
Abstract
Background:
Impairment of synaptic plasticity along with the formation of amyloid-β (Aβ) plaques and tau-protein neurofibrillary tangles have been associated with Alzheimer’s disease (AD). Earlier studies with rat and mouse hippocampal slices have revealed the association of AD with the absence of synthesis of memory related proteins leading to impairment in cognitive functions. The role of hydrogen sulfide (H2S), a gaseous neurotransmitter, has been gaining attention as a neuroprotective agent. However, its role in AD-like conditions has not been studied so far.
Objective:
To study the neuroprotective role of H2S in AD conditions using rat hippocampal slices and the organic molecule GYY4137, a slow releasing H2S donor.
Methods:
Electrophysiological recordings were carried out in rat hippocampal slices to look into the impairment of LTP, a cellular correlate of memory. The Aβ42 peptide was bath-applied to mimic AD-like conditions and checked for both late-LTP and synaptic tagging and capture (STC) mechanisms of the synapses. GYY4137 was applied to look into its neuroprotective role at different stages during the recording of fEPSP.
Results:
There has been a steady decline in the plasticity properties of the synapses, in the form of late-LTP and STC, after the application of Aβ42 peptide in the hippocampal slices. However, application of GYY4137 rescued these conditions in vitro.
Conclusions:
GYY4137, with its slow release of H2S, could possibly act as a therapeutic agent in cognitive dysfunctions of the brain, mainly AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease which results in impairment of cognitive functions and is being considered as the most common cause of dementia and is becoming increasingly common especially with the world facing an aging population. 1 The formation of amyloid-β (Aβ) plaques and tau protein neurofibrillary tangles have been associated with AD, eventually resulting in neurodegeneration.2,3, 2,3 Additionally, hippocampal dependent episodic memory loss in AD condition has been found to be related to synaptic dysfunction.4–6
Synaptic plasticity, which is the change in the transmission efficacy of synapses, takes on different forms including long-term potentiation (LTP) which is widely believed to be the cellular corelate of learning and memory.7,8, 7,8 LTP is the enhancement of synaptic transmissions and can exist as short-lasting LTP (early-LTP), where there is no synthesis of protein and therefore degrades over time or long-lasting LTP (late-LTP) which is dependent on protein synthesis and can therefore persist for longer periods. 9 However, in the AD condition, the protein synthesis process is impaired, 10 preventing the expression of LTP which in turn affects cognitive function. 11 It is widely accepted that the formation and storage of memories depend on synaptic plasticity, which must occur at specific synapses and at precise times. 12 The synaptic tagging and capture (STC) hypothesis, which forms the basis of associative plasticity, states that proteins synthesized by a strongly-tetanized synaptic input can be utilized by another weakly tetanized independent input such that the early-LTP in the latter is transformed into late-LTP in a time dependent manner.13–17 To model AD in vitro, earlier studies applied Aβ1–42 peptide to hippocampal slices from mice and rats.18–20 Furthermore, late-LTP and STC are impaired in aged mice and also in mouse models with neurodegenerative diseases such as AD.11,21–23, 11,21–23 Therefore, finding ways to prevent or ameliorate such impairments is crucial in the search for new therapeutic options.
Recently, the role of various neurotransmitters in the progression of AD has been studied intensively. 3 Hydrogen sulfide (H2S), a gaseous neurotransmitter has been found to exhibit neuroprotective effects.24–27 Various studies have shown its beneficial roles in AD, such as reducing inflammation and apoptosis, and enhancing NMDA receptor function, which in turn promotes neuronal function and cognition.28–30 In clinical studies, AD patients exhibited a decreased level of S-adenosylmethionine, an activator of the H2S producing enzyme (Cystathionine β-synthase, CBS).5,31,32, 5,31,32 Therefore, based on its neuroprotective effects, H2S donors such as sulfide salts (Na2 S and NaHS) shows potential to be a promising therapeutic target.29,33, 29,33 The organic molecule GYY4137 is a slow releasing H2S donor, 34 with its effect ranging from vasorelaxant to anti-inflammatory to inhibition of tumor growth.32,34, 32,34 However, how GYY4137 can be used as an effective agent to prevent plasticity deficits in AD has yet to be studied. Thus, we were intrigued as to whether the exogenous release of H2S via GYY4137 in Aβ1–42 peptide-treated rat hippocampal slices could prevent plasticity deficits. Our results provide insights into the mechanisms of plasticity rescue by GYY4137 in an AD like condition.
MATERIALS AND METHODS
Slice preparation and incubation
A total of 112 slices from 50 male Wistar rats (5–7 weeks old), housed under 12/12-h light/dark cycle and supplied with food and water ad libitum, were used for the experiments in this study. All the procedures carried out were approved by the Institutional Animal Care and Use Committee (IACUC), National University of Singapore.
The rats were anaesthetized using CO2 and decapitated. The brains were then removed quickly and placed in 4°C artificial cerebrospinal fluid (aCSF), maintained. The aCSF, which is a modified Krebs-Ringer solution, consisted of the following (in mM): 124 NaCl, 3.7 KCl, 1.2 KH2PO4, 1 MgSO4.7H2O, 2.5 CaCl2, 24.6NaHCO3, and 10 D-glucose, saturated with 95% O2 and 5% CO2 (carbogen) at a rate of 16 L/h. The right hippocampus was then isolated and 400μm thick transverse slices were prepared using a manual tissue chopper (Stoelting, Wood Dale, IL, USA). These slices were then quickly transferred onto a nylon net fixed in an interface chamber (Scientific Systems Design, Ontario, Canada) and incubated at 32°C with a constant perfusion of carbogenated-aCSF at a constant flow rate of 1 mL/min. The slices were incubated for 3 h before starting the experiments.
Electrophysiology recordings
Two monopolar, lacquer coated stainless steel electrodes (5 MΩ; AM Systems, USA) were positioned at the stratum radiatum (sr) layer of the hippocampal CA1 region which contains the Schaffer collateral (SC) fibers. They were used to stimulate two independent synaptic inputs (S1 and S2) of a single neuronal population thus evoking field excitatory postsynaptic potentials (fEPSP). A third electrode was positioned in the apical dendritic layer between the two stimulating electrodes and was used as a recording electrode to record fEPSPs (Fig. 1A). The specificity of the pathways (S1 and S2) was tested using the method described in a previous study. 35 The signals obtained were amplified using a differential amplifier (Model 1700, AM Systems) and then digitalized via the CED 1401 analog-to-digital converter (Cambridge Electronic Design, Cambridge, UK). Signals were monitored online and recorded with a custom-made software, PWIN (Leibniz Institute for Neurobiology, Magdeburg, Germany). The software was also used to regulate stimulation.

Aβ1–42 application impairs late-LTP. A) Schematic representation of the two-pathway experimental paradigm showing the position of the electrodes in the CA1 region of the hippocampus. B) After a stable baseline of 30 min, late-LTP was induced in S1 (close circles) via STET which maintained throughout the recording period. The control potentials in S2 (open circles) remained stable throughout the recording period. C) After a stable baseline of 30 min, early-LTP was induced in S1 (close circles) via WTET which gradually declined to baseline level. The control potentials in S2 (open circles) remained stable throughout the recording period. D) Timeline representation of experiment protocol. E) After Aβ1–42 application for 3 h and a stable baseline of 30 min, late-LTP was induced in S1 (close circles) via STET which declined back to baseline level. The control potentials in S2 (open circles) remained stable throughout the recording period. F) After Aβ1–42 application for 3 h and a stable baseline of 30 min, early-LTP was induced in S1 (close circles) via WTET which declined back to baseline level. The control potentials in S2 (open circles) remained stable throughout the recording period. The STET induction protocol for late-LTP is represented by three solid arrows, while early-LTP induction by WTET is represented by a hatched single arrow. Representative fEPSP traces at –15 min (closed line), +30 min (hatched line) and +240 min (dotted line). Scale bar for all the traces vertical: 3 mV; horizontal 3 ms. Error bars indicate±SEM.
After the 3 h incubation period, an input-output curve (afferent stimulation versus fEPSP slope) was plotted prior to carrying out the experiments and the basal stimulation intensity was set at the value which evoked a fEPSP of 40% of the maximum slope value. For recording, four 0.2 Hz biphasic constant-current pulses with 0.1 ms/polarity were used at each time point. A stable 30 min baseline was recorded before the induction of long-term potentiation (LTP) or drug application. Early-LTP was induced using a weak tetanization (WTET) protocol consisting of a single stimulus train with a burst of 21 pulses at 100 Hz (pulse duration of 0.2 ms). 36 While late-LTP was induced using a strong tetanization (STET) protocol consisting of three stimulus trains, each with a burst of 100 pulses at 100 Hz (pulse duration of 0.2 ms with an intertrain interval of 10 min). More details about the preparation, incubation and electrophysiological procedures can be obtained from our previous publication. 36
Pharmacology
The Aβ1–42 peptide (AnaSpec, Fremont, CA, USA) oligomer was prepared 24 h before the start of experiment to allow for the oligomerization of the peptides. Briefly, Aβ1–42 peptide films were added to hexafluoroisopropanol (HFIP) and stored at –20°C. The peptide films were then dissolved in dimethyl sulfoxide (DMSO) and the DMEM/F-12 medium was added without phenol red and were stored at 4°C for 24 h. The final concentration of Aβ1–42 peptide used was 200 nM and was protected from light during application.20,37,38, 20,37,38 GYY4137 was dissolved in carbogenated aCSF at a concentration of 100μM. 34 Emetine dihydrochloride hydrate (Sigma-Aldrich) which was used as protein synthesis inhibitor, was stored as a stock solution of 20 mM in water. While the transcriptional inhibitor, anisomycin (Tocris Biosciences, Bristol, UK) was stored as a stock solution of 25 mM in DMSO. Emetine and anisomycin were diluted in aCSF to a working concentration of 20μM and 25μM respectively. For the stock solutions prepared in DMSO, the final DMSO concentration was maintained below 0.1%, which has been shown to not affect the basal responses of synapses. 39
Statistical analysis
All the data reported in this paper are represented as mean±SEM. For electrophysiology data, the average values of the slope function of the fEPSP (millivolts per milliseconds) were expressed as percentages of average baseline values. The values were analysed using the Wilcoxon signed rank test (Wilcox test) at specified time points for comparison within a group and the Mann-Whitney U-test (U-test) was used for comparison between groups. For western blot data, the Student’s t-test was used. p < 0.05 was considered to be statistically significantly different (*p < 0.05, **p < 0.001, and ***p < 0.0001). All statistical analyses were performed and graphs were prepared using the GraphPad Prism Software (San Diego, CA, USA, www.graphpad.com).36,37, 36,37
RESULTS
Exogenous application of Aβ1–42 impairs late-LTP but not early-LTP
Firstly, we investigated late-LTP and early-LTP in hippocampal CA1 pyramidal neurons. The two-pathway experimental paradigm depicted in Fig. 1A was used to study both plasticity processes. A stable baseline of 30 min was recorded by delivering low frequency test stimulation every 5 min in S1 and S2, followed either by STET or WTET in S1. STET resulted in a late-LTP in S1 which remained statistically significant for 240 min (Fig. 1B, close circles, Wilcox test, p = 0.0039; U-test, p = 0.0019, n = 9) and WTET resulted in an early-LTP which was statistically significant for 60 min (Fig. 1C, close circles, Wilcox test, p = 0.0156, n = 7) and eventually declined to baseline levels. The control potentials in S2 remained stable in both experiments throughout the entire recording period of 240 min (Fig. 1B, open circles; Fig. 1C, S2, open circles).
Next, the Aβ1–42 peptide was incubated with the hippocampal slices for 3 h, as a model system to mimic an AD like pathology (Fig. 1D). Late-LTP and early-LTP in these slices were then studied. Consistent with earlier reports,20,40, 20,40 late-LTP but not early-LTP maintenance was impaired in slices bath applied with Aβ1–42 peptide. STET induction resulted in a statistically significant potentiation in S1 which maintained for 180 min (Fig. 1E, close circles, Wilcox test, p = 0.0156, n = 8) then declined to baseline levels. WTET induction resulted in an early-LTP in S1 which declined to baseline levels (Fig. 1F, close circles, n = 5). Taken together, these data corroborate with the earlier reports20,40, 20,40 that Aβ1–42 peptide impairs long-term plasticity without affecting short-term plasticity.
GYY4137 rescues late-LTP in Aβ1–42 peptide treated hippocampal slices
We then examined the effect of GYY4137 (100μM) on the early and late plasticity in the CA1 region of the rat hippocampal slices. The baseline recordings in both S1 and S2 were maintained for 240 min (Fig. 2C, n = 5), similar to the control (Fig. 2B, n = 7). Interestingly, the addition of GYY4137 to the Aβ1–42 peptide treated slices rescued the deficits in late-LTP seen in Fig. 1E. STET induction resulted in a late-LTP in S1 which remained statistically significant for 4 h (Fig. 2D, close circles, Wilcox test, n = 0.0020; U-test, n = 0.0004, n = 10). The baseline stimulation in S2 was stable throughout the experiment showing that the basal neuronal properties were not altered by the application of Aβ1–42 peptide and GYY4137 (Fig. 2D, open circles).
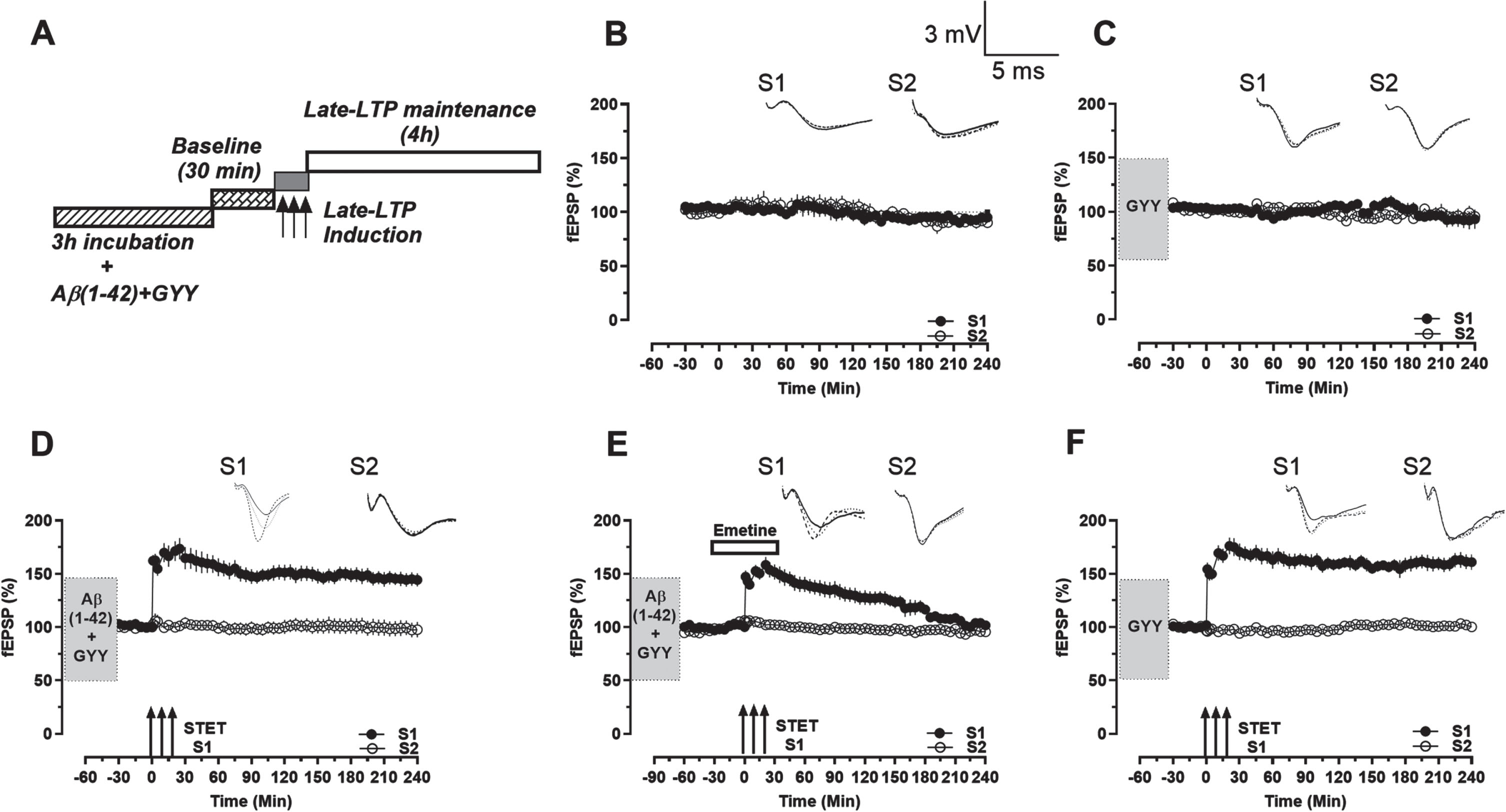
Rescue of late-LTP in Aβ1–42 treated hippocampal slices using GYY4137. A) Timeline representation of experiment protocol. B) Recording of stable baseline in S1 (close circles) and S2 (open circles) with low frequency stimulation. C) After GYY4137 application for 3 h, stable baseline in S1 (close circles) and S2 (open circles) with low frequency stimulation was recorded. D) After Aβ1–42 + GYY4137 application for 3 h and a stable baseline of 30 min, late-LTP was induced in S1 (close circles) via STET which maintained throughout the recording period. The control potentials in S2 (open circles) remained stable throughout the recording period. E) After Aβ1–42 + GYY4137 application for 3 h, emetine application for 1 h, from 30 min before STET to 30 min after and a stable baseline of 30 min, late-LTP was induced in S1 (close circles) via STET which declined to baseline level. The control potentials in S2 (open circles) remained stable throughout the recording period. F) After GYY4137 application for 3 h, and a stable baseline of 30 min, late-LTP was induced in S1 (close circles) via STET which maintained throughout the recording period. The control potentials in S2 (open circles) remained stable throughout the recording period. Symbols, traces, and scale bars are consistent with the figures presented earlier. Error bars indicate±SEM.
Next, to confirm the protein synthesis dependency of late-LTP maintenance, tetanization was carried out in the presence of the protein synthesis inhibitor Emetine (20μM) on GYY4137 and Aβ1–42 peptide treated slices. Emetine was bath applied for 1 h, from 30 min before STET to 30 min after. STET resulted in a potentiation in S1 which remained statistically significant till 180 min (Fig. 2E, close circles, Wilcox test, n = 0.0078; U-test, n = 0.0047, n = 8) and eventually returned to baseline levels. The baseline stimulation in S2 was stable throughout the experiment (Fig. 2E, open circles). Experiments were carried out to test whether GYY4137 has any effect on control late-LTP, which revealed no visible effect (Fig. 2F). STET induction resulted in a late-LTP in S1 which remained statistically significant for 240 min (Fig. 2F, close circles, Wilcox test, n = 0.0020; U-test, p < 0.0001, n = 9). The baseline in S2 remained relatively stable throughout the experiment (Fig. 2F, open circles).
GYY4137 rescues synaptic tagging and capture (STC) in Aβ1–42 peptide treated hippocampal slices
As GYY4137 is able to rescue deficits in late-LTP, we wanted to investigate the effect of GYY4137 on associative plasticity. A strong-before-weak (SBW) experimental paradigm was employed wherein a protein-synthesis dependent late-LTP was induced in S1 input while the protein-synthesis independent early-LTP was induced in S2 with an interval of 30 min between them.41,42, 41,42 Tagging experiments with control slices resulted in a late-LTP in S1 (Fig. 3A, close circles) after STET and the conversion of early-LTP to late-LTP in S2 after WTET (Fig. 3A, open circles) which remained statistically significant up to 240 min in both S1 and S2 (Wilcox test, n = 0.0078, n = 8). However, STC was impaired in slices treated with Aβ1–42 peptide (Fig. 3B). STET resulted in a potentiation which remained statistically significant till 90 min in S1 (Fig. 3B, close circles, Wilcox test, n = 0.0313, n = 6) and till 60 min in S2 (Fig. 3B, open circles, Wilcox test, n = 0.0313, n = 6).
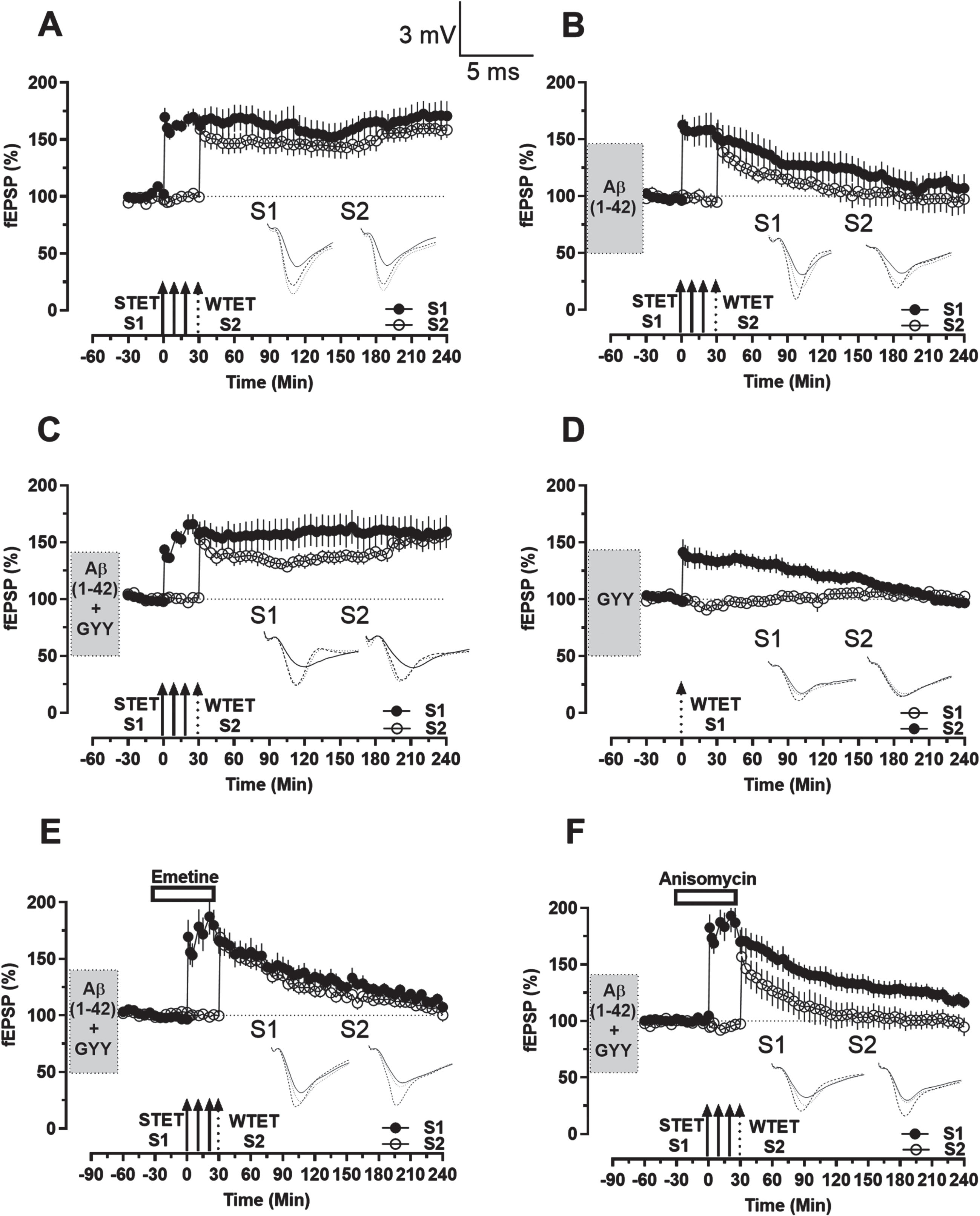
Rescue of STC in Aβ1–42 treated hippocampal slices using GYY4137. A) Induction of STC using the SBW paradigm. After a 30 min baseline, late-LTP was induced in S1 (close circles) via STET followed by the induction of early-LTP in S2 (open circles) via WTET after 30 min which resulted in late-LTP in both synaptic inputs that maintained throughout the recording period. B) After Aβ1–42 application for 3 h and a stable baseline of 30 min, using the same paradigm as A resulted in a potentiation that declined to baseline level in both synaptic inputs. C) After Aβ1–42 + GYY4137 application for 3 h and a stable baseline of 30 min, using the same paradigm as A resulted in late-LTP in both synaptic inputs that maintained throughout the recording period. D) After GYY4137 application for 3 h, and a stable baseline of 30 min, early-LTP was induced in S1 (close circles) via WTET which declined to baseline level. The control potentials in S2 (open circles) remained stable throughout the recording period. E, F) After Aβ1–42 + GYY4137 application for 3 h, emetine (E)/Anisomycin (F) application for 1 h, from 30 min before STET to 30 min after and a stable baseline of 30 min, using the same paradigm as A resulted in a potentiation that declined to baseline level in both synaptic inputs. Symbols, traces, and scale bars are consistent with the figures presented earlier. Error bars indicate±SEM.
We then investigated whether GYY4137 was able to rescue the tagging process in the Aβ1–42 treated slices. The induction of STET resulted in a late-LTP in S1 (Fig. 3C, close circles) that was able to transform the early-LTP produced by WTET induction in S2 into late-LTP (Fig. 3C, open circles), as it was observed that both S1 and S2 expressed statistically significant potentiation which remained till 240 min (Wilcox test, n = 0.0039 and Wilcox test, n = 0.0078 in S1 and S2 respectively, n = 9).
Additionally, WTET induction resulted in an early-LTP (Fig. 3D, close circles) in S1 which was statistically significant till 90 min (Wilcox test, n = 0.0313; U-test, n = 0.0087, n = 6). This shows that GYY4137 does not alter the short-term plasticity of the synapses, nor has the ability, of its own, to transform the short-term plasticity into long-term plasticity.
However, Emetine and Anisomycin application impaired the tagging process due to the inhibition of protein synthesis. STET in the presence of Emetine resulted in a potentiation that remained statistically significant till 180 min in both S1 (Fig. 3E, close circles, Wilcox test, n = 0.0391, n = 8) and S2 (Fig. 3E, open circles, Wilcox test, n = 0.0156 for S2, n = 8). STET in the presence of Anisomycin resulted in a potentiation that remained statistically significant till 50 min in S1 (Fig. 3F, close circles, Wilcox test, n = 0.0156, n = 7) and till 40 min in S2 (Fig. 3F, open circles, Wilcox test, n = 0.0156, n = 7).
Taken together, these data suggest that STC is impaired in hippocampal slices treated with Aβ1–42 peptide, and that GYY4137 is able to rescue the deficits in STC possibly through the synthesis of new PRPs. This is evident from the fact that the rescue effect is absent when protein synthesis is inhibited by Emetine and Anisomycin.
DISCUSSION
Our experiments indicate that exogenous application of Aβ1–42 oligomers in the hippocampal area CA1 impairs late-LTP but has no effect on early-LTP. More importantly, the impairment of late-LTP can be reversed by the addition of GYY4137 to the Aβ1–42 peptide treated slices. Our studies showing the protective effects of H2S donor in alleviating plasticity deficits are in line with other studies that showed that H2S attenuates spatial memory impairment and hippocampal neuroinflammation in β-amyloid rat model of AD. 30 H2S is known to prevent neuronal and synaptic deterioration and improve deficits in memory, cognition, and learning. 43 It is being proposed that H2S can modulate the activity of the NMDA receptors through the cAMP pathway which activates the CREB. 44 The rescued LTP during the application of the Aβ peptide by GYY4137 may also be due to a similar mechanism. GYY4137 also might mediate synthesis of plasticity related proteins as emetine blocked the GYY4137 mediated rescue of LTP in Aβ treated slices. This is supported by findings that H2S facilitates LTP and regulates intracellular calcium levels, which are critical for learning and memory. 43
H2S activates the TrKB – mTOR-AMPA signaling pathways to exert antidepressant effects that are indirectly associated with synaptic protein synthesis or restoration of synaptic plasticity in major depressive disorders, therefore activating TrkB signalling could be another mechanism by which H2S confers neuroprotection. 45
Our experiments investigating STC on slices treated with Aβ peptide did not yield any tagging as compared to the control, providing evidence for the lack of associative memory in the AD condition as reported in earlier studies.46,47, 46,47 This may be due to impairments in either tag setting or protein synthesis. The dependence of protein synthesis in tagging process was further confirmed by performing the tagging experiments in the presence of protein synthesis inhibitors. Both Emetine and Anisomycin inhibited the STC process, confirming its role in maintaining the tagging process. Interestingly, our experiments with GYY4137 on Aβ treated slices found that H2S is able to rescue the associative properties of the neurons.
Aberrant metabolism of H2S are implicated in the development of neurodegenerative diseases such as AD and Parkinson’s disease. 43 Moreover, various H2S donors have exhibited beneficial therapeutic effects in models of these diseases by targeting key pathological features, such as Aβ production in AD and neuroinflammation in Parkinson’s disease, which also are also in line with our studies showing H2S donor rescues synaptic deficits in an Aβ model. It might rescue synaptic deficits by preventing neuronal damage or due to its anti-inflammatory effect as numerous in vivo studies have shown that H2S not only prevents neuronal and synaptic damage but also improves memory, cognition, and learning deficits.48–50 The neuroprotective properties of H2S might be primarily due to its anti-inflammatory or antioxidant effects.
The present study shows that the H2S could be a potential therapeutic agent in the treatment of various neurodegenerative diseases such as AD. However, further studies are required in vivo, specifically behavioral studies to confirm if there is an improvement in cognitive functions with the use of GYY4137. Furthermore, a detailed biochemical analysis to decipher the molecules that are regulated by H2S can provide insights into the pathways involved which can in turn reveal novel therapeutic targets for AD.
AUTHOR CONTRIBUTIONS
Anoop Manakkadan (Data curation; Formal analysis; Investigation; Writing – original draft); Dolly Krishnan (Data curation); Sheila Rui Xia Ang (Data curation; Formal analysis; Writing – review & editing); Sreedharan Sajikumar (Conceptualization; Funding acquisition; Supervision; Validation; Writing – review & editing).
Footnotes
ACKNOWLEDGMENTS
We thank Prof. Philip Keith Moore, Department of Pharmacology, National University of Singapore, for providing GYY4137.
FUNDING
This work was supported by the Ministry of Education Academic Research Fund Tier 3 (MOE2017-T3-1-002), NUHS seed fund (NUHSRO/2020/145/RO5 + 6/Seed-Sep/05) and Ministry of Health (MOH-000641-00), Singapore National Research Foundation (NRF2019-NRF-ISF003-3141) to S.S.
CONFLICT OF INTEREST
Sreedharan Sajikumar is an Associate Editor of this journal but was not involved in the peer-review process of this article nor had access to any information regarding its peer-review.
DATA AVAILABILITY
The data supporting the findings of this study are available within the article.