins labeled red. Green-labeled proteins represent those that were both increased or decreased by two-fold change in peak intensity and significant at a p-value less than 0.1. Final sample sizes for CSF analysis were vehicle, n = 5, M. vaccae NCTC 11659, n = 6. Alpha-1-M, alpha-1-macroglobulin; ApoA-I, apolipoprotein A-I; ApoB-100, apolipoprotein B-100; C1s, complement C1s subcomponent; CD5L, CD5 antigen-like; M-CK, creatine kinase, M-type; CPN2, carboxypeptidase N subunit 2; IgG-2a, Ig gamma-2A chain C region; IGFBP-3, insulin-like growth factor-binding protein 3; ITIH1, inter-alpha trypsin inhibitor, heavy chain 1; SgII, secretogranin-2; Serpin A3N, serine protease inhibitor A3N; Serpin A4, serine proteinase inhibitor, clade A, member 4.
GLM analysis found a significant difference in abundance of nine out of 199 proteins between M. vaccae- and vehicle-treated animals. M. vaccae treatment increased the abundance of complement C1s subcomponent (C1s, p = 0.050), insulin like growth factor binding protein 3 (IGFBP-3, p = 0.093), Ig gamma-2A chain C region (IgG-2a, p = 0.095), and secretogranin-2 (SgII, p = 0.079) (Fig. 4). M. vaccae treatment decreased apolipoprotein B-100 (ApoB-100, p = 0.044), CD5 molecule like (CD5L, p = 0.088), creatine kinase, M-type (M-CK, p = 0.091), carboxypeptidase N subunit 2 (CPN2, p = 0.066), and serine proteinase inhibitor, clade A, member 4 (Serpin A4, p = 0.094) (Fig. 4). ApoB-100, M-CK, and CPN2 were found to be downregulated by M. vaccae treatment in both fold change and GLM analyses (Fig. 4). The top 15% of proteins of interest in the CSF, determined by p-value, can be found in Supplementary Table 3.
Importantly, M. vaccae treatment increased the abundance of IgG-2a in both plasma and CSF.M. vaccae treatment increased the abundance of Serpin A4 in the plasma but decreased the abundance of Serpin A4 in the CSF. ApoC-II, which was increased in plasma, was not detected in CSF.
CSF protein-protein associations and functional enrichment analysis
STRING analysis revealed a significant number of protein-protein interactions, with thirteen nodes and seven association edges (PPI enrichment p-value = 7.51×10–7) (Fig. 5), implicating these proteins in biologically related pathways. Of note, coexpression occurred between ApoA-I and ApoB-100, and each of these apolipoproteins was independently linked to coexpression with SgII and Alpha-1-M (Fig. 5). CPN2 was linked to coexpression with CD5L and Serpin A4 (Fig. 5).
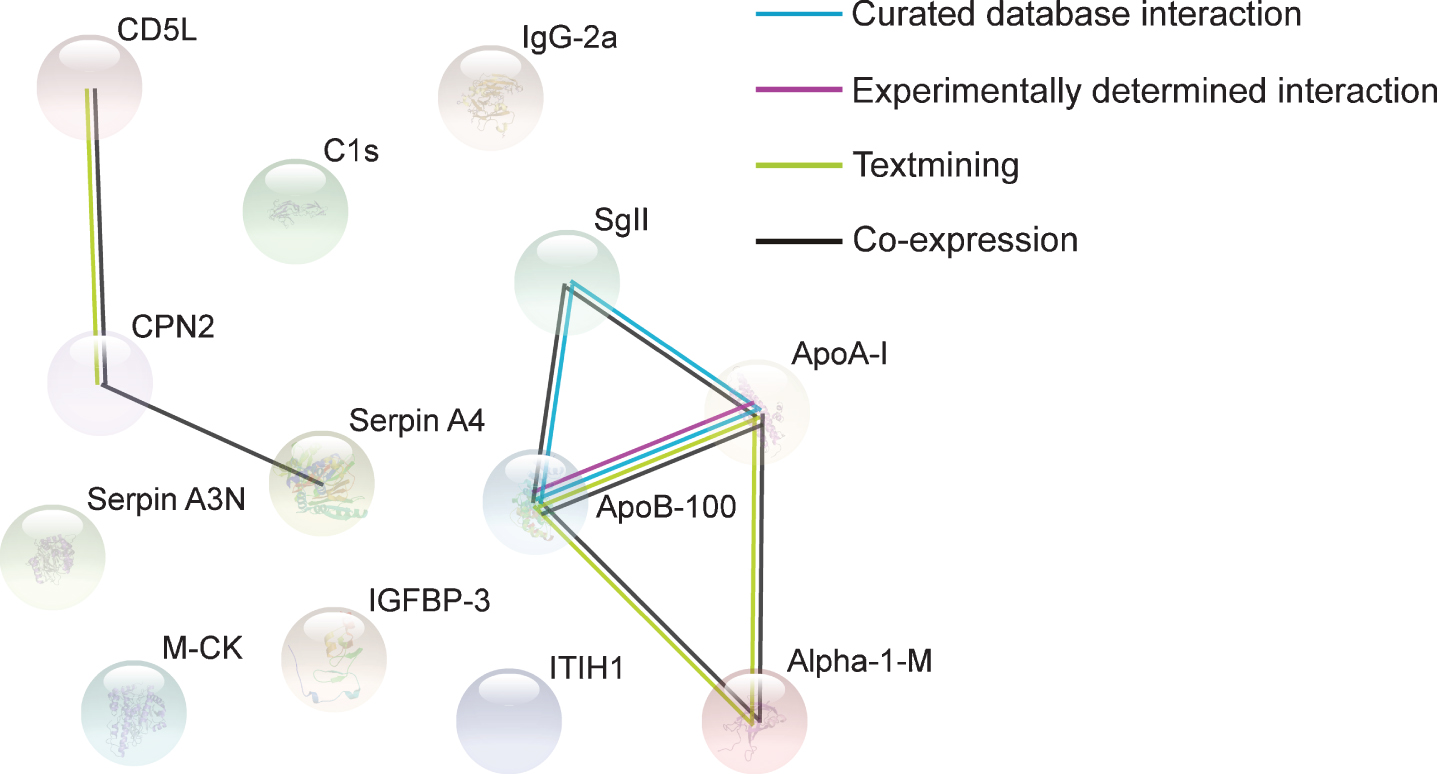
Protein associations in the cerebrospinal fluid (CSF). Proteins presented are those whose abundance was altered (upregulated or downregulated) by M. vaccae NCTC 11659 treatment, as assessed seven days following the final immunization. Associations were computed by STRING database (v.11). Final sample sizes for CSF analysis were vehicle, n = 5, M. vaccae NCTC 11659, n = 6. Alpha-1-M, alpha-1-macroglobulin; ApoA-I, apolipoprotein A-I; ApoB-100, apolipoprotein B-100; C1s, complement C1s subcomponent; CD5L, CD5 antigen-like; M-CK, creatine kinase, M-type; CPN2, carboxypeptidase N subunit 2; IgG-2a, Ig gamma-2A chain C region; IGFBP-3, insulin-like growth factor-binding protein 3; ITIH1, inter-alpha trypsin inhibitor, heavy chain 1; SgII, secretogranin-2; Serpin A3N, serine protease inhibitor A3N; Serpin A4, serine proteinase inhibitor, clade A, member 4.
In order to better understand how M. vaccae might be altering different biological functions, functional enrichment gene analyses, including pathway analysis and gene ontology (GO) analyses, were performed separately on subsets of proteins that were upregulated by immunization with M. vaccae (N = 5: Serpin A3N, C1s, IGFBP-3, IgG-2a, and SgII) and those that were downregulated by immunization with M. vaccae (N = 8: Alpha-1-M, ApoA-I, ApoB-100, CD5L, CPN2, ITIH1, M-CK, Serpin A4) using DAVID bioinformatics resource.
There were no pathways enriched by the set of upregulated proteins. Functional enrichment analysis for GO, biological process, revealed the top two hits to be “complement activation, classical pathway” and “innate immune response” (Fig. 6, Supplementary Table 4). Functional enrichment analysis for GO, cellular component, indicated the top three hits to be “extracellular region,” “blood microparticle,” and “extracellular space” (Fig. 6, Supplementary Table 4). There were no results for the functional enrichment analysis for GO, molecular function, within the subset of proteins upregulated by M. vaccae.
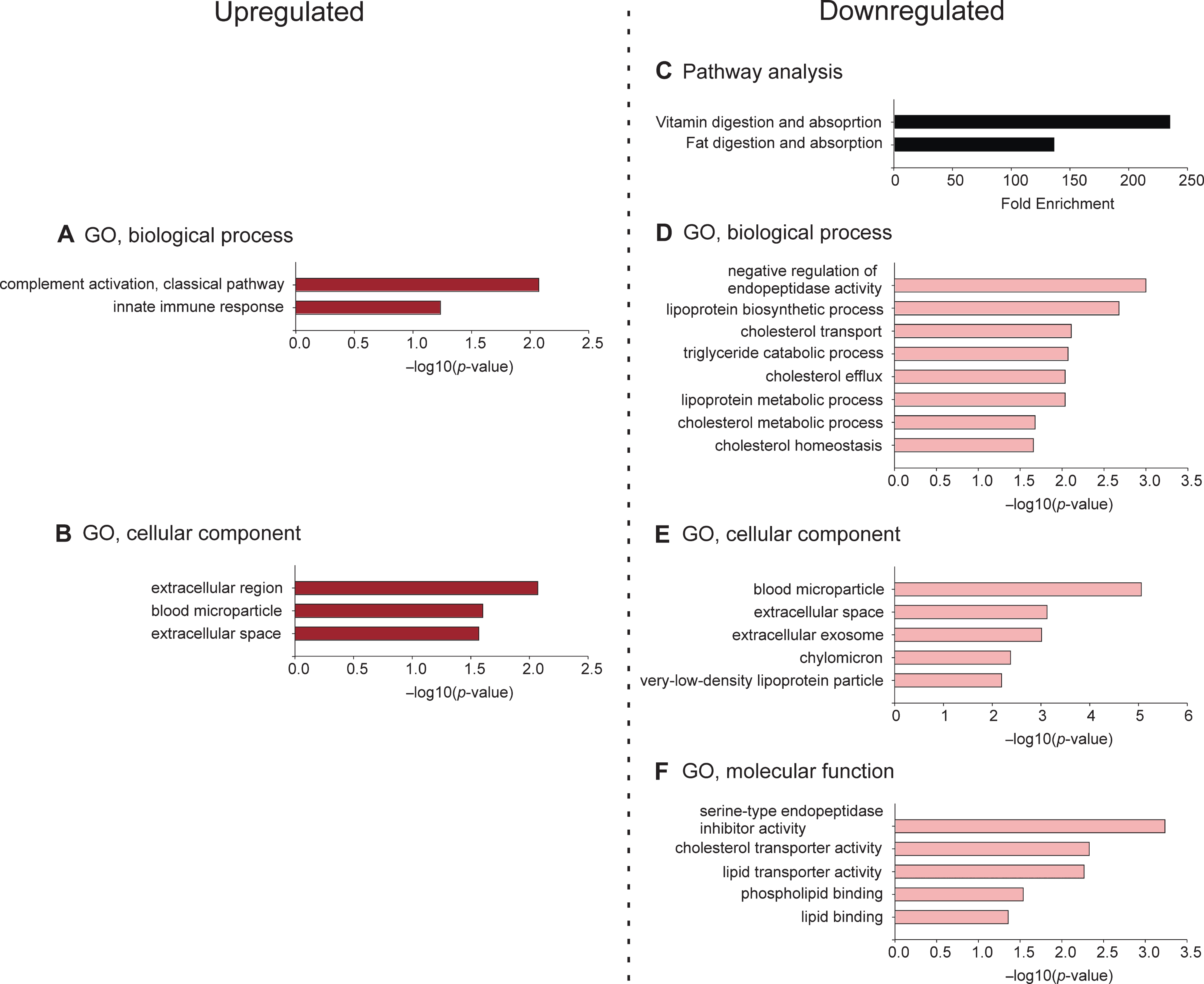
Functional enrichment analyses, including pathway analysis and gene ontology (GO) analyses, for proteins altered in the cerebrospinal fluid (CSF) by immunization with M. vaccae NCTC 11659. Gene enrichment analyses included (A) biological processes for upregulated proteins, (B) cellular components for upregulated proteins, (C) enriched pathways for downregulated proteins, (D) biological processes for downregulated proteins, (E) cellular components for downregulated proteins, and (F) molecular functions for downregulated proteins. Analyses were completed using DAVID bioinformatics resource. Values for GO analyses represent –log10(p-value) for enrichment of gene set contributing to each pathway, biological process, cellular component, and molecular function using a modified Fisher’s exact test (EASE) score; values for pathway analysis represent fold enrichment of the gene set relative to the rat genome. GO, gene ontology.
Among downregulated proteins, the top two pathways were “vitamin digestion and absorption” and “fat digestion and absorption” (Fig. 6, Supplementary Table 4). Functional enrichment for GO, biological process, revealed the top three hits, as ordered by p-value, to be “negative regulation of endopeptidase activity,” “lipoprotein biosynthetic process,” and “cholesterol transport” (Fig. 6, Supplementary Table 4). Functional enrichment for GO, cellular component, indicated the top three hits, as ordered by p-value, to be “blood microparticle,” “extracellular space,” and “extracellular exosome” (Fig. 6, Supplementary Table 4). Functional enrichment for GO, molecular function, indicated the top three hits, as ordered by p-value, to be “serine-type endopeptidase inhibitor activity,” “cholesterol transporter activity,” and “lipid transporter activity” (Fig. 6, Supplementary Table 4).
Hippocampal mRNA expression
It has been reported that M. vaccae treatment specifically increases Il4 mRNA expression, IL-4 protein expression, and IL-4-responsive gene expression in the rat hippocampus [5, 31]. Thus, in parallel with our proteomics findings, we investigated the expression of several immunoregulatory markers in the hippocampus. Here, we confirmed the previous results, showing that immunization with M. vaccae increased hippocampal Il4 mRNA expression (p < 0.05) but did not affect Il10 or transforming growth factor beta 1 (Tgfb1) mRNA expression in the hippocampus (Fig. 7). An effect of M. vaccae treatment on Il10 mRNA expression in the hippocampus approached statistical significance (p = 0.05) (Fig. 7). In this study, immunization with M. vaccae did not alter mRNA expression of IL-4-responsive genes, including Cd200, Cd200r1, and Mrc1 (Supplementary Table 5).
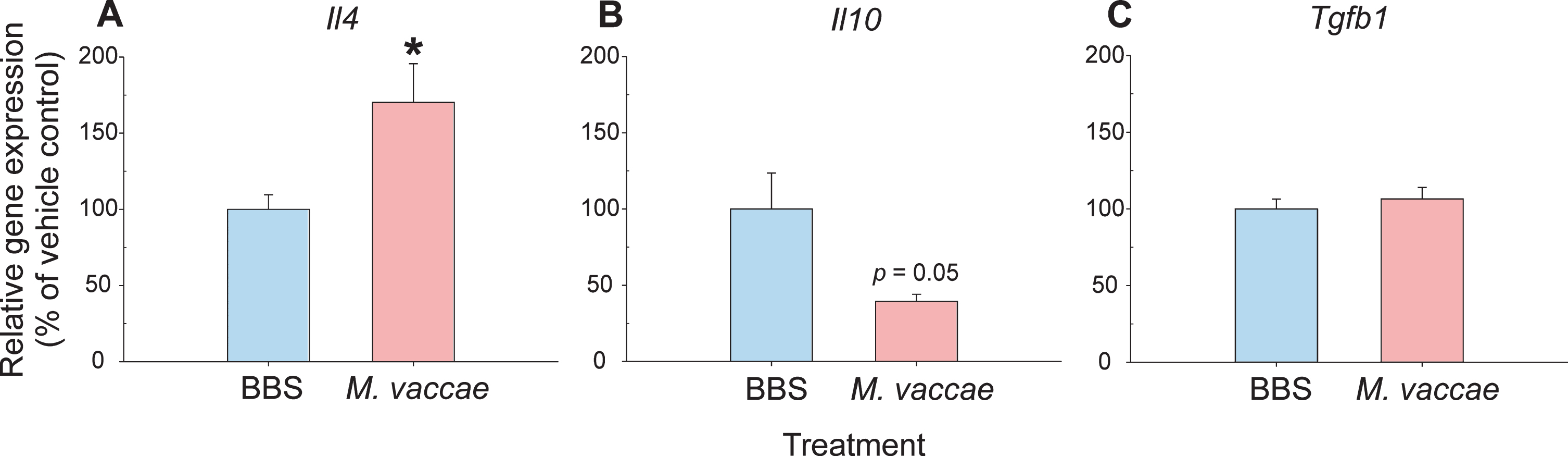
Effects of a heat-killed preparation of Mycobacterium vaccae NCTC 11659 (M. vaccae), administered subcutaneously once per week for three weeks, on anti-inflammatory cytokine mRNA expression in the rat hippocampus measured seven days following the final immunization. Expression of A) interleukin (Il)4, B) Il10, and C) transforming growth factor beta 1 (Tgfb1) mRNA in the hippocampus of M. vaccae-treated and borate-buffered saline (BBS) vehicle-injected control rats. Measurements were made using RT-qPCR and the 2–ΔΔCt method, with Actb, encoding β-actin, as a reference. Bars represent the mean + SEM. Sample sizes: Il4 (BBS, n = 8; M. vaccae, n = 8); Il10 (BBS, n = 6; M. vaccae, n = 6); and Tgfb1 (BBS, n = 8; M. vaccae, n = 8). *p < 0.05. BBS, borate-buffered saline; Il, interleukin; M. vaccae, Mycobacterium vaccae NCTC 11659; Tgfb1, transforming growth factor-β1.
DISCUSSION
Immunization with M. vaccae NCTC 11659 altered the relative abundances of specific proteins in plasma and CSF, assessed one week following the final immunization. Analysis of the plasma proteome revealed effects of immunization with M. vaccae on three proteins: ApoC-II, IgG-2a, and Serpin A4. Analysis of the CSF proteome revealed effects of immunization with M. vaccae on 13 proteins. We observed increases in IgG-2a in plasma and IgG-2a and C1s in CSF, which may be useful biomarkers of immune activation following immunization with M. vaccae. Further, several of the proteins altered in the CSF are involved in peripheral lipid metabolism and immune-modulating functions, which may be primary biological features induced by M. vaccae. Two proteins that were altered by immunization with M. vaccae were altered in both plasma and CSF, IgG-2a, and Serpin A4. This, together with the finding that hippocampal expression of mRNA encoding a subset of seven of the proteins was not altered following immunization with M. vaccae (Supplementary Table 5, for primer details see Supplementary Table 1), suggests that altered protein levels in the CSF may be due to: 1) altered filtration of blood by the choroid plexus; 2) altered translocation of proteins across the blood-brain barrier (BBB); or 3) altered clearance from the CSF, rather than altered expression in brain parenchyma. However, immunization with M. vaccae did increase expression of hippocampal Il4 mRNA, consistent with previous studies and consistent with development of an anti-inflammatory immunophenotype in the brain.
Analysis of the plasma proteome revealed effects of immunization with M. vaccae on three proteins. In the plasma, IgG-2a and Serpin A4 were upregulated, whereas ApoC-II was downregulated, by immunization with M. vaccae. Previous studies of immunization with heat-inactivated mycobacteria, including the M. vaccae strains or their isolated protein derivatives, described increases in IgG-2a in serum of both mice and rats [88–92] that increase 14, 21, and 28 days after injection [90] and prime the immune system for subsequent challenge [91]. This timeline is consistent with our studies, with measurement of plasma proteins 21 days following the first immunization. Likewise, mycobacteria alter the abundance of Serpin A4 and ApoC-II in human plasma [93]. While our plasma dataset was not enriched for any pathways annotated in DAVID, it is important to point out that IgG-2a and Serpin A4 are involved in immune pathways, whereas ApoC-II is involved in lipid homeostasis, paralleling results from our CSF data, below (see Table 1). It is also worth noting that among the top 21 proteins of interest in the plasma (i.e., top 15% of all proteins measured in the plasma based on p-value; Supplementary Table 2), five (24%) of these overlap with the top proteins of interest in the CSF (i.e., top 15% of all proteins measured in the CSF based on p-value; Supplementary Table 3) and fold change analysis results in the CSF. Thus, the implications for plasma proteins may be more meaningful when combined with CSF data.
Proteins altered by immunization with M. vaccae NCTC 11659 in plasma and cerebrospinal fluid (CSF) and their primary functions
1All protein and gene names were identified by UniProt database (https://www.uniprot.org/). 2All functions were obtained from NCBI Gene (https://www.ncbi.nlm.nih.gov/gene/) unless otherwise indicated. *indicates proteins that were altered in both plasma and CSF.
Analysis of the CSF proteome revealed effects of immunization with M. vaccae on 13 proteins. Previous studies have shown that infection with mycobacteria can alter the abundance of several of these proteins (ApoA-I, Alpha-1-M, CD5L, IGFBP-3, Serpin A3, Serpin A4) in the plasma or serum of different animal models, including rodents, cattle, and primates, as well as in humans [93–97]. In the present study, our protein set upregulated by M. vaccae was associated with immune activation, whereas the proteins downregulated by M. vaccae were highly enriched for lipid homeostasis. In this case, both high-density lipoprotein (HDL) and low-density lipoprotein (LDL)-associated proteins were downregulated by M. vaccae. ApoA-I constitutes an HDL-binding protein, whereas ApoB-100 constitutes an LDL-binding protein (NCBI RefSeq NP_036870.1, RefSeq NP_062160.2; Table 1). Of note, the functional enrichment for gene ontology, cellular component, indicated “very-low-density lipoprotein particle” to be downregulated by M. vaccae treatment (Fig. 6, Supplementary Table 4). Importantly, CPN2, ITIH1, C1s, CD5L, Serpin A4, and IgG-2a may be HDL-associated proteins, whereas Alpha-1-M and IGFBP-3 may be LDL-associated proteins, despite not being annotated as such by DAVID [98–103]. HDL has been shown to bind directly to Mycobacterium avium. This binding is thought be dependent on binding of ApoA-I to lipids in the outer cell wall of M. avium and may be one mechanism through which M. avium escapes from host innate immunity [104]. Although it is not known if HDL binds to M. vaccae NCTC 11659, direct interaction between M. vaccae NCTC 11659 and HDL is one mechanism through which M. vaccae may alter plasma and CSF concentrations of HDL-associated proteins, particularly as HDL appears to traverse the BBB [105].
We observed increases in IgG-2a in plasma and IgG-2a and C1s in CSF, which may be biomarkers of immune activation following immunization with M. vaccae and potentially other mycobacteria. Increases in these immunological markers may be paired because IgG-2a is a potent activator of the classical complement system in rodents, including rats [106, 107]. Indeed, proteins upregulated by M. vaccae in the CSF are enriched for the biological process “complement activation, classical pathway” (Fig. 6, Supplementary Table 4). Heat-killed preparations of M. vaccae are not pathogenic; on the contrary, previous studies have shown that immunizations with M. vaccae induce long-term immunoregulatory phenotypes in the periphery and the CNS of rodents [5, 31]. In the current study, plasma and CSF were not taken until 21 days after the first immunization (one week after the final immunization), and the increased abundance of these immune pathways may point to long lasting, complex immunological changes biased toward promotion of stress resilience effects of M. vaccae. It is important to mention that physiological immune responses can initiate simultaneous proinflammatory and anti-inflammatory events and that classically characterized cytokines can have pleiotropic effects, i.e., both proinflammatory and anti-inflammatory effects, depending on context [108, 109]; further, measurements of immune markers are dependent on the time course, immunological context, and tissue of the immune activation and resolution [110, 111]. Additionally, components of classical complement activation, namely C1q, are involved in immunoregulatory processes that induce M2 macrophage polarization and tissue repair [109, 112]. In fact, breakdown of apolipoproteins can lead to activation of complement, such that increases in innate immune signaling proteins may be a response to the altered lipid metabolism/decreased apolipoprotein abundance in plasma and CSF [113]. Interestingly, changes in lipid metabolism (also enriched in our protein set) are thought to be intimately connected to innate and adaptive immune responses as well as resolution of the inflammatory process through resolvins and lipoxins [110, 115]. It is currently not known if M. vaccae alters resolvin or lipoxinsignaling.
Several of the proteins altered in the CSF following immunization with M. vaccae are involved in both peripheral lipid metabolism and immune-modulating functions, which may be primary biological features induced by M. vaccae. Of the proteins upregulated by M. vaccae, Serpin A3N and IGFBP-3 may be of importance. Orthologs of Serpin A3N are upregulated following M. tuberculosis infection in mice and humans [62, 96]. Serpina3n encodes the serine protease inhibitor A3N, also known as alpha-1-antichymotrypsin. As part of the serpin superfamily, Serpin A3 is well known for its involvement in the acute inflammatory process [116], but it has also been implicated in fatty acid metabolism in association with peroxisome proliferator-activated receptor (PPAR) signaling [117, 118]. IGFBP-3 is decreased in the serum of humans infected with M. tuberculosis, contrary to our results in the CSF [93]. IGFBP-3 is thought to be involved in several signaling pathways, including apoptosis, cellular metabolism, and inflammation; it mediates glucose and cholesterol metabolism by binding insulin-like growth factor 1 (IGF-1) and 2 (IGF-2), but it may also exert anti-inflammatory and metabolic functions independent of IGF-1 and IGF-2 [119]. IGFBP-3 is highly expressed in immunoregulatory M2 macrophages, and can prevent cytokine-induced insulin resistance [120]; thus the increase of IGFBP-3 in CSF after M. vaccae treatment may be associated with a number of physiological responses, including lipid homeostasis and immunoregulation, and its role should be further investigated. Of the proteins downregulated by M. vaccae, CD5L may be of particular importance. CD5L has been reported to be increased after M. tuberculosis infection, contrary to our results [97, 121]; however, CD5L is a regulator of cholesterol biosynthesis in immune cells such as macrophages [122] and T helper 17 cells [123], and thus may be downregulated in the context of the altered lipid metabolism described in our functional enrichment analysis.
Studies have shown that mycobacteria can induce metabolite signatures in the host plasma that are associated with the phospholipid structures found in the bacteria’s cell membrane. The mycobacteria may be interacting with the host to alter lipid metabolism through these phospholipid components [63, 124]. Various unsaturated fatty acids, derived from phospholipids, can alter lipoprotein secretion of human intestinal cells [125]. In support of this idea, functional enrichment analysis of downregulated proteins in the CSF included the molecular function “phospholipid binding” (GO:0005543). In addition, among the top 15% of proteins of interest in the plasma and CSF, 62% of proteins in the plasma (13 of 21) and 47% of proteins in the CSF (14 of 30) are associated with lipoproteins, primarily HDL (Supplementary Tables 2 and 3). Interestingly, among the top 15% of proteins of interest (determined by p-value) in the CSF, ectonucleotide pyrophosphatase/phosphodiesterase 2 (E-NPP 2, p = 0.102, fold change = 1.50), a protein that acts as a phospholipase to catalyze the production of lysophosphatidic acid from lysophosophatidylcholine (NCBI RefSeq NP_476445.2), and lipoprotein-associated phospholipase A2 (lp-PLA2, also known as platelet-activating factor acetylhydrolase, or PAF-AH, p = 0.151, fold change = 1.40), a lipoprotein-associated protein that hydrolyzes oxidized phospholipids to produce lysophosphatidylcholine [126], were upregulated, although these results were not significant (Supplementary Table 3). Other members of the phospholipase A2 superfamily produce polyunsaturated fatty acids and eicosanoids from phospholipid membranes, which promote downstream signaling of PPARα, PPARγ, and PPARδ pathways [127]. Indeed, an interesting pattern has emerged from our dataset that may include phospholipid metabolism and inflammatory/resolving pathways.
A recent study identified a novel lipid metabolite of M. vaccae NCTC 11659, 10(Z)-hexadecenoic acid, with anti-inflammatory properties. Interestingly, this free fatty acid specifically mediates PPARα signaling in macrophages and may thereby be able to modulate the immune system through a known regulator of lipid metabolism [23] (KEGG: hsa03320). We have been able to associate a number of the proteins altered by M. vaccae with a more general PPAR signaling pathway. For example, PPARα agonists alter expression of metabolic proteins such as ApoA-I, ApoB-100, and IGFs in the monkey liver [128]. PPARα agonist treatment increases catabolism of apolipoproteins like ApoA-I and ApoB-100 [129], which were both decreased in the CSF in our study. Likewise, expression of SgII, a regulator of immune function, may be altered by treatment with fatty acids, including PPARγ agonists [130, 131]. Serpin A3, which was increased in CSF in our study, alters lipid homeostasis by activating PPARγ signaling [117]. IGFBP-3 can directly interact with the PPARγ pathway to regulate adipocyte differentiation, although other downstream immunoregulatory effects may be possible [132]. CD5L also acts as a regulator of both inflammatory pathways and lipid metabolism. In fact, CD5L secreted from activated macrophages can increase lipolysis and decrease the activity of PPARγ [133]. Since other mycobacteria strains are known to activate the PPARγ pathway [134], it may be possible that various unsaturated, monounsaturated, or polyunsaturated fatty acid components of M. vaccae can cause changes in activation of both PPARα and PPARγ pathways. In light of these findings, we were unable to find altered expression of Ppara (encoding PPARα), Pparg (encoding PPARγ), or Ppard (encoding PPAR δ) in the hippocampus (Supplementary Table 5, for primer details see Supplementary Table 1), which may suggest altered PPAR signaling in the periphery contributes to lasting effects found in the CSF. Supporting this idea, there has been increasing interest in targeting peripheral PPAR signaling for the treatment of psychiatric disorders such as post-traumatic stress disorder [135] and neurological disorders like AD [136] because of the roles that PPARs play in metabolic and inflammatory signaling from the periphery to the CNS; intriguingly, recent studies have shown that the gut microbiome can induce alterations in the PPAR signaling pathways [137, 138]. Further research should be done to investigate the possible altered PPAR signaling in the periphery of animals immunized with M. vaccae.
Two proteins that were altered by immunization with M. vaccae were altered in both plasma and CSF, IgG-2a, and Serpin A4. This, together with the finding that hippocampal expression of mRNAs encoding a subset of seven of the proteins that were differentially expressed in CSF were not altered following immunization with M. vaccae (Supplementary Table 5, for primer details see Supplementary Table 1), suggests that altered protein levels in the CSF may be due to: 1) altered filtration of blood by the choroid plexus; 2) altered translocation of proteins across the BBB; or 3) altered clearance of these proteins from the CSF, rather than altered expression in brain parenchyma. This idea is also supported by the finding that, although PPAR-associated proteins were found to be altered in the CSF, the mRNA expression of PPARs in the hippocampus did not differ between M. vaccae and vehicle-treated animals. This is not to say that protein abundance of any proteins in the hippocampus did not change, nor that mRNA or protein expression are not different in other brain regions, and this should be tested in future experiments. However, proteins altered in the CSF show gene expression patterns that are highest in peripheral tissues such as liver (Apoa1, Apob, Cpn2, C1s, Itih1, Serpina3, Serpina4) and kidney (Igfbp3). Indeed, peripheral bacteria and their metabolites can alter the permeability of the BBB or blood-CSF barrier at the choroid plexus [139–141]. Lipopolysaccharide (LPS) as well as other bacterial components, can increase permeability of the BBB [142, 143], whereas interactions between host and gut bacteria that produce short-chain fatty acids can decrease BBB permeability [140, 145]. The overlap of altered proteome observed in plasma and CSF (particularly considering Serpin A4 was increased in plasma but decreased in CSF) may therefore implicate host responses that are associated with BBB integrity. For example, NF-κB is thought to play a direct role in BBB permeability [146, 147], and the increase in plasma Serpin A4 may act as an antagonist for this pathway, as Serpin A4 is known to decrease NF-κB activation in various tissues [148–150]. In addition, proteins that associate with lipoproteins may be more easily transported across the BBB, and peripheral changes in lipid homeostasis may explain changes in abundance of HDL- and LDL-associated proteins in the CSF [151–153]. Corroborating our findings and supporting peripheral changes in similar clusters of proteins, studies in humans have shown that intermittent fasting increases Serpin A3 and decreases apolipoprotein A-II (ApoA-II), ApoC-II, and ApoB in the plasma of humans, following the pattern seen in the plasma and CSF of M. vaccae-immunized rats in our study [154]. This provides evidence that peripheral changes in abundance of these proteins may frequently occur together.
Immunization with M. vaccae increased expression of hippocampal Il4 mRNA, consistent with previous studies and consistent with development of an anti-inflammatory immunophenotype in the brain. We have previously shown an anti-inflammatory effect of M. vaccae associated with stress-resilient behavior and prevention of cognitive impairment in a rat model of post-operative cognitive dysfunction [5, 31]. For example, immunization with M. vaccae can induce an immunoregulatory phenotype in the hippocampus of unstressed animals, including increases in mRNA expression of Il4, Cd200r1, and Mrc1 (i.e., CD206, mannose receptor C-type 1), an IL-4 responsive gene and a marker of M2 macrophages [31]. The increased expression of hippocampal Il4 mRNA found in our study is intriguing because it implies altered immunological signaling in the brain, although the mechanisms underlying this anti-inflammatory immunophenotype are unknown. It could be possible that IL-4 signaling is a response of alternatively activated macrophages to the altered lipid and immune signaling in the CSF [155]. Likewise, IL-4 signaling could be driven by T cells, possibly stimulated by peripheral dendritic cells, residing in the choroid plexus [156, 157]. Interestingly, cathepsin S (CatS), a marker of antigen-presenting cells, may be upregulated by 70% in the CSF, although this effect was not significant (p = 0.11, fold change = 1.71).
Limitations
Although our study constitutes a positive first step to understanding effects of mycobacteria on lipid-immune pathways in the plasma and CSF, our proteomics results are limited by our small sample sizes (for plasma proteomics, vehicle, n = 7, M. vaccae, n = 8; for CSF proteomics, vehicle, n = 6, M. vaccae, n = 5). Although repeated immunizations withM. vaccae alter proteomic and hippocampal gene expression, our data do not establish a mechanism of action. Our data are also limited by the single time point of tissue collection; future studies should evaluate the effects of immunization with M. vaccae at multiple time points. Considering the relatively few immune markers associated with M. vaccae treatment, we may be witnessing some time point within the resolution phase of an immune response, or it may be that these immune-related proteins are associated with alternative signaling pathways, not necessarily tied to inflammation. Furthermore, this experiment was conducted only in male rats; future studies should include female rats to determine if there are sex differences. Likewise, results may be different in a population of aged animals immunized with M. vaccae, due to age-related changes in immune and metabolic function. Future studies may also benefit from including an additional stressor component, to understand how these biological markers may associate with behavioral outcomes.
Clinical implications and future directions
Our study demonstrates that immunization with M. vaccae alters proteins in the plasma and CSF associated with immune signaling and lipid homeostasis, providing us with a more holistic perspective of M. vaccae’s direct effects on the physiology of undisturbed, unstressed animals. These data complement previous studies showing effects of M. vaccae on cytokine and immunoregulatory profiles [5, 31], neurotransmitter and hormonal changes in the brain [27–29, 72], and stress-resilient behavior [5, 31]. An important next step will be to explore effects of M. vaccae on the proteome in stressed animals, and the potential role of these proteomic changes in the stress resilience (including resilience to stress-induced cognitive impairment) properties of M. vaccae. Our current data support the hypothesis that M. vaccae works by activating anti-inflammatory and immunoregulatory immune pathways that can protect against inflammation-potentiated stress-induced behaviors [5, 26]; however, in line with previous studies [30], we now have evidence to suggest that altered lipid metabolism is also at play, potentially due to bioactive components, including fatty acid components, of the bacteria.
Considering that M. vaccae increases anti-infla-mmatory and immunoregulatory immune signaling and alters lipid homeostasis in the periphery and the brain, M. vaccae treatments may be relevant in the context of diseases that are marked by a dysfunctional cellular metabolic component coupled with inflammation, such as AD [158]. Intriguingly, previous studies have shown that immunization withM. vaccae can protect aged rats from cognitive decline after surgery, prevent surgery-induced expression of IL-1β protein, and increase Il4 mRNA in the hippocampus in a model of post-operative cognitive dysfunction in aged rats [5]. Our study has shown that M. vaccae decreases LDL- and HDL-associated apolipoproteins in plasma and CSF and has confirmed previous studies demonstrating increases in Il4 mRNA in adult male rats [5, 31]. Dysregulated lipid (including cholesterol, phospholipid, and lysophospholipid) and glucose metabolism is a key feature of AD [159, 160], and new studies suggest dysregulated abundance of ApoA-I and ApoB are associated with increased risk of AD development [161, 162]; in our study, we found that M. vaccae decreased ApoA-I and ApoB-100 in CSF. Phospholipase A2 (PLA2), which cleaves phospholipids to form lysophospholipids, may be beneficial for the treatment of AD, in combination with amyloid-β (Aβ)-targeted vaccines [163]; in our study, we found trending increases in lp-PLA2 and corresponding E-NPP 2 in the cerebrospinal fluid of M. vaccae-treated rats. In addition, injections of virus expressing IL-4 into the hippocampus of AD mouse models prior to the onset of the disease prevent accumulation of Aβ plaques and glial cell activation in the hippocampus during pathogenesis of the disease, and further, IL-4 injections prevent spatial-learning deficits in these mice [164]. Likewise, IL-4 alleviates Aβ-induced neuroinflammation and inhibition of long-term potentiation in rats [165]. Therefore, M. vaccae may provide effects in young animals that are protective against dementia pathology later in life, in part by increasing IL-4 expression in the hippocampus. Preventative versus treatment effects of M. vaccae will be an important consideration of future experiments, in particular because immune function, including M. vaccae-induced changes in immune signaling, is altered across the lifespan [5, 166].
In the context of AD, it is important to have an alternative activation of the immune system that promotes clearance of aggregated proteins but prevents chronic inflammatory states [167]. IL-4 secretion is indicative of an immunoregulatory phenotype and a shift toward M2 macrophages that promote healing and tissue repair [168], as is PPAR pathway activation [169, 170]. In addition, immunoglobulins like IgG-2a might be important to initiate macrophage phagocytosis [171], and clearance of aggregated proteins such as Aβ in AD [172]. Toll-like receptor 2 (TLR2) signaling, which is also activated by M. vaccae [173], in microglia may offer neuroprotection against oligomeric Aβ [174]. Exposure to bacteria and other “old friends” may promote this type of Th1/Th2 balanced immune function [14, 15]. Indeed, the microbiome is becoming recognized for its potential to affect vulnerability and/or resilience in the development of AD [34]. For example, LPS may indirectly enhance the translocation of Aβ proteins across the BBB in AD [175]. On the contrary, persons with AD have increased permeability at the BBB and choroid plexus, suggesting that probiotics promoting BBB integrity may be beneficial to persons with AD [176, 177]. In fact, probiotics such as Bifidobacterium breve A1 prevent Aβ-induced memory impairments in mice [178]. Considering M. vaccae’s inflammation-buffering and stress-resilience effects, particularly in aged animals, immunization with M. vaccae in the context of AD may be an important direction for future research.
Potential relevance to the etiology, pathophysiology, and treatment of AD
Excitement is building for the potential of the microbiome to treat disease, and there is a recent momentum to study microbiome-host interactions in the context of AD. Our data provide evidence that treatment with immunobiotics, such as M. vaccae NCTC 11659, may influence metabolic signaling, altering the abundance of AD-associated proteins, in the host. Several lines of research, combined with these current data, suggest that immunization with M. vaccae might be an effective therapy for the prevention or treatment of AD-like pathology. Although the current study does not provide behavioral data, prior studies have described enhanced cognition after treatment with M. vaccae in rodents, including improvements in learning and memory in adult mice [179] and the prevention of cognitive deficiencies after surgery in aged rats, associated with anti-inflammatory signaling in the hippocampus [5]. M. vaccae may also be attractive for AD research because of its unique ability to act as an adjuvant to the immune system [8, 180] by activating TLR2 signaling in dendritic cells [173, 174], and by conferring protective immunoregulatory processes in the periphery and the brain [5, 31]. Considering that the immune-stimulating properties of M. vaccae strains are comparable to those of BCG in TB studies [91, 181] and that BCG administration may help prevent and treat AD [2, 3], M. vaccae is a compelling novel therapeutic candidate for AD pathologies. Collectively, these data argue for future research of M. vaccae immunizations in AD rodent models as well as models of other neurodegenerative disorders, including Parkinson’s disease, age-related cognitive decline, dementia, and Huntington’s disease [182].
Immunization with M. vaccae NCTC 11659, pathobionts, and pathogens
Although we have presented our data in the context that immunization with M. vaccae NCTC 11659 is inducing anti-inflammatory and immunoregulatory effects, and that these effects mediate the stress resilience effects of M. vaccae NCTC 11659, we cannot exclude the possibility that the beneficial effects of M. vaccae NCTC 11659 may be due to effects on immunologic responses to pathogens or “pathobionts”, either in our animal studies, or, particularly, in humans. For example, we have shown that chronic psychosocial stress in mice results in a three order of magnitude increase in the relative abundance of Helicobacter spp., which is highly correlated with histological damage to the colon [26]. In subsequent studies, we showed that the stress-induced histological damage to the colon is absent when mice are housed in a specific-pathogen-free facility, and that stress-induced histological damage to the colon can be reinstated following inoculation of mice with Helicobacter typhlonius [183]. In this case, immunization with M. vaccae NCTC 11659 did not prevent stress-induced proliferation of Helicobacter spp., but instead appeared to induce an immune tolerance of Helicobacter spp., thus preventing the stress-induced inflammation and pathophysiology [26, 183]. Indeed, induction of immune tolerance to pathogens is gaining traction as a potential strategy for dealing with some pathogens, in essence converting pathogens or pathobionts into harmless symbionts [184–189]. In addition to disease tolerance, immunization with M. vaccae NCTC 11659 could promote beneficial outcomes by reducing pathogen load; for example, evidence suggests that patients with infection with M. tuberculosis have a higher risk of dementia [190] (see also evidence for increased bacteria in AD postmortem brain [191], and review by Mawanda and Wallace [36]). This hypothesis is supported by a recent Phase III clinical trial using M. vaccae NCTC 11659 in patients with M. tuberculosis infection, demonstrating more rapid clearance of mycobacteria [8].
Footnotes
ACKNOWLEDGMENTS
These studies were funded by the National Institute of Health (NIH) R21 grant awarded to C.A.L., S.F.M., M.G.F. (PIs), and L.K.F. (Co-I). We gratefully acknowledge Jared D. Heinze and James E. Hassell, Jr. for conducting M. vaccae and borate-buffered saline injections, and Yongkang Kim for consulting and advice related to data analysis. Purchase of a Thermo Orbitrap Velos mass spectrometer was made possible with a grant from W.M. Keck Foundation.