
Abstract
Background:
Cromolyn is an anti-neuroinflammatory modulator with a multifactorial mechanism of action that has been shown to inhibit amyloid-β (Aβ) aggregation and enhance microglial uptake and clearance of Aβ.
Objective:
We report the effects of fluoro-cromolyn derivatives on microglial cell toxicity and microglial clearance of Aβ42.
Methods:
Microglial cell toxicity for cromolyn derivatives were determined in naive BV2 microglial cells. Microglial clearance assays were performed with Aβ42 in naive BV2 microglial cell line and single cell clone BV2 line expressing CD33WT. PET imaging was performed for three F-18 analogs in a rhesus macaque.
Results:
All compounds but derivative
Conclusion:
Substantial uptake of Aβ42 in both naïve BV2 and BV2-CD33WT cells observed with
INTRODUCTION
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder that afflicts over 35 million people throughout the world, including 5.8 million Americans age 65 and older [1]. Currently there is no effective disease-modifying therapy available to slow or stop the destruction of neurons that cause AD symptoms [2]. Present biochemical and pathological evidence support the concept that an imbalance between production and clearance of soluble amylo-id-β (Aβ42) resulting in abnormal accumulation of Aβ oligomers are responsible for the synaptotoxic effects that lead to neuronal stress, abnormal tau phosphorylation, synapse collapse, and memory im-pairment [3, 4]. Soluble Aβ monomers aggregate to form toxic higher order oligomers (dimers, trimers, up to dodecamers), which readily lead to extracellular deposition of fibrillar amyloid neuritic plaques that are resistant to degradation. Aβ oligomer accumulation drives tauopathy and neurodegeneration [5], all which combine to activate a microglial-driven neu-roinflammatory response. Microglia are critical for Aβ phagocytosis; however, chronic inflammatory activation compromises microglial clearance of Aβ contributing to disease progression prior to clinical symptoms [6, 7]. Hence, interfering with the assembly of Aβ peptides from monomer to oligomeric spe-cies and fibrils, or promoting their clearance from the brain, are targets of anti-Aβ directed therapies in AD [8–10].
Microglia are the primary immune cells of the central nervous system (CNS) that play an important role in removal of cellular debris and aberrant protein debris. Microglial cells respond to the presence of AD pathological lesions (plaques and tangles) by changing their morphological characteristics, expre-ssing numerous cell surface receptors, and surroun-ding the lesions [11–13]. Microglial cells can assume activation states in a spectrum ranging from phagocy-tic/neuroprotective to pro-neuroinflammatory. Under normal physiological conditions, microglia secrete anti-inflammatory cytokines and supportive growth factors for neuroprotection and neuro repair, while also clearing proteinaceous and cellular debris. Under pathological conditions, such as amyloid plaque acc-umulation, tauopathy and neurodegeneration in the AD brain, classically-activated microglia release to-xic pro-inflammatory cytokines that induce gliosis dramatically amplify neurodegeneration. This pro-cess leads to more neuronal cell death and debris and accelerates disease progression. Research eff-orts have focused on converting microglia from the pro-neuroinflammatory activation state to the neuro-protective/anti-neuroinflammatory activation state in which the toxic effects are reduced and phagocytic activity toward Aβ is enhanced [14–17]. This is a potential avenue for new therapeutic approaches to treating and preventing AD.
Cromolyn (
Cromolyn compounds
Numerous studies indicate the involvement of chronic neuroinflammation in the pathogenesis of AD [2, 27]. The sustained activation of microglia and other immune cells in the brain release a variety of proinflammatory and toxic products, including reactive oxygen species, nitric oxide, and cytokines which exacerbate both amyloid and tau pathology. We have been investigating the potential utility of cromolyn as a therapeutic for slowing down AD progression and have reported that cromolyn inhibits Aβ aggregation, enhances microglial recruitment to plaques in transgenic AD mice, and promotes phagocytosis and clearance of Aβ [8, 28]. Cromolyn was found to efficiently inhibit the aggregation of Aβ monomers into higher order oligomers and fibrils in vitro, without affecting Aβ production; in vivo, the levels of soluble Aβ were decreased by over 50%after only 1 week of daily intraperitoneally administered cromolyn in APP/PS1 AD mice [8]. This reduction in Aβ levels was largely attributed to the ability of cromolyn to promote microglial recruitment to Aβ plaques and to induce microglial phagocytosis of Aβ [28].
In a study of the effects of cromolyn on a the SOD1 G93A transgenic mouse model of familial amyo-trophic lateral sclerosis, cromolyn treatment significantly delayed the onset of neurological symptoms, increased motor neuron survival in the lumbar spinal cord, and decreased the expression of pro-inflam-matory cytokines/chemokines in the lumbar spinal cord and plasma. Together, these findings suggest that cromolyn sodium provides neuroprotection by decreasing neuroinflammatory response to disease pathology [29]. These neuroprotective mechanisms strongly suggest that cromolyn acts directly in the brain. To better understand the mechanism of action and to develop derivatives with enhanced brain penetration several fluoro-cromolyn derivatives have been investigated. Here, we report on the effect of fluoro-cromolyn derivatives on Aβ uptake by microglial cells, as well as microglial cell toxicity. We also describe the synthesis of F-18 labeled cromolyn analogs and their PET imaging pharmacokinetics in nonhuman primate brain.
MATERIALS AND METHODS
All experiments involving nonhuman primates were performed in accordance with the U.S. Department of Agriculture (USDA) Animal Welfare Act and Animal Welfare Regulations (Animal Care Blue Bo-ok), Code of Federal Regulations (CFR), Title 9, Cha-pter 1, Subchapter A, Part 2, Subpart C, §2.31. 2017. Experiments were approved by the Animal Care and Use Committee at the Massachusetts General Hos-pital. The animal used in this study was an adult male Rhesus macaque (13 years old). Prior to each study, animals were sedated with ketamine/xylazine (10/0.5 mg/kg IM) and were intubated for maintenance anesthesia with isoflurane (1-2%in 100%O2). A venous catheter was placed for infusion of the radiotracer and, where applicable, an arterial catheter was placed for sampling of the arterial input function. The animal was then positioned on a heating pad on the bed of the scanner for the duration of the study.
A GE PETtrace 16.5 MeV cyclotron (GE Healthcare, Waukesha, WI, USA) was used for [18F]fluoride production by the 18O(p,n)18F nuclear reaction to irradiate 18O-enriched water (Isoflex Isotope, San Francisco, CA). Dynamic PET sinograms were performed on a GE Discovery MI (GE Healthcare) PET/CT scanner.
Chemistry
Hydroxy-cromolyn
Preparation of cromolyn derivatives for microglial cell-based assays
Cromolyn derivatives,
Generation of the single cell clone BV2 microglial line stably expressing human CD33 (BV2-CD33WT)
To generate a single cell clone BV2 line stably expressing human CD33, a well of a 6-well-plate containing naive BV2 cells were transiently transfected with the pcDNA3.1 plasmid encoding human wild-type CD33 (CD33WT) using Lipofectamine-Plus (Life Technologies), according to the manufacturer’s instructions. The following day the media was cha-nged. Two days post transfection, we started the process of isolating a clonal population of BV2 cells st-ably expressing CD33WT. For this purpose, cells were trypsinized and centrifuged. Afterwards, cells were washed with phosphate buffer saline (PBS) and centrifuged. The cell pellet was resuspended in FACS sorting buffer that consists of 2%heat-inactivated fetal bovine serum, 2%B27 supplement (Gibco) in PBS. Subsequently, cells were incubated in FACS sorting buffer containing Fc block (1μg/ml, anti-mouse CD16/32, clone 93, Biolegend) for 10 min on ice. Afterwards, cells were labeled with either anti-CD33 antibody-FITC (mouse IgG1, clone HIM3-4, BD Pharmingen) or IgG1-FITC (BD Pharmingen) as control for 30 min on ice. Cells were rinsed with FACS sorting buffer and centrifuged. Then, cells were gently resuspended in FACS sorting buffer and filtered into polystyrene filter top tubes (BD Falcon) for FACS sorting.
BV2 cells expressing CD33WT were visualized based on CD33 expression, using FACS ARIA (BD Biosciences). The anti-CD33 antibody-FITC bound to CD33 expressed on the cell surface, while cells treated with the isotype antibody IgG1 served as control. Single cells were sorted based on the expression levels of CD33 detected in the FITC channel, using FACS ARIA. Single cells were sorted into 96-well plates containing cell growth media: DMEM (Lonza), 10%heat-inactivated fetal bovine serum, 2 mM L-glutamine, 1%penicillin/streptomycin, and geneticin at 400μg/ml (G418 sulfate, Life Technologies). The pcDNA3.1-CD33WT vector contains the neomycin resistance gene for selection of stable cell lines using geneticin. After one week, half of the cell growth media was replaced with fresh media in each well every three days. After an additional week, each single cell clone was trypsinized and transferred into one well in 24-well plates for cell expansion. Upon reaching confluency, each single cell clone was trypsinized and plated into two wells in 6-well plates. Once confluent, cells from one well were frozen in heat-inactivated fetal bovine serum containing 10%dimethyl sulfoxide (Sigma Aldrich), while cells in the second well were lysed in RIPA lysis buffer (EMD Millipore) supplemented with protease inhibitors (Roche) and phosphatase inhibitors (Thermo Fisher Scientific). We obtained several cell clones that we screened for CD33 expression by using western blotting and anti-human CD33 antibody (mouse monoclonal, clone PWS44, Leica Biosystems). Finally, the single cell clone line expressing CD33WT (BV2-CD33WT) was identified and confirmed by western blotting. The corresponding frozen cell vial was rapidly thawed and plated in cell growth media in a 10 cm culture dish to allow cell expansion.
Cromolyn compounds microglial toxicity and Aβ uptake assays in microglia

Radiosynthesis of
Naive BV2 microglial cell line or single cell clone BV2 line stably expressing CD33WT (BV2-CD33WT) were plated in 12-well plates at the density of 5x10E5 cells for naïve BV2 and 7x10E5 cells for the BV2-CD33WT cell clone, in proliferating media: DMEM containing 10%heat-inactivated fetal bovine serum, 2 mM L-Glutamine and 1%penicillin/streptomycin (Life Technologies). On the following day, cells were treated with DMSO (control) or cromolyn derivatives at different concentrations in proliferating media for 3 h.
PET imaging
A CT scan was acquired for attenuation correction of the PET images. Emission PET data collection was initiated after bolus injection of F-18 cromolyn derivatives (207.2 MBq) into the lateral saphenous vein of a rhesus macaque followed by a saline flush. Dynamic PET imaging was performed for up to 180 min with arterial blood sampling. Total volume of distribution (vt) was estimated using compartmental modeling (one- and two-tissue configurations) and graphical analysis techniques (Logan regression methods). Time bins were 6×10, 8×15, 6×30, 8×60, 8×120, and 30×300 s for 18F-TS274 and 6×10, 8×15, 6×30, 8×60, 8×120, and 18×300 s. Imaging started at the same time as bolus injection. Dynamic PET sinograms were reconstructed using the VPFXS reconstruction with 34 subsets and 3 iterations. Final reconstructed images had voxel dimensions of 256×256×89 and voxel sizes of 1.7×1.7×2.8 mm. The GE discovery MI scanner used for these studies has a spatial resolution around 5 mm at the center of the field of view. Dynamic PET sinograms were reconstructed using the VPFXS reconstruction with 34 subsets and 3 iterations. Final reconstructed images had voxel dimensions of 256×256×89 and voxel sizes of 1.7×1.7×2.8 mm. The GE discovery MI scanner used for these studies has a spatial resolution around 5 mm at the center of the field of view.
Whole blood and plasma time-activity curves were obtained by linear interpolation of radioactivity concentrations (kBq/cc) measured from the arterial blood samples. Blood samples were drawn at different time points during the entire experiment. Arterial samples of 1–3 mL were acquired every 30 s immediately following radiotracer injection during the first 5 min and decreased in frequency to every minute, 2 min, 5 min, 15 min, and 30 min toward the end of the scanning duration. Metabolite analysis was performed on standard and the plasma samples drawn at 5, 10, 15, 30, 60, 90, 120, and 180 min. These samples were assayed with column-switching radio-HPLC to determine the fraction of total radioactivity that was attributable to intact radiotracer. Plasma protein binding was measured in blood drawn immediately prior to tracer injection.
Radiofluorination
5, 5’- [(2-[18F]Fluoro- 1, 3-propanediyl) bis(oxy)] bis[4- oxo-4H- 1- benzopyran- 2- carboxylic acid diethyl ester] (4a)
A [18F]fluoride solution (50 mCi) was trapped onto a QMA Sep-Pak Cartridge (Sep-Pak plus light, Waters, Milford, MA) and then released by a solution of Kryptofix-2.2.2. (6 mg) and potassium carbonate (2 mg) in acetonitrile and water (1 mL, v/v 7:3) into a sealed Wheaton 5 mL reaction vial. The [18F] flu-oride solution was azeotropically dried with the aid of nitrogen gas at 120°C for 10 min. The K18F/Kry-ptofix complex was dried three times at 120°C by the addition of 1 mL of acetonitrile followed by evapo-ration of the solvent using a nitrogen flow. A solution of 1,3-bis[tolylsulfonyl)oxy]-2-[(trifluoromethyl)-sulfonyl]oxy-propane (4 mg,
5, 5’- [(2-[18F]Fluoro- 1, 3- propanediyl) bis(oxy)] bis[4- oxo-4H- 1- benzopyran- 2- carboxylic acid] (2a)
Following the procedure for the synthesis of
5, 5’-(2-[18F] Fluoro- 1, 3- propanediyl) bis(oxy)] bis[4- oxo-4H- 1- benzopyran- 2- carboxylic acid di-tert-butyl ester] (10)
A sealed Wheaton 5 mL reaction vial containing 50 mCi of fluorine–18 in 1 mL of 18O-enriched water, Kryptofix-2.2.2. (6 mg), and potassium carbonate (2 mg) was heated at 120°C and solvent was evaporated with the aid of nitrogen gas. The K18F/Kryptofix complex was dried three times at 120°C by the addition of 1 mL of acetonitrile followed by evaporation of the solvent using a nitrogen flow. A solution of the tosylate cromolyn t-butyl diester
RESULTS
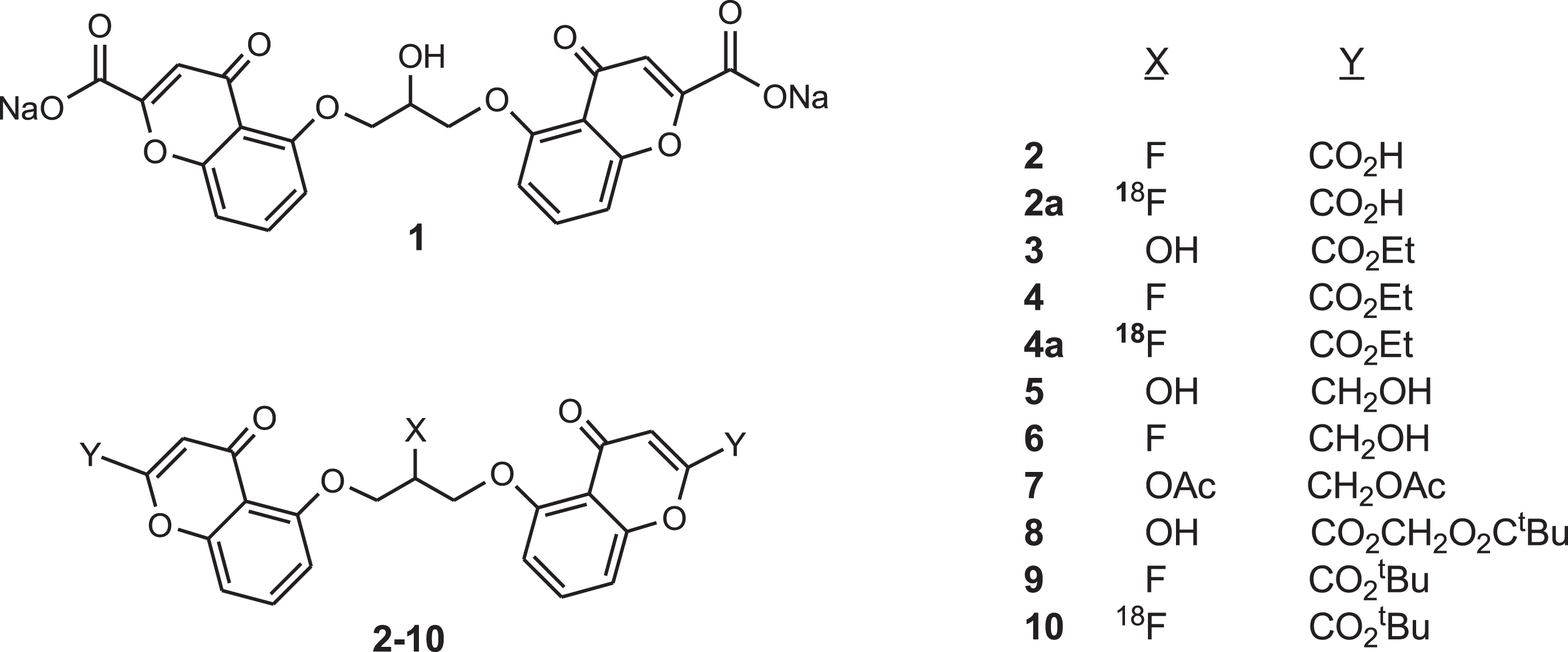
Cromolyn microglial cell toxicity
Naive BV2 microglial cells were treated with DMSO (control) or hydroxy-cromolyn
Microglial toxicity of cromolyn derivatives compared to DMSO.
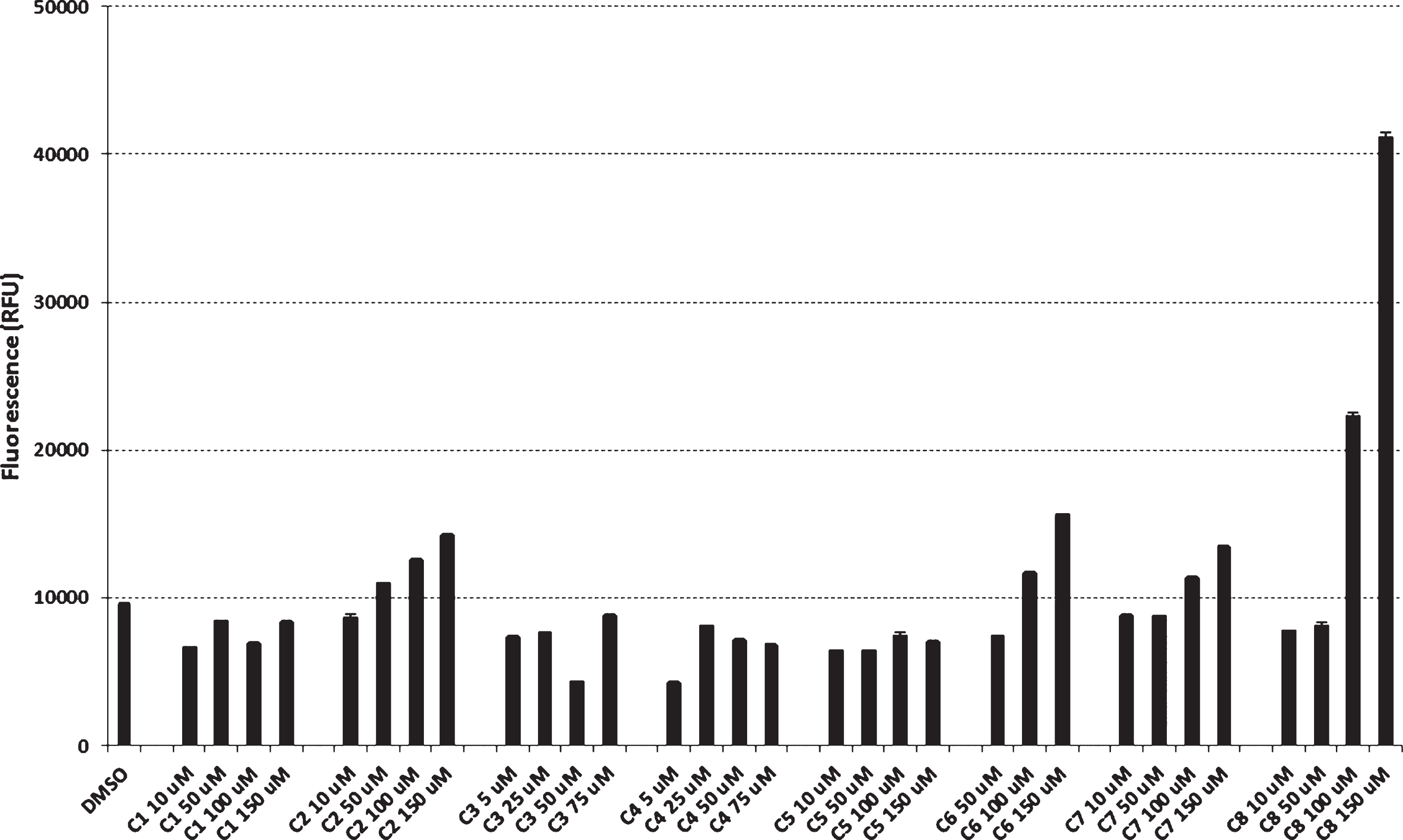
Effect of cromolyn derivatives on Aβ42 uptake in microglial BV2-CD33WT activated cells.
Cromolyn derivative effect on microglial uptake of Aβ42
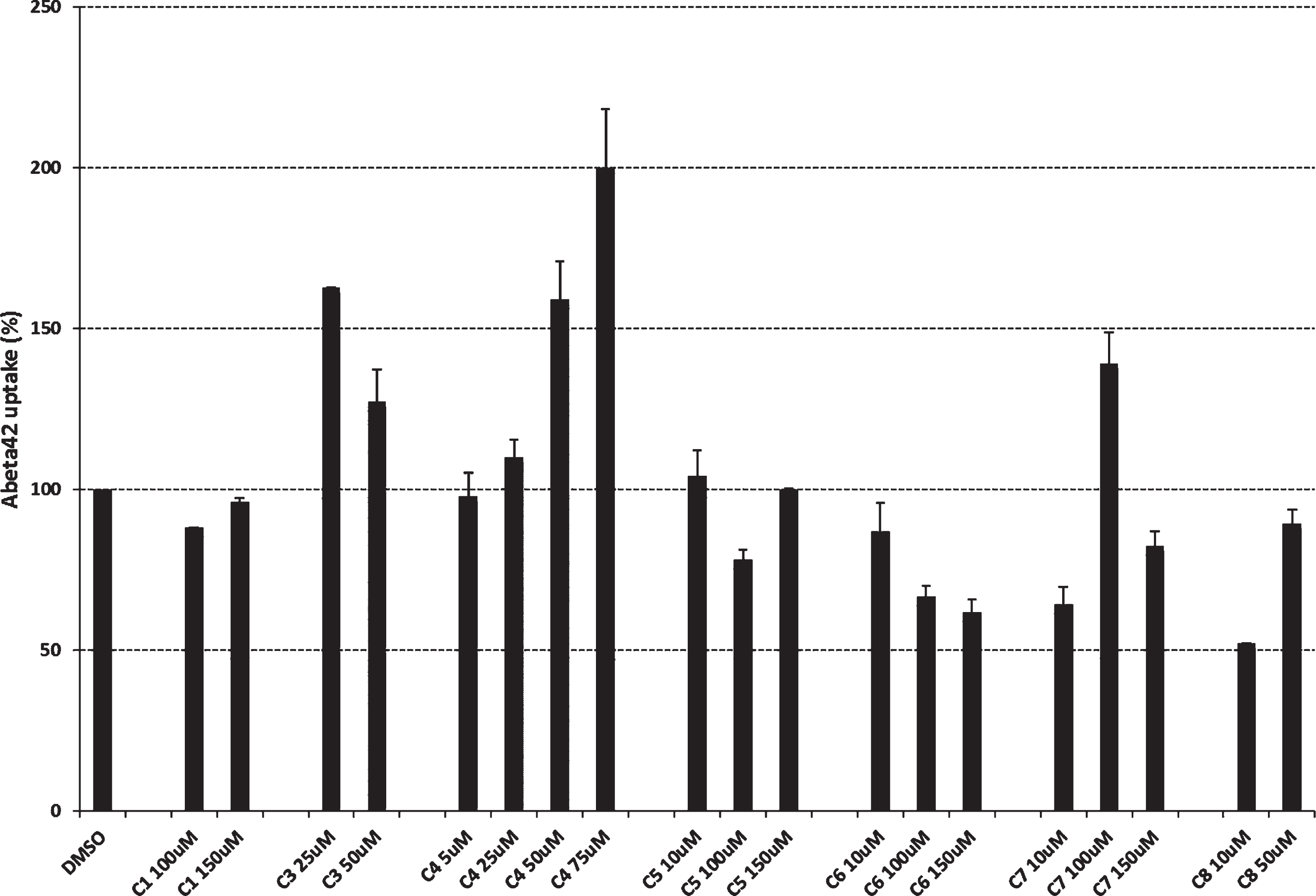
To explore the effects of cromolyn derivatives on Aβ42 uptake, we employed a microglial cell-based assay. Specifically, naïve BV2 microglial cell cultures were treated with cromolyn derivatives
Effect of cromolyn derivatives on Aβ42 uptake in naïve BV2 microglial cells.
We previously showed that the AD risk factor CD33 inhibits uptake and clearance of Aβ42 in mic-roglial cell cultures, a process that requires the sia-lic acid-binding V-Ig domain of CD33 [31]. Here, we generated a single cell clone BV2 line stably expressing human wild-type CD33 (BV2-CD33WT). The effect of cromolyn derivatives on microglial uptake and clearance of Aβ42 was investigated in the single cell clone BV2-CD33WT microglial line. BV2-CD33WT cells were treated with DMSO (control) or cromolyn derivatives at different concentrations ranging between 5 and 150μM. Results for compounds
Radiofluorination
Radiosynthesis of cromolyn
PET imaging
The aim of this PET study was to measure brain exposure and regional distribution of selected F-18 labeled cromolyn analogs administered intravenously in a monkey with an intact BBB. Pharmacokinetics of three F-18 analogs (diethyl ester
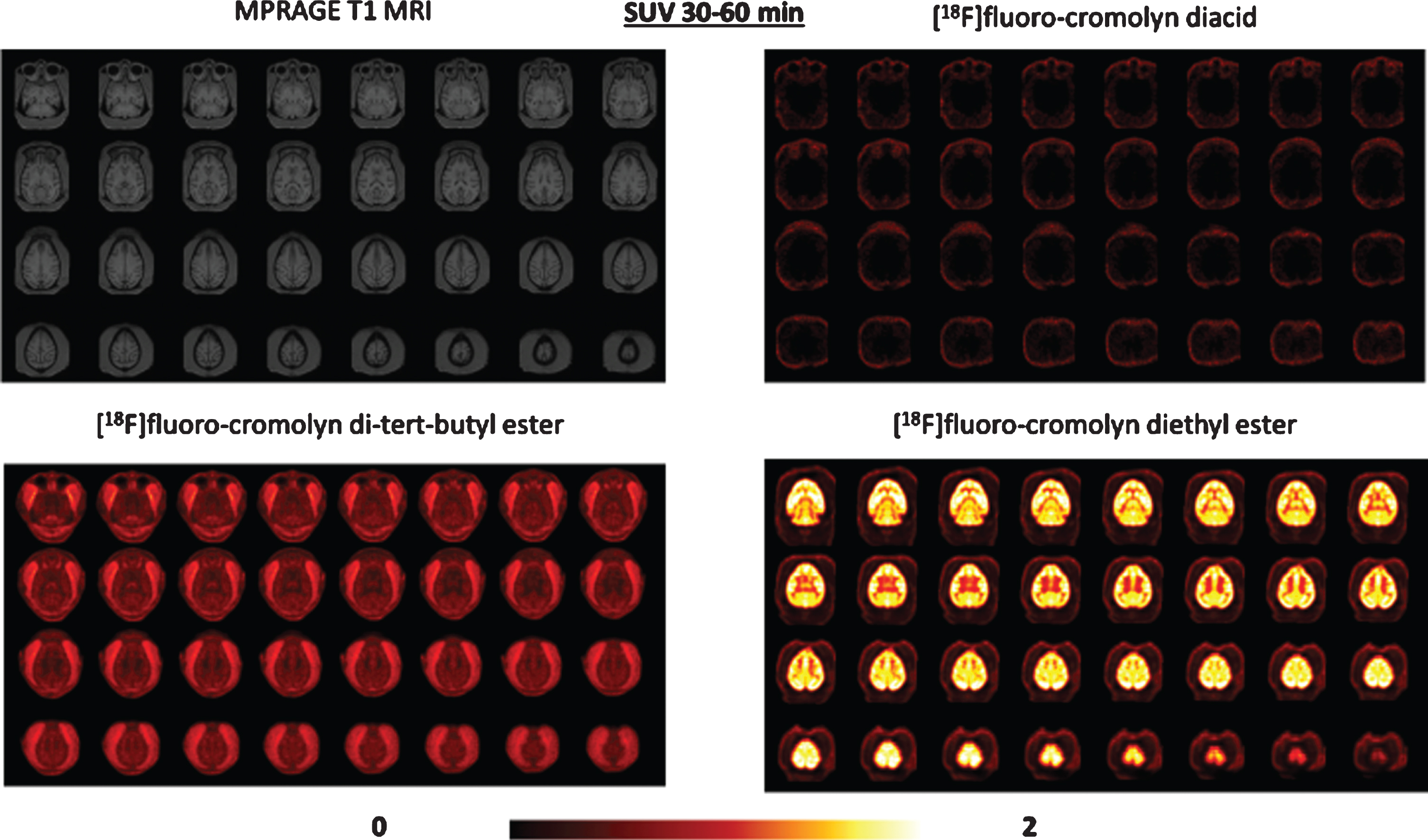
PET imaging data comparison of [18F]fluoro-cromolyn diacid
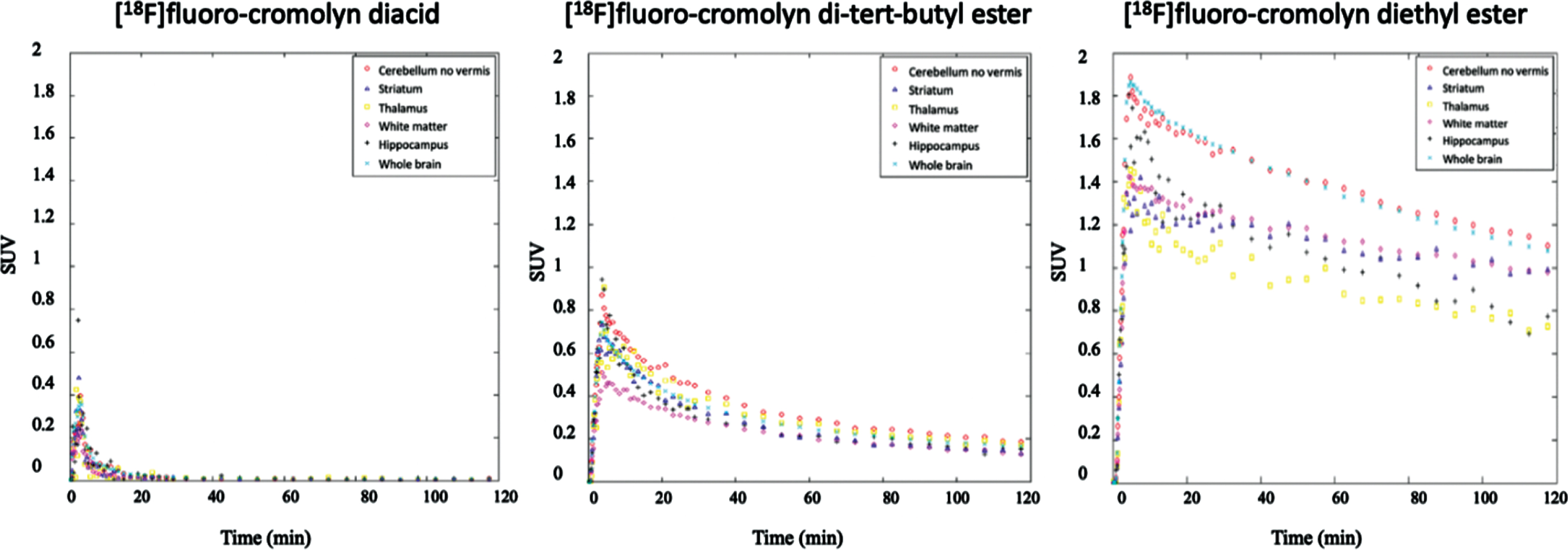
Time-activity curves from PET imaging data of [18F]fluoro-cromolyn diacid
PET images obtained with [18F]fluoro-cromolyn diethyl ester
DISCUSSION
Our results indicate that fluoro-cromolyn derivatives represent potential therapeutic agents for treating and preventing AD by reducing pro-neuro-inflammatory activation of microglia, enhancing mic-roglial phagocytosis and promoting clearance of Aβ. Several fluoro-cromolyn and non-fluoro-cromolyn derivatives were investigated in this initial screening to better understand the structure-activity relationship of cromolyn to microglial uptake and clearance of Aβ42 and to develop derivatives with enhanced brain penetration. Replacement of OH in cromolyn by F and esterification of the acid or reduction to the alcohol are expected to result in enhanced lipophilicity and thus greater brain permeable compounds. Moreover, fluorine derivatives offer the added advantage of monitoring brain penetration by labeling with flourine-18 and using PET imaging. Additionally, cromolyn derivatives containing labile substituents such as ethyl ester, t-butyl ester, acetate or methyl pivalate ester could act as lipophilic transient derivatives or prodrugs to deliver cromolyn
PET imaging with three [18F]fluoro-cromolyn derivatives revealed the order of brain tracer penetr-ation to be diethyl ester
In summary, these results indicate that fluoro-cromolyn derivatives represent potential therapeutic agents for treating and preventing AD by reducing pro-neuroinflammatory activation of microglia and simultaneously promoting clearance of Aβ via microglial phagocytosis.