
Abstract
Background:
Alzheimer’s disease (AD) is a neurodegenerative disease characterized by progressive decline in cognitive function. Type 2 diabetes mellitus (T2DM) is an important risk factor for AD. Glucose-dependent insulinotropic polypeptide (GIP) has been identified to be effective in T2DM treatment and neuroprotection.
Objective:
The present study investigated the neuroprotective effects and possible mechanisms of DAla2GIP-Glu-PAL, a novel long-lasting GIP analogue, in APP/PS1 AD mice.
Methods:
Multiple behavioral tests were performed to examine the cognitive function of mice. In vivo hippocampus late-phase long-term potentiation (L-LTP) was recorded to reflect synaptic plasticity. Immunohistochemistry and immunofluorescence were used to examine the Aβ plaques and neuroinflammation in the brain. IL-1β, TNF-α, and cAMP/PKA/CREB signal molecules were also detected by ELISA or western blotting.
Results:
DAla2GIP-Glu-PAL increased recognition index (RI) of APP/PS1 mice in novel object recognition test, elevated spontaneous alternation percentage of APP/PS1 mice in Y maze test, and increased target quadrant swimming time of APP/PS1 mice in Morris water maze test. DAla2GIP-Glu-PAL treatment enhanced in vivo L-LTP of APP/PS1 mice. DAla2GIP-Glu-PAL significantly reduced Aβ deposition, inhibited astrocyte and microglia proliferation, and weakened IL-1β and TNF-α secretion. DAla2GIP-Glu-PAL also upregulated cAMP/PKA/CREB signal transduction and inhibited NF-κB activation in the hippocampus of APP/PS1 mice.
Conclusion:
DAla2GIP-Glu-PAL can improve cognitive behavior, synaptic plasticity, and central pathological damage in APP/PS1 mice, which might be associated with the inhibition of neuroinflammation, as well as upregulation of cAMP-/PKA/CREB signaling pathway. This study suggests a potential benefit of DAla2GIP-Glu-PAL in the treatment of AD.
INTRODUCTION
Alzheimer’s disease (AD) is a progressive neurodegenerative disease in the central nervous system. According to World Alzheimer’s Disease Report 2018, dementia affects more than 47 million people worldwide, and this number will increase to 152 million by 2050 [1]. However, there is no effective drug to stop the AD process up to now. It is well known that the main pathological hallmarks in the AD brain include high-density of amyloid-β (Aβ) plaques and neurofibrillary tangles, as well as progressive neuronal loss and brain atrophy [2]. Aβ deposition in the brain may be the basis for the pathological changes in AD, which triggers tau protein hyperphosphorylation and ultimate neuronal degeneration. It is reported that intracerebral ventricular [3] or intra-hippocampal [4] injection of Aβ suppressed long-term potentiation (LTP) of synaptic transmission in the hippocampus CA1 region of rat; Aβ damaged spatial learning and memory in rats [5]; and Aβ evoked neuroinflammation in the hippocampus [6, 7]. In turn, inflammatory factors (such as interleukin 1 beta (IL-1β) and TNF-α) accelerated Aβ deposition [8, 9]. Therefore, it is a promising strategy for the treatment of AD by pre-venting Aβ accumulation and neuroinflammatory response.
Type 2 diabetes mellitus (T2DM) is an important risk factor for AD [10, 11]. Several studies have shown close association between abnormal glucose or insulin metabolism and AD development [12, 13]. For example, Talbot et al. [14] confirmed that insulin signaling was severely desensitized in the brain of AD patients, even in non-diabetic patients. Insulin is a key growth factor which not only regulates energy meta-bolism but also affects structural and functional integrity of synapses in the central nervous system [15]. A clinical study demonstrated that long-acting intranasal insulin improves cognition for adults with mild cognitive impairment or early-stage AD dementia [16]. In view of the similarities and close assoc-iation between AD and T2DM, drugs originally developed for T2DM treatment have been tested as potential treatments for AD. However, the desensitization of the insulin receptor, one of the underlying mechanisms of neurodegeneration in AD [17], made it neither advisable nor helpful to give insulin to people with AD. Moreover, chronic administration of insulin increased insulin desensitization and had dangerous effects on blood sugar levels in AD patients [16]. Glucose-dependent insulinotropic polypeptide (GIP), a 42-amino acid incretin growth factor, is a potential candidate in the treatment of AD as it acti-vates a parallel signaling pathway to insulin, namely, the incretin signaling [18]. GIP mainly modulates the insulin-signaling pathway with insulin-like meta-bolic hormones, rather than directly alleviating insulin resistance by facilitating insulin receptor exp-ression. In the meantime, the hypoglycemic effect of GIP is glucose-dependent [19]. So, it will be safe for AD patients with normal blood glucose level. Moreover, GIP receptors are expressed in the brain esp-ecially on larger neurons such as the cerebral cortex and hippocampal pyramidal neurons [20]. More im-portantly, GIP has been justified to be neuroprotec-tive and regenerative [21]. For example, GIP prevents the detrimental effects of Aβ on synaptic plasticity [22] and spatial learning and memory [23]; GIP also promotes axonal regeneration after sciatic nerve injury [20]. However, endogenous GIP can be qui-ckly degraded by the enzyme dipeptidyl peptidase IV (DPP-IV), with only 2 minutes of biological half-life [24]. Several enzyme-resistant super-GIP molecules have been designed, such as DAla2GIP [18], which has a significantly extended half-life of several hours. In our previous experiments, we have shown that the GIP analogue could cross the blood-brain barrier [25, 26]. DAla2-GIP-Glu-PAL is a structure-modified peptide of DAla2GIP by the addition of a C-16 fatty acid to Lys37 via a glutamate linker. So, DAla2-GIP-Glu-PAL has higher resistance to DPP-IV cleavage [27] and stronger lipid solubility [28] than DAla2GIP. It has been reported that intraperitoneal (i.p.) injection of DAla2-GIP-Glu-PAL showed markedly improved antidiabetic potential by enhanced glucose-lowering and prominent insulinotropic actions [27]. In particular, it has been recently reported that i.p. injection of DAla2-GIP-Glu-PAL showed neuroprotective roles in a chronic Parkinson’s disease mouse model [29]. How-ever, it remains to be investigated whether DAla2-GIP-Glu-PAL also has a neuroprotective effect on AD mouse and what is the possible protective mechanism of DAla2-GIP-Glu-PAL. Holscher et al. [18, 31] have tested the effects of DAla2-GIP on AD model mice in previous preclinical work. But these studies mainly observed the effects of DAla2-GIP, not DAla2-GIP-Glu-PAL, on cognitive behavior and pathological characteristics in AD mice, without in-volving detailed mechanism. Therefore, it will be interesting to test whether this GIP analogue has neuroprotective properties in AD animal model and to clarify its underlying mechanisms.
In the present study, we used DAla2-GIP-Glu-PAL, not DAla2-GIP, and investigated its effects on multiple cognitive behaviors in 9-month-old APP/PS1 tr-ansgenic mouse model of AD. More importantly, the present study clarified the electrophysiological and molecular mechanisms of DAla2-GIP-Glu-PAL reversing cognitive impairment by recording in vivo hippocampus late phase LTP and measuring cAM-P/PKA/CREB signal pathway.
MATERIALS AND METHODS
Animals and treatments
Heterozygous 9-month-old male APPswe/PS1dE9 (APP/PS1) mice with a C57BL/6J background and wild-type (WT) littermates were purchased from the Institute of Laboratory Animal Sciences (SCXX: 2013 -0002) and bred at the Research Animal Center of Shanxi Medical University with approval of the Shanxi Committee on Ethics of Animal Research. Animals were kept in a room with controlled temperature (20–24°C), humidity (60% –80%), light-dark cycle (12/12 h), and free access to standard rodent chow and water. The APP/PS1 and WT mice were ra-ndomly divided into four groups (n = 12 in each group): WT+PBS, APP/PS1+PBS, WT+DAla2GIP-Glu-PAL, and APP/PS1+DAla2GIP-Glu-PAL.
Peptides and drug treatment
DAla2GIP-Glu-PAL was presented by Professor Christian Holscher of Lancaster University, UK. The molecular weight of DAla2GIP-Glu-PAL is 5351.19, with a purity of 98%. The powdery DAla2GIP-Glu-PAL was dissolved in 0.01 M PBS solution at a final concentration of 5μM and stored in a refrigerator at –20°C after dispensing. APP/PS1 and WT mice were injected (i.p.) with DAla2GIP-Glu-PAL (25 nmol/kg) or PBS (0.01 M) at a volume of 0.2 ml once a day for three weeks prior to behavioral study, maintaining the injection during all the behavioral tests.
Novel object recognition test in open field
Novel object recognition test (NORT) was performed in an open field. The apparatus consisted of an open field arena (length, 55 cm; width, 55 cm; hei-ght, 30 cm). Each mouse was placed in the middle of the open-field and was allowed to freely explore for 5 min. All behaviors of mice were recorded by an infrared camera fixed 2 m above the center point of the open field. The camera was connected to a video recorder, monitor, and a computer. The movement traces and running distance of the animals in the open field were analyzed by Ethovision 3.0 software system (Noldus Information Technology, Wageningen, the Netherlands). In the NORT, mice were first subjected to a 10 min acquisition trial, during which they were placed in the open field in the presence of two identical objects (cubes) situated at 15 cm from the arena wall (acquisition task). On completion of 10 min exploration, the mouse was returned to its cage for a 3 h delay. Then, the mouse was placed back and exposed to one familiar object (cube) and a novel object (ball) for 10 min (test task). The objects were placed in the same locations as the objects in the pre-vious acquisition task. The total time spent for exploring each object (when the animal’s snout was directly toward the object at a distance ≤ 2 cm) was recorded, and the recognition index (RI) was defined as the am-ount of time exploring the novel object over the total time spent exploring both objects times [(time exp-loring the novel object/total time exploring both obj-ects)×100% ]. A higher RI indicates greater memory retention.
Y maze spontaneous alternation test
Spontaneous alternation in Y maze was tested 24 h after NORT. The Y maze has three arms separated from each other at the same angle (120°), and each arm was 30 cm long, 12 cm high, and 5 cm wide. Each mouse was placed at the intersection point of three arms and allowed to move freely for 8 min. The ent-ries of mouse into each arm were recorded and every entry different from last two entries was considered as a successful alternation. The alternation percentage, an index of spatial working memory, was calculated according to the following: (number of alternations)/ (total number of arm entries –2)×100%.
Morris water maze test
After Y maze test, the mice were subjected to the Morris water maze (MWM) test to evaluate the long-term spatial memory of mice. The maze was a circular pool with a diameter of 120 cm and a wall he-ight of 50 cm. The pool was filled with water at a temperature of 23±2°C, and the water was made opaque with non-toxic white tempera paint. Various prominent visual cues were positioned on the inner wall of the pool. A platform (diameter, 12 cm; height, 29 cm) was submerged approximately 1.0 cm below the wat-er surface at the midpoint of one quadrant of the pool. The swimming activity of each mouse was monitored via a camera mounted overhead, and a video tracking system (Ethovision 3.0, Noldus Information Technology) was used to collect movement information (latency, swim path, distance, and speed) of mice. In the hidden platform test, each mouse was trained four times per day for 5 consecutive days. On each experimental day, a trial was initiated by placing each mouse in the water facing the pool wall in one of the four quadrants and allowing it to swim freely to the escape platform. When mouse found the platform, it was allowed to stay on it for 5 s. If it did not find the platform within 60 s, the mouse was guided gently to the platform. And then the mouse was placed back to the home cage for 20 s before the next trial. On the next day after the hidden platform tests, each animal was given 60 s probe trial to evaluate its memory ret-ention ability. During the probe test, the platform was removed and the searching behavior of mice in the target quadrant (where the platform was located during hidden training) was measured.
In vivo hippocampal L-LTP recording
After behavioral tests, in vivo hippocampal L-LTP recording was performed. Mice were anesthetized with 5% chloral hydrate (0.07 ml/10 g, i.p.) and pla-ced in a stereotaxic device for acute surgery and L-LTP recording. A bound stimulating/recording electrode was inserted into the CA1 region of hippo-campus, with a tip location of the recording electr-ode at 2.0 mm posterior to the bregma and 1.5 mm lat-eral to the midline. Baseline field excitatory postsyn-aptic potentials (fEPSPs) were monitored for 30 min with repeated test stimuli (intensity, 30–50% of maxi-mal EPSPs; frequency, 0.033 Hz) delivered to the Schaffer-collateral/commissural pathway. High-fre-quency stimulation (HFS) was applied to induce L-LTP of fEPSPs. The HFS consisted of 3 series sti-muli with a 5 min interval, each series containing 3 trains of 20 stimuli with an inter-stimulus interval of 5 ms (200 Hz) and an inter-train interval of 30 s, as described in our previous studies [7, 8]. After 3 series of HFS, fEPSPs were evoked at 0.033 Hz for at least 3 h. The slope of fEPSP was normalized to ba-sal fEPSP and averaged. To test whether presynaptic mechanism was involved, paired-pulse facilitation (PPF), a short-term enhancement of synaptic transmission, was also observed in the CA1 region by us-ing paired stimuli with an interval of 50 ms before HFS. The change in the PPF ratio (calculated by dividing the slope of the second fEPSP by the slope of the first fEPSP) reflects the alteration of neurotransmitter release from presynaptic terminals.
Immunohistochemistry and immunofluorescence labeling
After completion of LTP recording, the mice (n = 6 in each group) were sacrificed by heart perfusion fix-ation with phosphate-buffered saline (PBS) and 4% paraformaldehyde (PFA). The brains of the mice were rapidly dissected and post-fixed for 24 h at room tem-perature. Then the brains of the mice were dehydra-ted. The brain blocks were sliced into 25μm-thick coronal sections. For immunohistochemical staining, the sections were incubated in 5% H2O2 at room tem-perature for 15 min, followed by washing 3 times with 0.01 M PBS. After blocking in 5% goat serum (Solarbio, China) for 30 min, brain slices were incubated with primary antibodies [anti-β-Amyloid 1–16 anti-body (6E10), dilution 1:500, 803105, Biolegend, USA; anti-GFAP antibody, dilution 1:200, bs-0199R, Bioss, China, Anti-Iba1 antibody, dilution 1:200, ab178846, Abcam, UK] overnight at 4°C, and secondary antibodies (Biotin-goat anti-mouse IgG, dilution 1:100, BST10G22C01, Boster, China; Biotin-goat anti-rabbit IgG, dilution 1:100, BST10G22C03, Boster) were incubated at room temperature for 2 h after was-hing the primary antibodies with PBS three times. Then, the brain sections were incubated with ABC complex (1:200) and DAB method was applied for positive coloration. Images were analyzed with Ima-ge-Pro Plus 6.0 (Media Cybernetics, USA). For imm-unofluorescent staining, the sections were washed and blocked with 5% serum for 30 min. Then brain slices were incubated with primary antibodies [IL-1β antibody (M-20), dilution 1:200, sc-1251, Santa Cr-uz, USA] overnight at 4°C, and secondary antibodies [Alexa Fluor 488-conjugated Affinipure Bovine An-ti-Goat IgG (H+L)(1:100,116969, Jackson, USA] for 2 h at room temperature in the dark. The sections were incubated with DAPI for 10 min at room temperature. A laser scanning confocal microscope was used to ob-tain immunofluorescence images in the brain slices.
Western blot
One side of the mice hippocampus (n = 6 in each gr-oup) were dissected and used for measuring protein expression by western blot. The hippocampus was homogenized in tissue protein extraction reagent (Bo-ster, Inc.), supplemented with complete protease and phosphatase inhibitor (Boster, Inc.). The homogena-tes were centrifuged (30 min, 15,000 rpm, 4°C), and protein concentration was measured using BCA protein assay kit (Boster, Inc.). A total of 50μg of protein from each sample was used. Sample proteins were separated on 12% SDS-polyacrylamide gels. After electrophoresis, the proteins were transferred onto 0.45μm PVDF membranes (Solarbio, Inc. China) and nonspecific binding was blocked with 5% BSA (Solarbio, Inc.) in Tris-buffered saline containing 0.05% Tween-20 (TBST). The membranes were incubated with a primary antibody overnight at 4°C, followed by a secondary antibody for 2 h at 4°C. The primary antibodies used were as follows: Anti-Phospho-NF-κBp65 (Ser468) antibody (#3039, Cell Signaling Technology, USA, dilution 1:1000); Anti-NF-κBp65 antibody (#3034, Cell Signaling Technology, dilution 1:1000); Anti-Phospho-PKA R2/PKR2 (phospho S99) antibody (ab32390, Abcam, UK, dil-ution 1:1000); Anti-PKA R2/PKR2 antibody (ab3-8949, Abcam, dilution 1:3000); Anti-cAMP protein kinase catalytic subunit antibody (ab26322, Abcam, dilution 1:1000); Anti-phospho-CREB (phospho S133) antibody (ab32096, Abcam, dilution 1:500); Anti-CREB antibody (ab31387, Abcam, dilution 1:700). The secondary antibodies used were Biotin-goat anti-rabbit IgG (BST10G22C03, Boster, Inc., dilution 1:1000); Goat anti-rabbit IgG HRP (ZSGB-BIO, Inc. China, dilution 1:100,000). After rinsing with TBST, the immunocomplexes were visualized by chemiluminescence using the ECL (Beyotime, Inc.). The film signals were digitally scanned with a Fluor Chem Scanner (Protein Simple) and quantified with Alpha View SA software. β-actin (PR-0255 ZSGB-BIO Inc., dilution 1:3000) was utilized as an internal control for sample loading, and each blot was normalized to its corresponding β-actin value.
ELISA
The other side of the mice hippocampus (n = 6 in each group) were dissected and used to detect the contents of IL-1β and TNF-α by ELISA (SEA563Mu and SEA133Mu, CLOUD-CLONE CORP, USA). At the same time, ELISA was also used to detect insulin levels in mouse plasma (CEA448Mu, CLOUD-CL-ONE CORP). The mouse hippocampus was minced to small pieces and homogenized in fresh lysis buffer with a glass homogenizer on ice. The resulting suspe-nsion was sonicated with an ultrasonic cell disrupter till the solution is clarified. Then, the homogenates were centrifuged for 5 min at 10000 g. The supernat-ant was collected for analysis. And the mouse plasma was also collected for analysis. Prepare 5 wells for standard points and 1 well for blank. Three replicate wells were set for each concentration standard. Each well was added 50μl dilutions of standard, bla-nk and samples, and then 50μl detection reagent A was added to each well immediately. After incubation for 1 h at 37°C, 100μl of test solution B was added to each well. Then, 90μl of TMB substrate solution and 50μl of stop solution were added. The optical density of each well at 450 nm wavelength (OD value) was measured.
Statistical analysis
All values are expressed as means±standard erro-rs (SEM). Water maze navigation data were analyzed using three-way repeated measures analysis of variance (ANOVA). Other data were analyzed using two-way repeated measures ANOVA and post hoc Tukey’s test. Statistical significance was defined as p < 0.05, and all statistical analyses were performed using software packages SPSS 18.0 and SigmaPlot 12.0.
RESULTS
DAla2GIP-Glu-PAL treatment enhanced novel object recognition ability in APP/PS1 mice
Before object recognition test, the motor capability of mice was evaluated in the open field task. As shown in Fig. 1A, there was no significant difference in total moving distance among the four groups (APP/PS1: F (1,47) = 2.135, p = 0.151; DAla2GIP-Glu-PAL: F (1,47) = 0.440, p = 0.510; APP/PS1×DAla2GIP–Gl-u-PAL interaction: F (1,47) = 0.186, p = 0.669). Thus, no evidence showed that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment affect the motor capability of the mice. Based on the instinct of mice to explore novel things, NORT was performed on the second day after the open field test. Two-way ANOVA showed that APP/PS1 gene mutation and GIP analogs treatment had significant main effects on RI (APP/PS1: F (1,47) = 10.521, p = 0.002; DAla-2GIP-Glu-PAL: F (1,47) = 7.931, p = 0.007; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,47) = 4.554, p = 0.038). Tukey’s post hoc tests (Fig. 1B) showed that the RI in the APP/PS1+PBS mice (42.64% ±4.12%) significantly lower than that in the WT + PBS group (61.99% ±4.19%, p < 0.001), indicating that the APP/PS1 mice spent less time in exploring the novel object. However, the RI in the APP/PS1 + DA-la2GIP-Glu-PAL group (60.45% ±2.82%) had a significant increase compared with the APP/PS1 + PBS mice (p < 0.001).
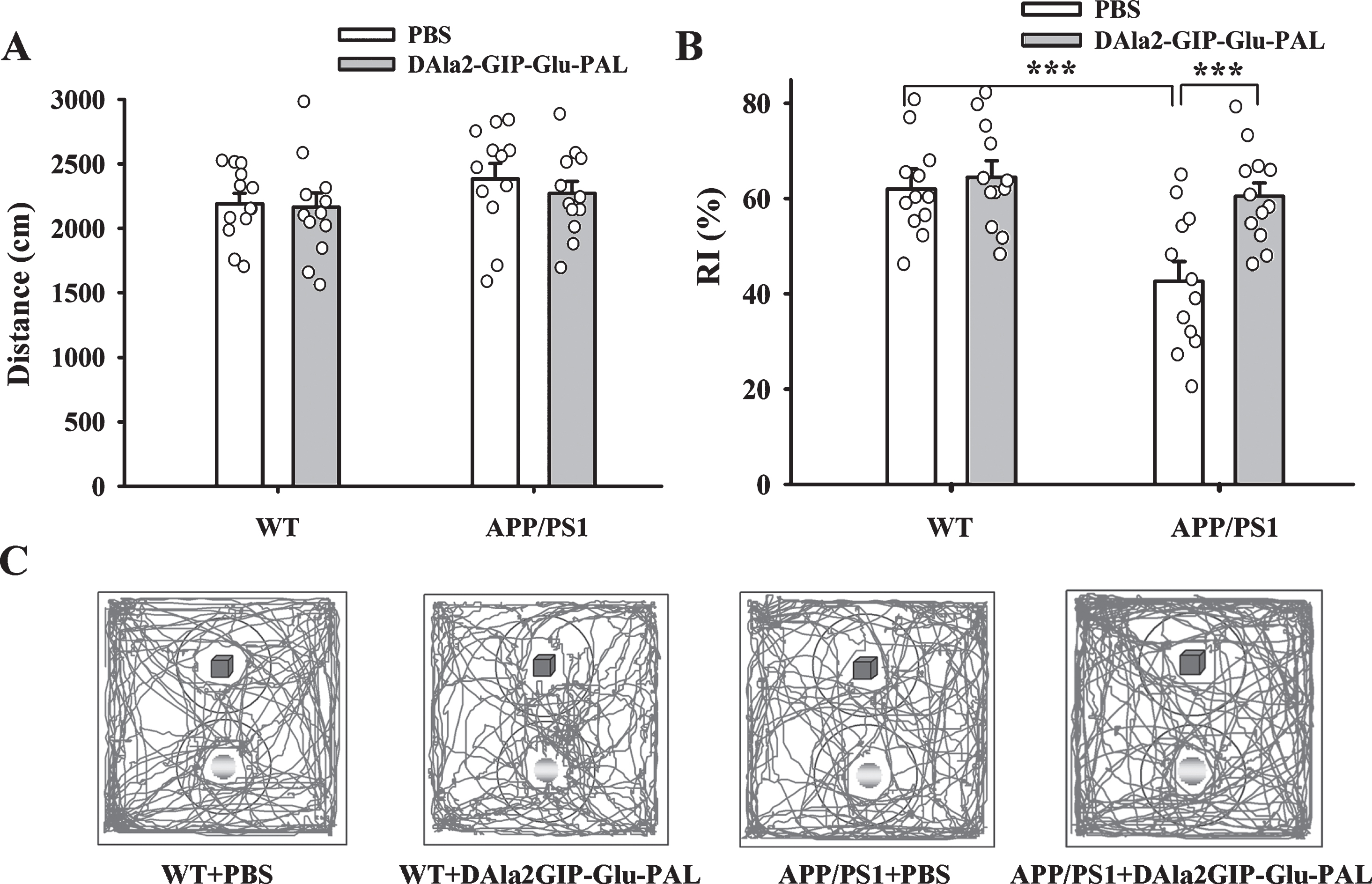
DAla2GIP-Glu-PAL treatment improved new object recognition ability in APP/PS1 mice. A) Histograms showing the total running distance of mice in the open field, no significant difference between groups (n = 12 for each group). B) Histograms showing the new object RI of mice. The reduced RI in APP/PS1 mice was significantly reversed by DAla2GIP-Glu-PAL treatment (***p < 0.001, n = 12 for each group). C) Representative running traces of mice in new object recognition test. The blue cube represents the familiar object and the yellow ball represents the novel object.
DAla2GIP-Glu-PAL alleviated working memory deficits in APP/PS1 mice
The spontaneous alternation of mice in Y maze was tested to examine the working memory of anim-als. Two-way ANOVA showed that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had sig-nificant main effects on the spontaneous alternation of the mice (APP/PS1: F (1,47) = 13.723, p < 0.001; DAla2GIP-Glu-PAL: F (1,47) = 19.761, p < 0.001; APP/PS1×DAla2GIP-Glu-PAL interaction: (F (1,47) = 8.369, p = 0.006). Tukey’s post hoc tests showed that the percentage of correct alternation in APP/PS1 + PBS (46.00% ±1.99%) was significantly lower than that in WT + PBS group (61.58% ±2.27%, p < 0.01). After treatment with DAla2GIP-Glu-PAL, the correct alternation percentage in APP/PS1+DAla2GIP-Glu-PAL group (63.33% ±2.64%) had a significant incre-ase (p < 0.01) compared with APP/PS1 + PBS mice (Fig. 2A). Meantime, the total arm entries of mice did not show any significant difference among these gro-ups (p > 0.05, Fig. 2B), suggesting that the differences in spontaneous alternation among groups were due to impairment of spatial working memory rather than a disability of locomotor activity.

DAla2GIP-Glu-PAL treatment alleviated the working memory deficits of APP/PS1 mice in Y maze test. A) Histograms showing lower spontaneous alternation in the APP/PS1+PBS group and significant recovery after treatment with DAla2GIP-Glu-PAL (*p < 0.05, **p < 0.01, n = 12 for each group). B) Histograms showing no significant difference in total arm entries among the different groups.
DAla2GIP-Glu-PAL improved long term spatial learning and memory of APP/PS1 mice
MWM was used to assess the long-term spatial learning and memory of mice. The learning ability of mice to acquire spatial information was first ass-essed by a consecutive 5 days of hidden platform test. As shown in the Fig. 3A and 3C and Table 1, the esc-ape latency in all groups gradually decreased with the increase of training days. The mean escape latency had a significant increase in the APP/PS1 + PBS gro-up on days 3–5, compared with the WT + PBS group. Importantly, the increased escape latency was signifi-cantly decreased in the APP/PS1 + DAla2GIP-Glu-PAL group on days 4–5. This result indicates that chronic DAla2GIP-Glu-PAL treatment could impr-ove the spatial learning ability of APP/PS1 mice.
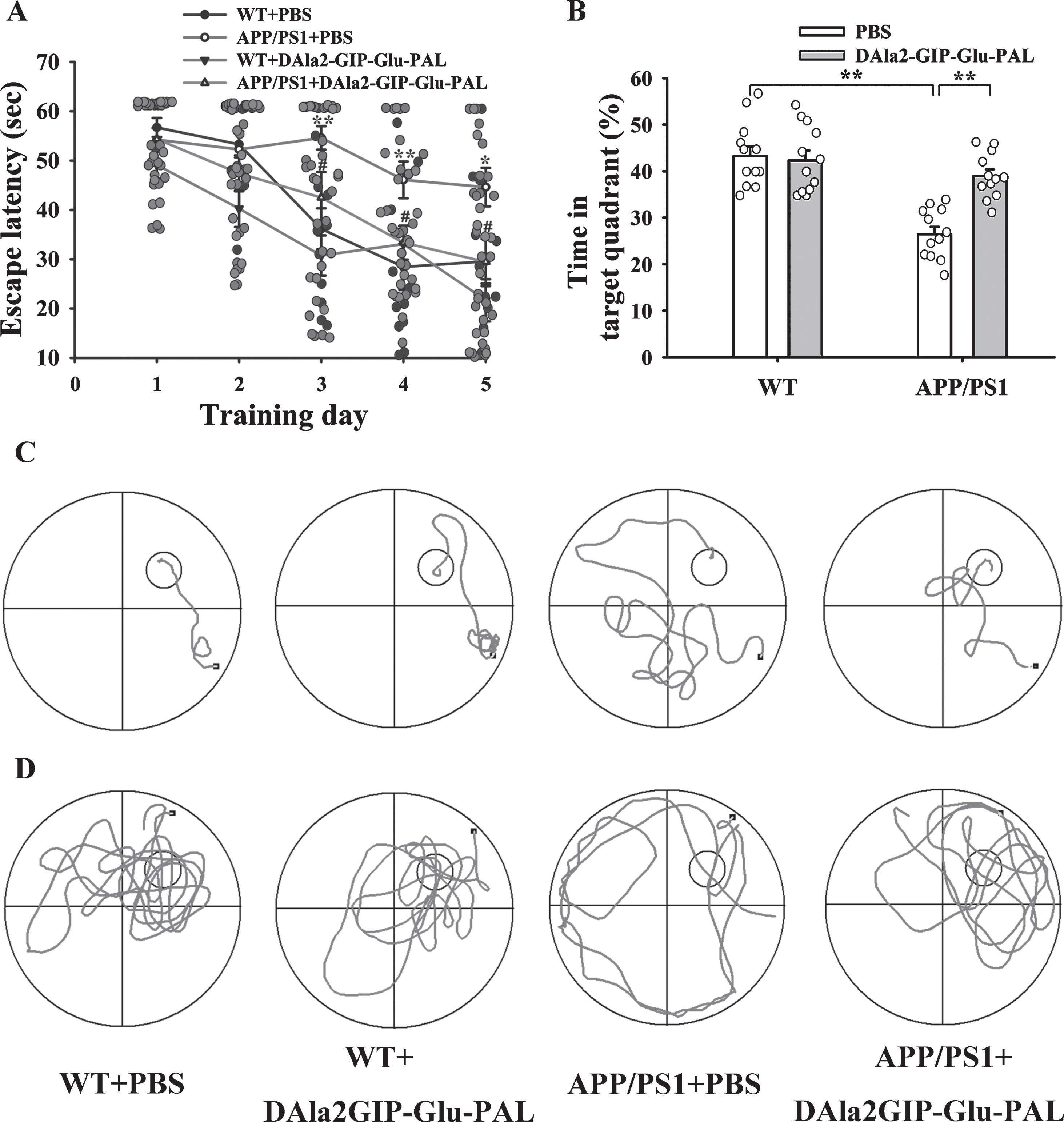
DAla2GIP-Glu-PAL treatment improved spatial learning and memory of APP/PS1 mice. A) Plots showing the changes in average escape latencies of mice in searching for the hidden platform during five consecutive training days (**p < 0.01 versus WT+PBS group; # #p < 0.01 versus APP/PS1+PBS group, n = 12 for each group). B) Histograms showing the swimming time percentage of the mice in the target quadrant, with a significant decrease in the APP/PS1-PBS group and a reversal after DAla2GIP-Glu-PAL treatment. C) Sample traces of mice in different groups in the hidden platform test on 4th day. D) Representative swimming traces of mice in the four groups during probe trials. The large circle represents the water maze pool, and the small circle represents the platform.
Mean escape latencies of mice during 5 training days (s, Mean±S.E.M.)
**p < 0.01 versus WT+PBS group; # #p < 0.01 versus APP/PS1+PBS group.
Then, spatial memory ability was tested on the 6th day with probe trials. The platform was removed, and mice were allowed to swim in the pool for 60 s. As shown in the Fig. 3B and 3D, the swimming time percentage of the mice in the target quadrant was 43.29±2.06%, 42.29±2.13%, 26.43±1.61%, and 39.00% ±1.38% in WT + PBS, WT + DAla2GIP-Glu-PAL, APP/PS1 + PBS, and APP/PS1 + DAla2GIP-Glu-PAL groups, respectively. Two-way ANOVA showed that there were significant main effects of APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment (APP/PS1: F (1,47) = 30.560, p < 0.001; DAla2GIP-Glu-PAL: F (1,47) = 10.161, p = 0.003), and a significant interaction between APP/PS1 gene mut-ation and DAla2GIP-Glu-PAL (F (1,47) = 13.831, p < 0.001). Tukey’s post hoc test showed a significant difference in memory retrieval between the APP/PS1 + PBS and WT + PBS groups (p < 0.001). Similarly, DAla2GIP-Glu-PAL treatment reversed the APP/PS1 gene mutation-induced reduction of swimming time in the target quadrant (p < 0.01). These results indicated that pretreatment with DAla2GIP-Glu-PAL protected against the spatial memory deficit in APP/PS1 mice.
To exclude the possibility that the difference in esc-ape latency might be caused by drug-induced visual and locomotor deficits, a visual platform test was performed after the probe trials. Statistical analysis showed that there was no significant difference in the swimming speed in probe trials and the swimming time arriving to the visible platform between groups (p > 0.05), suggesting that the changes in escape latency and swimming time percentage just resulted from the impairments of spatial learning and memory.
DAla2GIP-Glu-PAL rescued in vivo hippocampus L-LTP in APP/PS1 mice
To clarify the possible electrophysiological mech-anism underlying the neuroprotective effects of DAl-a2GIP-Glu-PAL on learning and memory, we inves-tigated the effects of DAla2GIP-Glu-PAL on the long-term synaptic plasticity by recording in vivo fEPSP and its potentiation in the hippocampus CA1 reg-ion. After 30 min of stable basal fEPSP recording, three series of HFS were given to induce L-LTP. The slopes of fEPSPs at 60 min, 120 min, and 180 min af-ter HFS were compared between groups (Fig. 4). Two-way ANOVA demonstrated that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had significant main effects on the fEPSP slope at 60 min (APP/PS1: F (1,26) = 3.321, p = 0.028; DAla2GIP-Glu-PAL: F (1,26) = 4.380, p = 0.005; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,26) = 4.917, p = 0.002), 120 min (APP/PS1:F (1,26) = 4.799, p = 0.003; DAla2GIP-Glu-PAL: F (1,26) = 4.215, p = 0.007; APP/PS1DAla2GIP-Glu-PAL interaction: F (1,26) = 6.096, p < 0.001), and 180 min (APP/PS1: F (1,26) =6.802, p < 0.001; DAla2GIP-Glu-PAL: F (1,26) = 4.425, p = 0.005; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,26) = 8.546, p < 0.001) post-HFS. Tukey’s post hoc test shown that the slopes of the fEPSPs imm-ediately after HFS abruptly increased from 100% to 219.5±1.16%, 202.34±4.39%, 220.93±2.74%, and 220.32±4.19% in the WT+PBS, APP/PS1+PBS, WT+DAla2GIP-Glu-PAL, and APP/PS1+Dala2GIP-Glu-PAL groups, respectively, indicating that LTP was successfully induced in the four groups (Fig. 4A). However, the L-LTP values in the APP/PS1-PBS group obviously decreased 40 min post-HFS. As shown in the Fig. 4A and 4C, compared with the WT+PBS mice, the L-LTP value was sig-nificantly suppressed at 60 min (131.92±8.57%, p < 0.05), 120 min (121.76±6.36%, p < 0.05), and 180 min (103.57±1.67%, p < 0.05) in APP/PS1+PBS mice. In contrast, there was a relatively large maintenance of L-LTP in the APP/PS1+DAla2GIP-Glu-PAL group at 60 min (177.28±5.97%, p < 0.05), 120 min (165.58±6.06%, p < 0.05), and 180 min (162.41±7.08%, p < 0.05) compared with the APP/PS1 + PBS group.
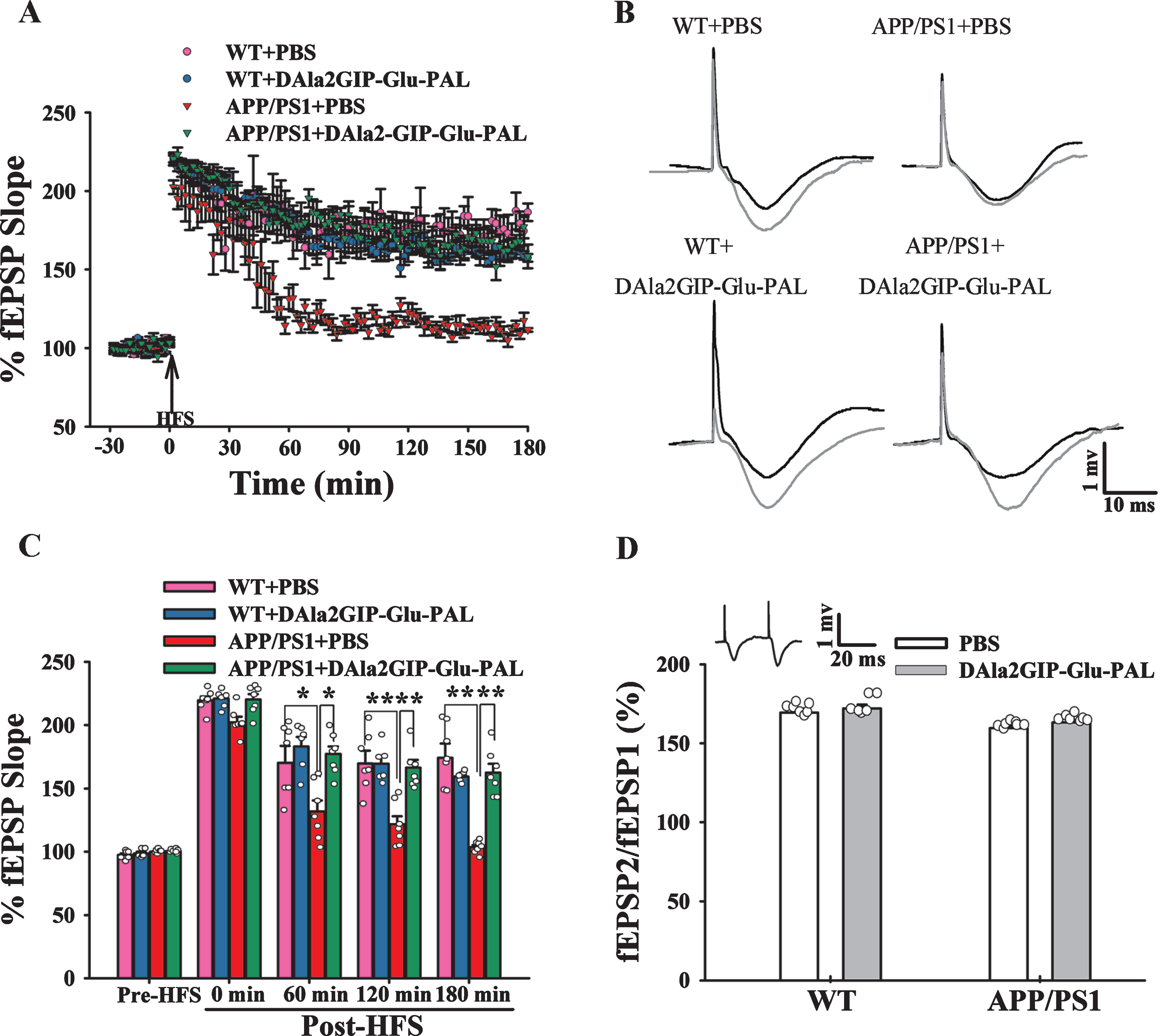
DAla2GIP-Glu-PAL treatment reversed hippocampal L-LTP suppression in APP/PS1 mice. A) Plots showing the test stimuli-induced fEPSPs in the hippocampal CA1 region of APP/PS1 mice before and after HFS (n = 6–7). B) Typical fEPSP traces recorded before (black line) and 3 h after (grey line) HFS in the four groups. Scale bars, 1 mV and 10 ms. C) Histograms showing the normalized fEPSP slope pre-HFS, 0 min, 60 min, 120 min and 180 min post-HFS (*p < 0.05 and **p < 0.01). D) Histograms showing the ratios of paired pulses in different groups (n = 6–7). Inset: a sample trace of fEPSP induced by paired pulses.
Further, paired pulse facilitation (PPF) was examined to detect the possible involvement of presynaptic mechanism in the effects of DAla2GIP-Glu-PAL on the LTP (Fig. 4D). Application of paired pulses to the Schaffer collaterals always induced PPF (see inset of Fig. 4D). Two-way ANOVA showed that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had no significant main effects on PPF (APP/PS1: F (1,26) = 0.369, p = 0.550; DAla2GIP-Glu-PAL: F (1,26) = 0.003, p = 0.956; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,26) = 0.298, p = 0.590). Thus, no evidence showed that APP/PS1 ge-ne mutation and DAla2GIP-Glu-PAL treatment affected presynaptic neurotransmitter release in the hippocampal CA1 region.
DAla2GIP-Glu-PAL treatment reduced Aβ plaques in the hippocampus of APP/PS1 mice
High-density of Aβ plaques is typical pathologi-cal hallmarks in the AD brains. We further evaluated the histopathological changes in the hippocampus of mice using immunohistochemistry and specific antibodies 6E10 (Fig. 5). Two-way ANOVA showed that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had significant main effects on the numbers of 6E10 positive plaques (APP/PS1: F (1,23) = 132.211, p < 0.001; DAla2GIP-Glu-PAL: F (1,23) = 20.039, p < 0.001; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,23) = 20.039, p < 0.001). As shown in the Fig. 5A, 6E10 positive plaques could be easily found in the hippocampus of APP/PS1 mice under low magnification. Tukey’s post hoc test showed that the number of Aβ plaques in APP/PS1+PBS group (15.17±1.64) was significantly more than that in WT+PBS group (0.00±0.00, p < 0.001), while DAla2GIP-Glu-PAL treatment effectively reduced the number in the hippocampus of APP/PS1 mice (6.67±0.95, p < 0.001). This result demonstrated that DAla2GIP-Glu-PAL treatment could attenuate cerebral pathological change in the APP/PS1 transgenic mice.
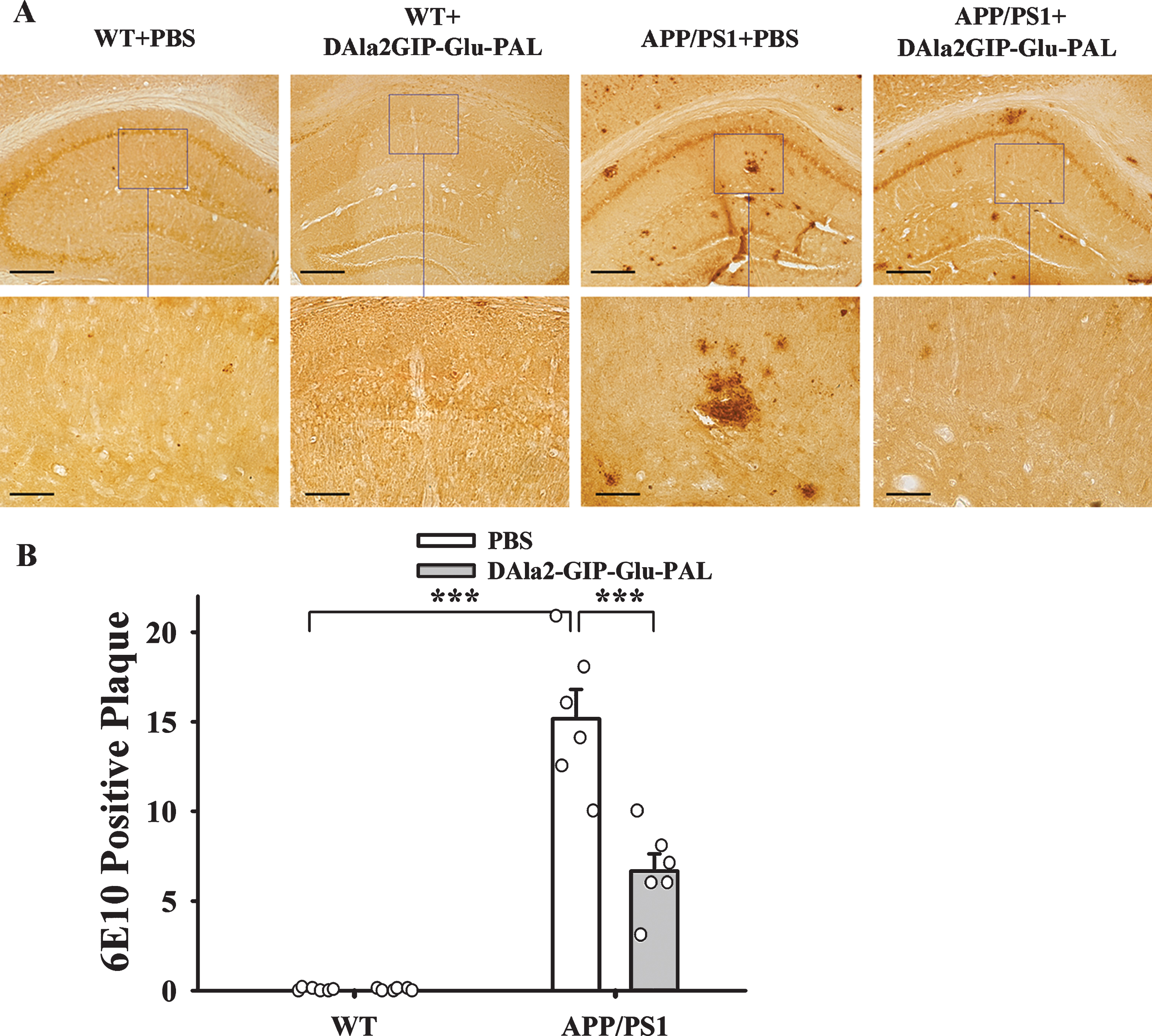
DAla2GIP-Glu-PAL treatment reduced Aβ plaques in the hippocampus of APP/PS1 mice. A) Photographs showing the distribution of Aβ immunoreactivity-positive plaques in the hippocampus of mice after treatment with DAla2GIP-Glu-PAL or PBS. The bars are 100μm and 50μm in the upper and lower rows, respectively. B) Histograms showing a significant decrease in number of Aβ plaques in the hippocampus of APP/PS1 mice after DAla2GIP-Glu-PAL treatment (***p < 0.001).
DAla2GIP-Glu-PAL attenuated inflammatory response in the hippocampus of APP/PS1 transgenic mice
In view of the fact that Aβ can promote glia cell proliferation and trigger inflammatory responses in the brain, we further examined the effects of DAla2GIP-Glu-PAL injection on the inflammatory responses in APP/PS1 mice by using immunohisto-chemistry, immunofluorescence, and ELISA. As shown in the Fig. 6, APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had significant main effects on the intensity of GFAP and Iba1, two cell markers for astrocyte and microglia (GFAP: APP/PS1:F (1,23) = 43.197, p < 0.001; DAla2GIP-Glu-PAL: F (1,23) = 13.259, p = 0.002; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,23) = 8.459, p = 0.009. Iba1: APP/PS1:F (1,23) = 78.572, p < 0.001; DAla2GIP-Glu-PAL: F (1,23) = 28.131, p < 0.001; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,23) = 16.683, p < 0.001). Tukey’s post hoc test showed that the GFAP (Fig. 6C) and Iba1 (Fig. 6D) immunoreactivity significantly increased in APP/PS1+PBS group compared with that in WT+PBS group (p < 0.001), while the increased levels of GFAP and Iba1 had significant reduction in DAla2GIP-Glu-PAL treated APP/PS1 group (p < 0.001).
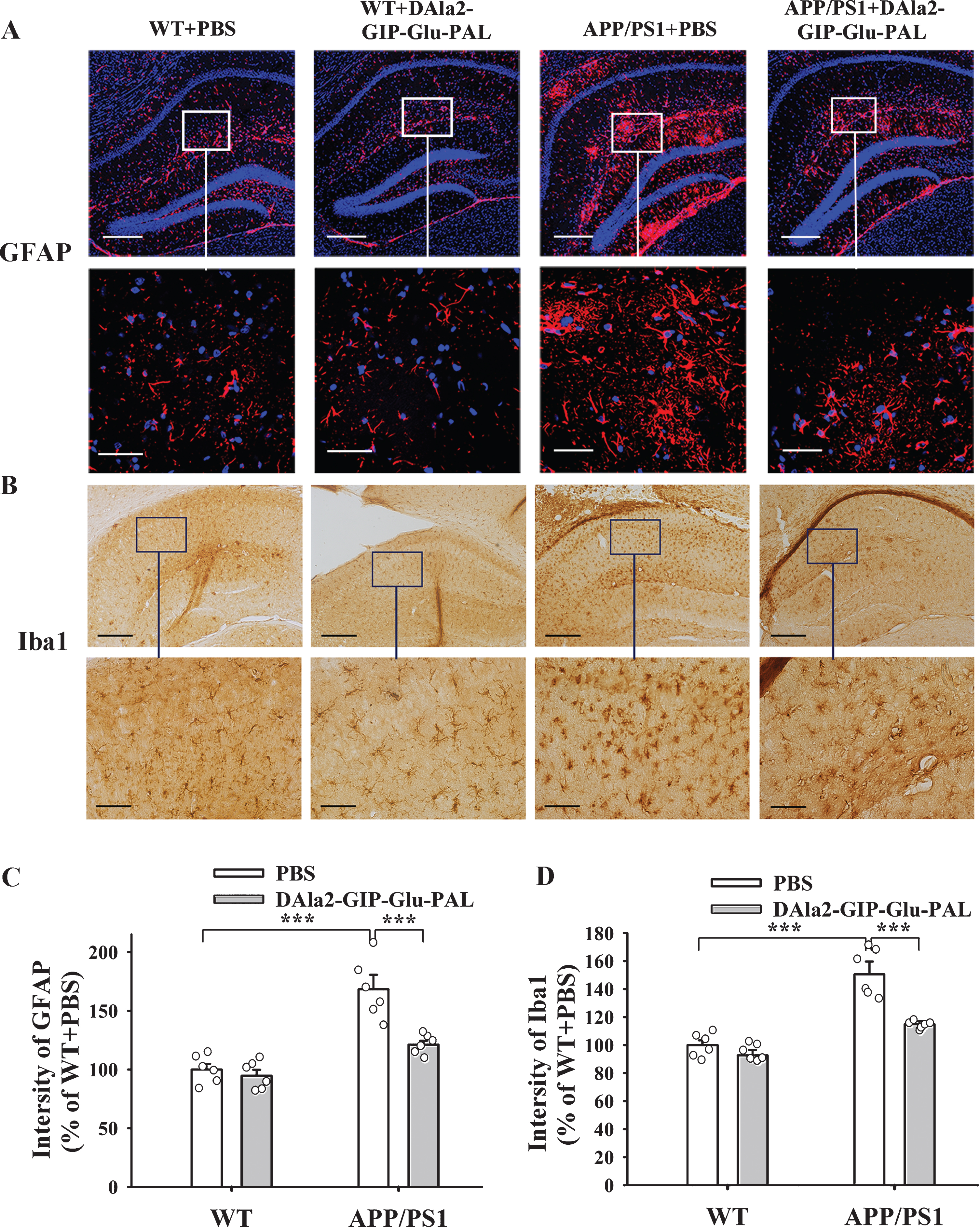
DAla2GIP-Glu-PAL attenuated astrocytosis and microcytosis in the hippocampus of APP/PS1 mice. A) Photographs showing immunofluorescence staining of GFAP positive cells. B) Photographs showing immunohistochemical staining of Iba1 positive cells. Bar = 100μm or 50μm in upper and lower row, respectively. C) and D) Histograms showing that the elevated GFAP and Iba1 immunopositive staining in the APP/PS1+PBS mice had significant declines after DAla2GIP-Glu-PAL treatment (***p < 0.001).
Because Aβ can stimulate astrocytes and micro-glia to synthesize and secrete inflammatory factors, we further observed the inflammatory factor IL-1β by immunofluorescence technique. As shown in the Fig. 7A, there were few IL-1β (green) immunopositive cells in the hippocampus of the WT+PBS and WT+DAla2GIP-Glu-PAL mice, while a lot of IL-1β positive cells were found in the APP/PS1+PBS group. The increased IL-1β positive cells were significantly reduced after treatment with DAla2GIP-Glu-PAL. We further tested the contents of IL-1β and TNF-α in the hippocampus by ELISA. Two way ANOVA showed that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had significant main effects and interaction on IL-1β and TNF-α con-tents (IL-1β: APP/PS1: F (1,23) = 159.271, p < 0.001; DAla2GIP-Glu-PAL: F (1,23) = 75.837, p < 0.001; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,23) = 47.718, p < 0.001. TNF-α: APP/PS1: F (1,23) = 197.469, p < 0.001; DAla2GIP-Glu-PAL: F (1,23) = 47.965, p < 0.001; APP/PS1×DAla2GIP-Glu-PAL int-eraction: F (1,23) = 64.808, p < 0.001). Tukey’s post hoc test showed that IL-1β and TNF-α contents were significantly increased in the APP/PS1+PBS group (796.95±26.20, 841.91±34.62) compared with the WT+PBS group (410.40±15.16, 344.25±14.78 p < 0.001), while treatment with DAla2GIP-Glu-PAL significantly reduced the values of IL-1β and TNF-α in the APP/PS1+DAla2GIP-Glu-PAL group (487.83±13.64, 504.73±15.98, p < 0.001, Fig. 7B, C).
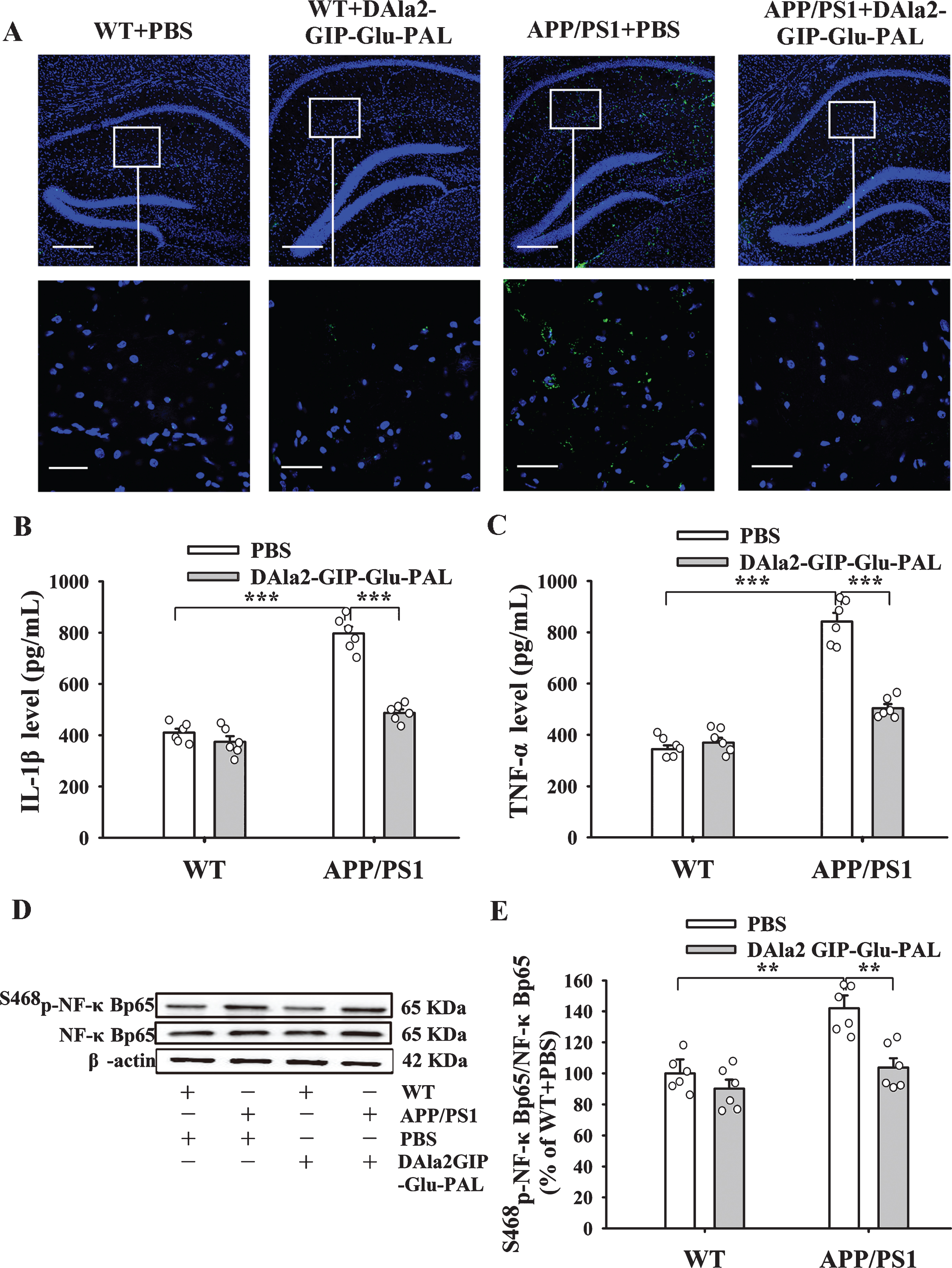
DAla2GIP-Glu-PAL inhibited inflammatory factors in the hippocampus of APP/PS1 mice. A) Photographs showing the immunofluorescence of IL-1β (green) positive cells in the hippocampus. The blue dots represent DAPI stained neuronal nuclei. The scale bars in the upper and lower rows are 100μm and 50μm, respectively. B) and C) Histograms showing the changes of IL-1β and TNF-α expression measured by ELISA (***p < 0.001). D) Representative Western-blotting bands for Ser468p-NF-κBp65, β-actin as an internal reference. E) Histograms showing that the increased level of NF-κBp65 (Ser468) in the APP/PS1+PBS group was significantly decreased by DAla2GIP-Glu-PAL (**p < 0.01).
In addition, considering that IL-1β and TNF-α can activate NF-κB to enter the nucleus and upregulate the expression of inflammatory factors, and trigger the inflammatory cells proliferation [32, 33], we also examined the expression level of NF-κBp65 (Ser468) in the hippocampus by western blot (Fig. 7D, E). Two-way ANOVA showed that APP/PS1 gene mutation and DAla2GIP-Glu-PAL treatment had significant main effects and interaction on NF-κBp65 (Ser468) expression level (APP/PS1: F (1,23) = 19.269, p < 0.001; DAla2GIP-Glu-PAL: F (1,23) = 14.293, p < 0.001; APP/PS1×DAla2GIP-Glu-PAL interaction: F (1,23) = 4.964, p = 0.038). Tu-key’s post hoc test showed that the value of NF-κBp65 (Ser468) in the hippocampus of APP/PS1+PBS mice (141.95% ±8.31%) was significantly hi-gher than that in other groups (p < 0.01), while DAla2GIP-Glu-PAL significantly reduced the expres-sion level of NF-κBp65 in APP/PS1 mice (p < 0.01). The value of NF-κBp65 (Ser468) in APP/PS1+DAla2GIP-Glu-PAL group (103.86% ±5.94%) was close to the level of normal control.
These results above indicate that chronic i.p. injection of DAla2GIP-Glu-PAL attenuated inflammatory response in the hippocampus of APP/PS1 mice.
The DAla2GIP-Glu-PAL treatment upregulated the expression levels of cAMP, S99p-PKA, and S133p-CREB in the hippocampus of APP/PS1 transgenic mice
GIP receptors belong to G-protein-coupled receptor superfamily. To further clarify the mechanism of DAla2GIP-Glu-PAL improving pathology and synaptic plasticity in APP/PS1 transgenic mice, we detected the expression levels of cAMP, S99p-PKA, and S133p-CREB in the hippocampus using wes-tern blotting techniques. As shown in Table 2, two-way ANOVA analysis showed that APP/PS1 gene mutation and DAla2GIP -Glu-PAL drug treatment have obvious main effects and interaction effects on the levels of cAMP, S99p-PKA, and S133p-CREB.
Two-way analysis of variance in hippocampus of each group of mice (n = 6 in each group)
Figure 8 demonstrated representative western blotting bands of cAMP, S99p-PKA, and S133p-CREB and statistical histograms in each group. Tukey’s post hoc test showed that the relative gray values of cAMP (Fig. 8A, B), S99p-PKA (Fig. 8C, D), and S133p-CREB (Fig. 8E, F) in the hippocampus of APP/PS1+PBS group were significantly decre-ased (p < 0.01) compared with those in WT+PBS group, while chronic treatment with DAla2GIP-Glu-PAL basically reversed all of these changes (p < 0.01). These results indicate that DAla2GIP-Glu-PAL upregulated cAMP/S99p-PKA/S133p-CREB signaling pathway in the hippocampus of APP/PS1-PBS mice.
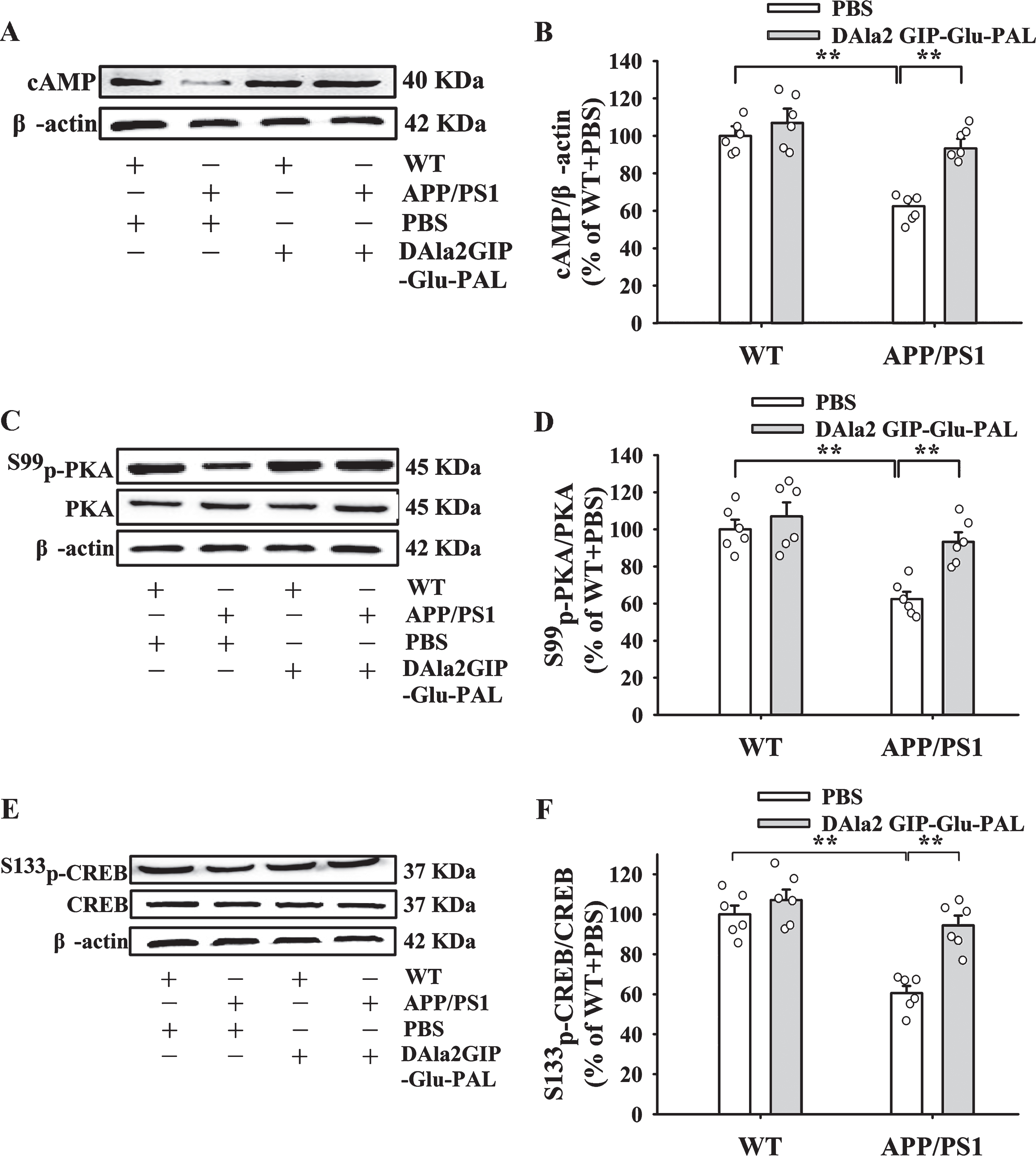
DAla2GIP-Glu-PAL upregulated the levels of cAMP, S99p-PKA and S133p-CREB in the hippocampus of APP/PS1 mice. A, C, E) Representative western blotting bands for cAMP, S99p-PKA, and S133p-CREB in different groups. DAla2GIP-Glu-PAL reversed the changes of protein content in hippocampus of APP/PS1 transgenic mice. B, D, F) Statistical histograms showing the relative gray values of cAMP, S99p-PKA/PKA, and S133p-CREB/CREB in each group. All values were expressed as percentages of WT+PBS mice. Significance was determined using a two-way ANOVA and post hoc Tukey’s multiple comparison tests (**p < 0.01).
DISCUSSION
Spatial working memory and reference memory are dependent on hippocampus functions [3]. Therefore, the memory disorder will gradually aggravate with the development of hippocampus dysfunction during AD processes. Although the exact pathogenesis of AD remains elusive, mutations of several genes such as amyloid-β precursor protein (APP), presenilin-1 (PS1), and presenilin-2 (PS2) has been closely linked to AD [32]. Moreover, the APP/PS1 mice with memory deficits and hippocampus pathology have been widely used in the AD research [33, 34]. In the present study, 9-month-old APP/PS1 mice were subjected to multiple behavioral tests. The decreased RI in NORT and reduced spontaneous alternation in Y maze clearly indicate impairment in short-term memory in the APP/PS1 mice, while the increased escape latency in place navigation and decreased swimming time in probe test in MWM suggest an obvious decline in spatial long-term learning and memory in APP/PS1 mice. These results are consistent with previous reports for the APP/PS1 mouse model of AD [18, 36]. Importantly, we found for the first time that i.p. injection of DAla2GIP-Glu-PAL effectively alleviated the cognitive deficits in the APP/PS1 transgenic mice, with significant improvements in working memory and long-term reference memory. These behavioral experiments suggest that DAla2GIP-Glu-PAL treatment may be an effective strategy for improvement of cognitive decline in AD.
Due to a close association between spatial memory and hippocampal synaptic plasticity, hippocampal LTP has been deemed to be a major cellular mechanism underlying learning and memory [37, 38]. Previously, we have demonstrated that intracerebroventricular injection of Aβ1–42 suppressed in vivo hippocampus L-LTP in normal rats [3]. In the present study, we confirmed that the in vivo L-LTP in the hippocampus CA1 region was also seriously impaired in the APP/PS1 double transgenic mice. The L-LTP value was significantly suppressed at 60 min, 120 min, and 180 min in APP/PS1 mice. Unlike early phase LTP (E-LTP), the L-LTP is involved in protein synthesis that is closely related to long-term memory. Of particular note is the L-LTP recordings in the present study were conducted on the same behaviorally trained mice, which thus providing further insights into the relationship between the spatial memory and hippocampal L-LTP in the APP/PS1 mice. Interestingly, DAla2GIP-Glu-PAL treatment improved hippocampus synaptic plasticity in the transgenic mice, with relative larger and longer maintenance of L-LTP in the DAla2GIP-Glu-PAL treated APP/PS1 mice. Therefore, the electrophysiological results of L-LTP support the behavioral improvement induced by DAla2GIP-Glu-PAL in the APP/PS1 mice.
Aβ is a main pathological biomarker in the AD brain, which triggers not only synaptic dysfunction and cognitive disorder, but also astrocyte and microglia proliferation and other neuroinflammatory responses [39–42]. The present study further confirmed that large amounts of Aβ plaques, astrogliosis, and microgliosis appeared in the hippocampus of 9-month-old APP/PS1 mice using immunohistochemistry and immunofluorescence staining. Besides, ELISA and western blot results showed that the contents of IL-1β, TNF-α, and phosphorylated NF-κBp65 in the brain of APP/PS transgenic mice were also dramatically increased. These results indicate that inflammatory damages such as astrogliosis, microgliosis and various inflammatory factors are closely associated with Aβ deposition in the brain of APP/PS1 transgenic mice. As we know, NF-κB is widely expressed in human tissue cells and is involved in cell proliferation, differentiation, apoptosis, and inflammation. When cells are stimulated by appropriate extracellular substances such as proinflammatory cytokines and neurotoxic Aβ, NF-κB can be activated and enters the nucleus to regulate gene expression of various proteins [43]. Some studies have confirmed that inflammatory factors such as TNF-α and IL-1β can activate NF-κB, which in turn not only upregulates inflammatory factors and triggers inflammatory cell proliferation such as astrocytes [44, 45], but also upregulates the expression of β-secretase and APP genes, and promotes Aβ production and deposition [46, 47]. So, the mutual dependence and mutual promotion between inflammatory response and pathological Aβ in the brain would greatly accelerate the progression of AD. Interestingly, chronic treatment with DAla2GIP-Glu-PAL in the present study broke the vicious circle and effectively reduced Aβ plaques and attenuated various parameters of inflammation, including astrogliosis, microgliosis, and inflammatory factor (IL-1β, TNF-α, and NF-κB) elevation. In these anti-inflammation effects of DAla2GIP-Glu-PAL, the effects on NF-κB appeared to be most prominent, in which their responses in APP/PS1 mice were almost completely prevented. We noticed that liraglutide, another GLP-1 analogue, also reduced hippocampal inflammation including reactive microgliosis and astrogliosis [47, 48], which supports our results above. Therefore, the present study justified that DAla2GIP-Glu-PAL treatment can effectively attenuated these cerebral pathological changes from Aβ plaques to inflammatory factors.
GIP, as an incretin growth factor, has a signaling pathway parallel to insulin, but with different receptors. GIP receptor (GIPR) expression has been found in large neurons such as the pyramid neurons in the cortex and hippocampus, granule neurons in the dentate gyrus, Purkinje neurons in the cerebellum, and basal brain areas [49]. It has been reported that activation of GIPRs by native GIP or protease-resistant analogs, such as Nacetyl-GIP, enhanced synaptic plasticity and protected synapses from the detrimental effects of Aβ fragments on LTP formation. On the contrary, GIPR KO mice showed clear impairments in memory formation, synaptic plasticity, and progenitor cell proliferation [50]. And GIP(3-30)NH2, a reversible competitive antagonist of GIPR in rodents [51] and as well as humans [52, 53], effectively antagonized GIP-mediated G protein-signaling as well as pancreatic hormone release. These findings clearly demonstrate that the GIPR plays an important role in neuronal communication and brain function. Therefore, we speculate that the neuroprotective effects of DAla2GIP-Glu-PAL by i.p. injection are mediated by GIPRs in the brain. As a GIPR, the activation of GIPR is coupled to increases in cAMP and intracellular Ca2 + levels, as well as activation of PI3K, PKA, and PKB. Our study also showed that the levels of cAMP/p-PKA/p-CREB in the hippocampus of APP/PS1 mice were significantly decreased, while chronic treatment with DAla2GIP-Glu-PAL effectively reversed the decline in these signal molecules. These results indicate that DAla2GIP-Glu-PAL, by upregulating cAMP/p-PKA/p-CREB signal pathway, can improve cognitive function and synaptic plasticity, and ameliorate pathological damage and inflammatory injury in the hippocampus of APP/PS1 mice. Li et al. also reported that DAla2GIP-Glu-PAL reduced damage of dopaminergic neurons and neuroinflammation in Parkinson’s disease mouse model by increasing CREB and Bcl-2 expression in the brain [29]. In addition, upregulation of PI3K/AKT1 and downregulation of GSK3β in the hippocampus of APP/PS1 transgenic mouse might be also linked to the neuroprotection of DAla2GIP-Glu-PAL [51, 54].
Much of the science including our laboratory has performed some pioneering, preliminary, but effective studies. For example, we tested and found that GLP-1/GIP dual agonist was more effective than liraglutide in reducing inflammation and enhancing GDNF release in the MPTP mouse model of Parkinson’s disease [55]; triple GLP-1/GIP/Gcg receptor agonist reduced Aβ, neuroinflammation, and oxidative stress in the cortex and hippocampus [56]; GLP-1/GIP/Gcg triagonist reduced cognitive deficits and pathology in the 3xTg mouse model of AD [41]. Pathak et al. also reported that N-ac(D-Ala2)GIP/GLP-1-exe improved recognition memory, accompanied by enhanced hippocampal neurogenesis and synapse formation in high fat fed mice [57]. These studies using co- or tri-anonists are certainly useful for testing neuroprotective effects, but it is hard to clarify the respective contribution of different receptors, as well as the exact molecular mechanism. A main purpose of the present study is to explore the exact molecular mechanism of GIPR-mediated neuroprotective effects. So, we used the single receptor agonist DAla2GIP-Glu-PAL to specifically activate GIP receptors.
In conclusion, the present study demonstrated for the first time that chronic application of DAla2GIP-Glu-PAL could not only effectively improve multiple cognitive behaviors and synaptic plasticity of APP/PS1 mice, but also significantly reduce Aβ plaques and neuroinflammatory responses in the brain. The neuroprotective functions of DAla2GIP-Glu-PAL in improving cognition and reducing Aβ plaques in APP/PS1 mice are most likely associated with upregulation of the cAMPP/PKA/CREB pathway and inhibition of neuroinflammatory responses, suggesting that DAla2GIP-Glu-PAL might be potentially beneficial in the treatment of AD.
Footnotes
ACKNOWLEDGMENTS
This research was supported by the following programs: National Natural Science Foundation of China (31471080, 31700918, 31600865 and 81600951); Fund Program for “Sanjin Scholars” of Shanxi Pro-vince; Alzheimer Society and Alzheimer Drug Dis-covery Foundation (124660); Fund for Shanxi Key Subjects Construction, FSKSC, Shanxi “1331 Pro-ject” Key Subjects Construction (1331KSC) and Key Laboratory of Cellular Physiology (Shanxi Medical University) in Shanxi Province; Scientific and technological Innovation Project of Colleges and Universities in Shanxi Province (2017166); Chang-zhi Medical College Doctoral Initiation Fund (BS17001).