
Abstract
Background:
Studies have reported that apolipoprotein E epsilon 4 (APOE ɛ4) has adverse effects on executive functions (EFs) in late adulthood. However, the results have been inconsistent. Insufficient measurements of executive functioning, uncontrolled clinical and demographic confounders, and moderation effects from other environmental factors are suspected to account for the inconsistency.
Objective:
This study used aggregate measures to examine the effects of APOE ɛ4 on four components of EFs, namely switching, working memory, inhibition, and reasoning. We further investigated whether high educational attainment, a proxy measure for cognitive reserve, moderates the adverse effects of ɛ4 on EFs.
Methods:
Cognitively unimpaired older participants were divided into groups based on APOE genotype and into subgroups based on educational attainment level. The demographic and clinical variables were matched between the groups. Four core components of the EFs were measured using a relatively comprehensive battery.
Results:
The results revealed that although no main effect of the APOE genotype was observed across the four EF components, the potentially adverse effects of ɛ4 on inhibition were alleviated by high educational attainment. A main effect of education on the reasoning component was also observed. The moderation analysis revealed that for older adults with 12 years of education or fewer, the relationship between the APOE ɛ4 genotype and inhibition performance became increasingly negative.
Conclusion:
This study highlights the distinctive role of response inhibition in the gene–environment interaction and underlines the importance of considering factors of both nature and nurture to understand the complex process of cognitive aging.
INTRODUCTION
Identifying risk factors and protective factors that explain individual differences in age-related cognitive decline is among the greatest challenges facing health care for older adults. The apolipoprotein E (APOE) ɛ4 allele is a well-established risk factor for dementia. Meta-analyses and several large-scale studies have indicated that cognitively unimpaired carriers of the APOE ɛ4 allele tend to have poorer performance on specific cognitive functions. However, the effects tend to be small in magnitude and have been inconsistent across studies [1–5]. For example, studies have reported reduced performance in several executive functions (EFs), including shifting [6, 7], working memory [8], and abstract reasoning ability [9] in nondemented older ɛ4 carriers compared with ɛ4 noncarriers. Other studies have not observed negative effects on EFs in ɛ4 carriers [10–12]. The mixed research findings may be related to the large variations in test selection and subcomponents of EF targeted by studies investigating the effects of the ɛ4 allele on EFs. EFs are complex higher cortical functions that comprise multiple subcomponents [13–15]. However, most of these studies have only employed one or very few measures of select EFs, which hinders the ability to compare the effects of the ɛ4 genotype on EFs across different studies.
Interest in investigating gene–environment interactions between APOE ɛ4 status and education on cognitive function and brain structure in late life has grown because higher educational attainment may enhance cognitive reserve, which protects against age-related cognitive decline [16–18]. However, results regarding the moderating effect of high educational attainment on ɛ4-associated cognitive disadvantages remain inconsistent. Several studies have demonstrated that high educational attainment reduces the negative effects of APOE ɛ4 on dementia incidence [19], general cognition [20–22], episodic memory and phonemic fluency [20], and a composite cognitive measure (letter fluency, similarity test, and word recall tests) [23]. However, other studies have reported either no association between educational attainment and cognitive decline in ɛ4 carriers [24] or a reverse relationship, in which ɛ4 carriers with a higher educational level displayed a more pronounced cognitive decline than education-matched noncarriers in episodic memory [25] or shifting ability [26], whereas carriers with a lower educational level did not. The cause of the inconsistency between studies remains unclear and may reflect differences in study samples or empirical methodologies.
Although evidence indicates a possible interaction effect between APOE ɛ4 and educational attainment on cognition, the studies reporting this evidence have certain drawbacks. For example, most of the studies only administered the Mini-Mental State Examination (MMSE)—a screening test for global cognition—or limited cognitive tests focusing on episodic memory or language. No studies have examined whether this interaction affects different EF components. EFs are of interest because they are commonly affected by education [17] and are reported to be associated with the APOE ɛ4 allele [1, 2]. Moreover, some studies [19–22] did not include clinical variables (e.g., vascular burden and emotional status), which may confound the findings regarding cognitive performance among participants. One study [20] did not control for demographic variables (e.g., age and educational level) for subgroup comparisons.
The present study addressed the aforementioned research gaps to investigate the interaction effect between level of educational attainment and APOE genotyping status on cognition, with an emphasis on EF, in cognitively unimpaired older adults. The aim of the present study was twofold. First, we examined the potential negative effect of the APOE ɛ4 allele on four EF components, namely abstract shifting, working memory, inhibition, and reasoning ability in nondemented older adults. The cognitive effect was expected to be subtle and inconsistently observed with different tests, and at least two measures were used for each of the EF components, based on the assumption that more assessment may yield a more reliable estimate of actual cognitive changes. We hypothesized that ɛ4 noncarriers would perform better than ɛ4 carriers on all four EF components. Second, we investigated whether an interaction effect could be observed between the APOE genotype and educational attainment. We hypothesized that higher educational attainment could reduce the negative effect of the APOE ɛ4 allele on the four components of EFs by enhancing cognitive reserve.
MATERIALS AND METHODS
Participants
This study recruited 68 community-dwelling ol-der adults without dementia, among whom 31 had at least one APOE ɛ4 allele (ɛ4+; homozygote, n = 2; heterozygote, n = 29), and 37 age- and education-matched non-ɛ4 (ɛ4–) cognitively unimpaired older adults. Potential participants were thoroughly screened through interviews to exclude individuals with a history of neurological or psychiatric disorder diagnoses, head injury with loss of consciousness, or alcohol or substance abuse. In addition, all participants were screened for mild cognitive impairment (MCI) through a battery of standardized neuropsychological tests, and only participants with an MMSE score of≥26, a global rating of 0 on the Clinical Dementia Rating (CDR) scale, and typical age-appropriate performance on the memory, attention span, visuospatial ability, and processing speed tests (see below) were included. The ɛ4 + and ɛ4–groups were further divided into subgroups based on the number of years of education completed: 1) the high cognitive reserve (HCR) group comprised individuals with more than 12 years of completed education (i.e., a high school diploma); 2) the low cognitive reserve (LCR) group comprised individuals with 12 years or fewer of completed education. Twenty participants were included in the ɛ4 + HCR group, 11 in the ɛ4 + LCR group, 22 in the ɛ4–HCR group, and 15 in the ɛ4–LCR group. The present study was approved by the Ethics Committee and Institutional Review Board at National Taiwan University Hospital and followed the guidelines established by the Declaration of Helsinki. Written informed consent was obtained from all participants before experimental procedures were performed.
Neuropsychological data acquisition
EFs
Four EFs (i.e., shifting, working memory, inhibi-tion, and reasoning) were assessed. The corresponding measures for each component were as follows. (1) Shifting: (1a) perseverative errors on the Modified Card Sorting Test [27], (1b) the delta time on the Color Trails Test (i.e., the result of subtracting the completion time of trial 1 from that of trial 2) [28], and (1c) the total number of correct designs in the switching condition on the Design Fluency Test from the Delis–Kaplan Executive Function System (D-KEFS) [29]. (2) Working memory: (2a) the total score of correct answers on the Letter Number Sequencing subtest of the Wechsler Adult Intelligence Scale-Third Edition (WAIS-III) [30] and (2b) the average percentage of correct answers (the score of total correct responses divided by the total possible score) on the spatial short-term memory (SSTM) task [31]. (3) Inhibition: (3a) the completion time for the inhibition condition in the Color–Word Interference Test of the D-KEFS [29], (3b) the delta reaction time (the result of subtracting the reaction time in the automatic condition from that in the inhibition condition) of the Hayling Test [32], and (3c) the stop-signal reaction time in the stop signal task [33]. (4) R easoning: (4a) the total score in the Similarity subtest and (4b) the total score in the Matrix Reasoning subtest of the WAIS-III [30]. Four composite variables were computed to represent the four EF components to amply the potential effect and reduce the type I error caused by multiple comparisons. Each of the four composite variables was generated by converting each participant’s scores on all measures of the corresponding cognitive domain into z-scores based on the mean and standard deviations of the full cohort. Logarithmic transformation or square root transformation was applied to variables that violated normality before conversion into z-scores. Moreover, to facilitate interpretation, reaction time variables were inverted for higher scores to represent better performance. All the z-scores of the corresponding EF component were then averaged.
Other neuropsychological functions and clinical characteristics
The MMSE and four neuropsychological tests, namely for assessing memory function (Logical Memory [LM] subtest of the Wechsler Memory Scale; Wechsler, 1997), attention span (digit span forward length of the WAIS-III), visuospatial ability (Block Design subtest of the WAIS-III), and processing speed (Digit Symbol Substitution subtests of the WAIS-III), were included. Relevant clinical characteristics, including a history of diabetes mellitus, hypertension or hyperlipidemia status, and family history of Alzheimer’s disease (AD), were also obtained. The CDR [34] and Geriatric Depression Scale (GDS)—short form [35] were also included to evaluate functional independence and depression status, respectively.
Statistical analyses
Group differences were analyzed using chi-squ-ared tests for categorical variables (e.g., sex, diabetes mellitus, hypertension, hyperlipidemia, and family history of AD), independent-sample t tests or analysis of variance for continuous variables with a normal distribution, and nonparametric tests (including the Mann–Whitney U test for two independent samples and Kruskal–Wallis H test for four independent samples) for continuous variables with nonnormal distributions. The significance level was set at p < 0.05 for the demographic and clinical variables, p < 0.0083 (Bonferroni correction) for the standardized neuropsychological variables, and p < 0.0125 (Bonferroni correction) for the executive function indexes to examine the overall group-level analyses. For post hoc analysis, pairwise Mann–Whitney U tests or Fisher’s least significant difference tests were performed as appropriate, with the alpha level set at p < 0.05. Hedges’ g, an unbiased estimate for small samples [36], was used to calculate the effect size for significant pairwise comparisons.
Furthermore, due to concerns over our relatively small sample sizes for the analytical analyses, separate moderation analysis was performed using the whole sample. Specifically, we used the PROCESS macro (version 3.5) to examine educational attainment as a potential moderator of the relationship between the APOE ɛ4 genotype and the performance of each of the four executive components. In the analysis, APOE ɛ4 status (dummy coded as 1 =ɛ4+, 0 =ɛ4–) was entered as a predictor. Years of educational attainment (continuous variable) was also entered as a predictor, along with the interaction term between APOE ɛ4 status and educational attainment, to predict the performance of the four EF components. The education variable was mean-centered (i.e., the variable mean was subtracted from every value of the variable) prior to the analysis. The age and sex variables were entered as covariates in all analyses. Results were presented as regression coefficients, reported as significant at the threshold of p < 0.0125 (Bonferroni correction) for each overall model, and were generated from a bootstrap resampling procedure with 5000 draws [37, 38]. Significant interaction effects were further examined through simple slope tests of the relationship between the APOE ɛ4 genotype at one standard deviation below the mean, the mean, and one standard deviation above the mean with the education variable, with the alpha level setting at p < 0.05. All analyses were performed using SPSS (Version 26.0, IBM Corp., Armonk, NY, USA).
RESULTS
Demographic and clinical characteristics
The ɛ4 + and ɛ4–groups did not differ in terms of age, educational level, frequency distribution of sex, history of diabetes mellitus, hypertension, hyperlipidemia, family history of AD, GDS scores, or CDR sum of boxes scores (all p values > 0.05, Table 1). The APOE genotype and CR subgroups did not differ in terms of age, frequency distribution of sex, history of diabetes mellitus, hypertension or hyperlipidemia status, family history of AD, GDS scores, or CDR sum of boxes scores (all p values > 0.05). However, educational attainment did differ significantly between subgroups (Kruskal–-Wallis H test, χ2 (3,N = 68) = 51.05, p < 0.001) with the two HCR groups having higher educational attainment levels than the two LCR groups. No difference in educational attainment was observed within the HCR or LCR groups (Table 2).
Demographic, clinical, and cognitive characteristics of the APOE ɛ4 carrier (ɛ4+) and noncarrier (ɛ4–) groups
AcSS, age-corrected scale score; AD, Alzheimer’s disease; CDR-SB, sum of boxes on the Clinical Dementia Rating scale; LM, Logical Memory subtest of the Wechsler Memory Scale-III; MMSE, Mini-Mental State Examination. aTwo-tailed p values on the Mann–Whitney U test. bTwo-tailed p values on the χ2 test. cTwo-tailed p values on independent-sample t tests.
Demographic and clinical characteristics of the four APOE genotype (ɛ4 + versus ɛ4–) by cognitive reserve (high versus low) groups
AD, Alzheimer’s disease; CDR-SB, sum of boxes score on the Clinical Dementia Rating scale; HCR, high cognitive reserve; LCR, low cognitive reserve. aTwo-tailed p values for analysis of variance. bTwo-tailed p values on the Kruskal–Wallis H test. Two-tailed post hoc comparisons were performed using a Mann–Whitney U test. cTwo-tailed p values on the χ2 test. d1, ɛ4 + HCR group; 2, ɛ4 + LCR group; 3, ɛ4–HCR group; 4, ɛ4–LCR group. *p<0.05 for comparisons between the four subgroups.
APOE effect on cognition
No significant differences were observed between the APOE ɛ4 + and ɛ4–groups in terms of performance on the MMSE, LM (both immediate and delayed recall), Digit Span Forward Length, Block Design, or Digit Symbol Substitution subtests or on the four EF composite indexes (all p values > 0.05, Table 1).
APOE by cognitive reserve effect on cognition
Performance on the MMSE, LM (both immediate and delayed recall), Digit Span Forward Length, Block Design, and Digit Symbol Substitution subtests did not differ between the four APOE genotype and CR subgroups (Table 3). The shifting and working memory composite index of the four subgroups did not differ, whereas the inhibition (F (3,53) = 4.29, p = 0.009, η2 = 0.20) and reasoning (F (3,64) = 8.13, p < 0.001, η2 = 0.28) indexes differed significantly.
Cognitive characteristics of the four APOE genotype (ɛ4 + versus ɛ4–) by cognitive reserve (high versus low) groups
AcSS, age-corrected scale score; HCR, high cognitive reserve; LCR, low cognitive reserve; LM, Logical Memory subtest of the Wechsler Memory Scale-III; MMSE, Mini-Mental State Examination. aTwo-tailed p values on the Kruskal–Wallis H test. bTwo-tailed p values for analysis of variance. Post hoc comparisons were performed using the least significant difference. c1, ɛ4 + HCR group; 2, ɛ4 + LCR group; 3, ɛ4–HCR group; 4, ɛ4–LCR group. *p<0.05 for comparisons between the four subgroups.
Post hoc analyses revealed that the APOE ɛ4 + LCR demonstrated significantly poorer performance on the inhibition composite index compared with the ɛ4 + HCR group (p = 0.001, Hedges’ g = 1.59), ɛ4–HCR group (p = 0.004, Hedges’ g = 0.94), and ɛ4–LCR group (p = 0.019, Hedges’ g = 1.14). Moreover, both HCR groups performed significantly better on the reasoning composite index than the two LCR groups (ɛ4 + HCR versus ɛ4 + LCR: p = 0.001, Hedges’ g = 1.34; ɛ4 + HCR versus ɛ4–LCR: p < 0.001, Hedges’ g = 1.36; ɛ4–HCR versus ɛ4 + LCR: p = 0.003, Hedges’ g = 1.07; ɛ4–HCR versus ɛ4–LCR: p = 0.001, Hedges’ g = 1.10) (Fig. 1).
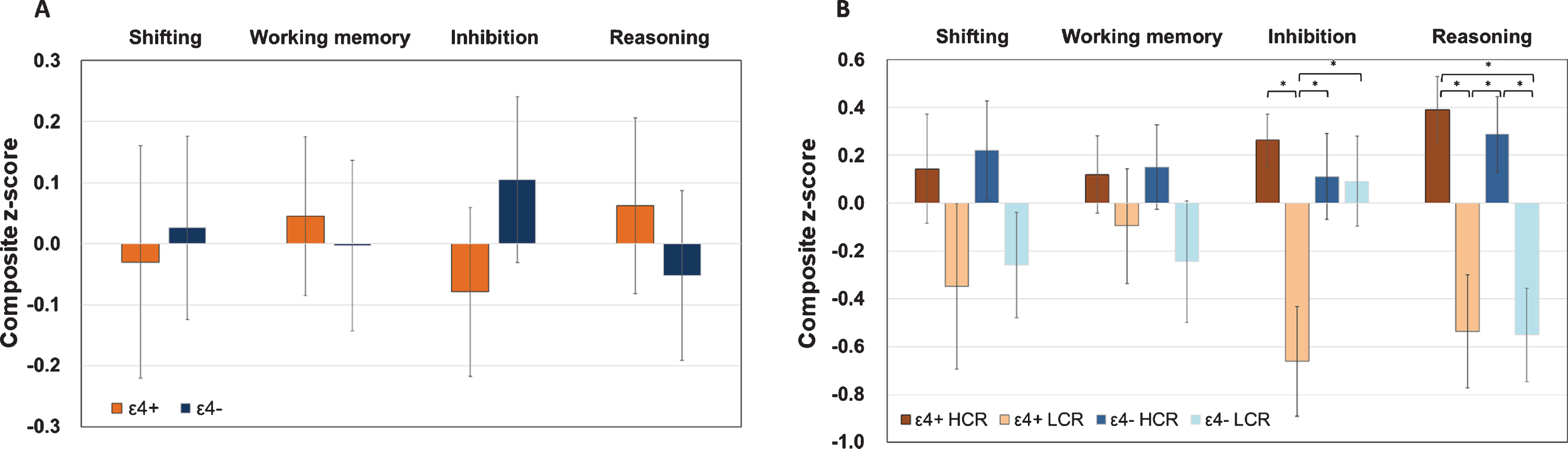
Performance on the four executive function indexes (A) for the apolipoprotein E (APOE) ɛ4 carriers (ɛ4+) and noncarriers (ɛ4–) and (B) for the four APOE genotype by cognitive reserve subgroups: ɛ4 + high cognitive reserve (HCR), ɛ4 + low cognitive reserve (LCR), ɛ4–HCR, ɛ4–LCR. Error bars denote the standard error of the mean. *p<0.05.
Education as a moderator
In the regression analysis for shifting and working memory composite indexes, after we controlled for participants’ age and sex, neither educational attainment (both p values > 0.05) nor APOE genotype (both p values > 0.05) was a significant independent predictor of shifting ability. Furthermore, the APOE by education interaction term was not statistically significant (both p values > 0.05).
In the regression analysis for inhibition ability, the total model was significant (F (5,62) = 3.79, p = 0.01) and explained approximately 27%of the variance in the inhibition composite index. In this model, after we controlled for participants’ age and sex, neither educational attainment nor APOE genotype was a significant independent predictor of inhibition ability (both p values > 0.05). However, the interaction term was statistically significant (b = 0.15, 95%CI [0.01, 0.28], t = 2.17, p = 0.03) in the model, indicating that education was a moderator of the effect of APOE ɛ4 on inhibition ability. In this model, the education×APOE interaction increased R2 by approximately 7%(p = 0.03). A simple slope analysis was subsequently conducted to aid interpretation of this interaction, and the results revealed a relationship between APOE genotype and the inhibition composite index only when educational level was at a low level (i.e., one standard deviation below the mean; b = –0.60, 95%CI [–1.14, –0.05], t = –2.21, p = 0.03), but not when it was at an average level (b = –0.18, 95%CI [–0.53, 0.18], t = –1.00, p = 0.32) or a high level (i.e., one standard deviation above the mean; b = 0.24, 95%CI [–0.27, 0.75], t = 0.95, p = 0.35). Further analysis employing the Johnson–Neyman technique indicated that for older adults with 12 years of education or fewer, the APOE ɛ4 genotype and inhibition performance were significantly related (b = –0.45, t = –2.01, p = 0.05). As educational attainment decreased, the relationship between the APOE ɛ4 genotype and inhibition performance became increasingly negative. The lowest educational attainment was 6 years (b = –1.38, t = –2.33, p = 0.02; Fig. 2A).
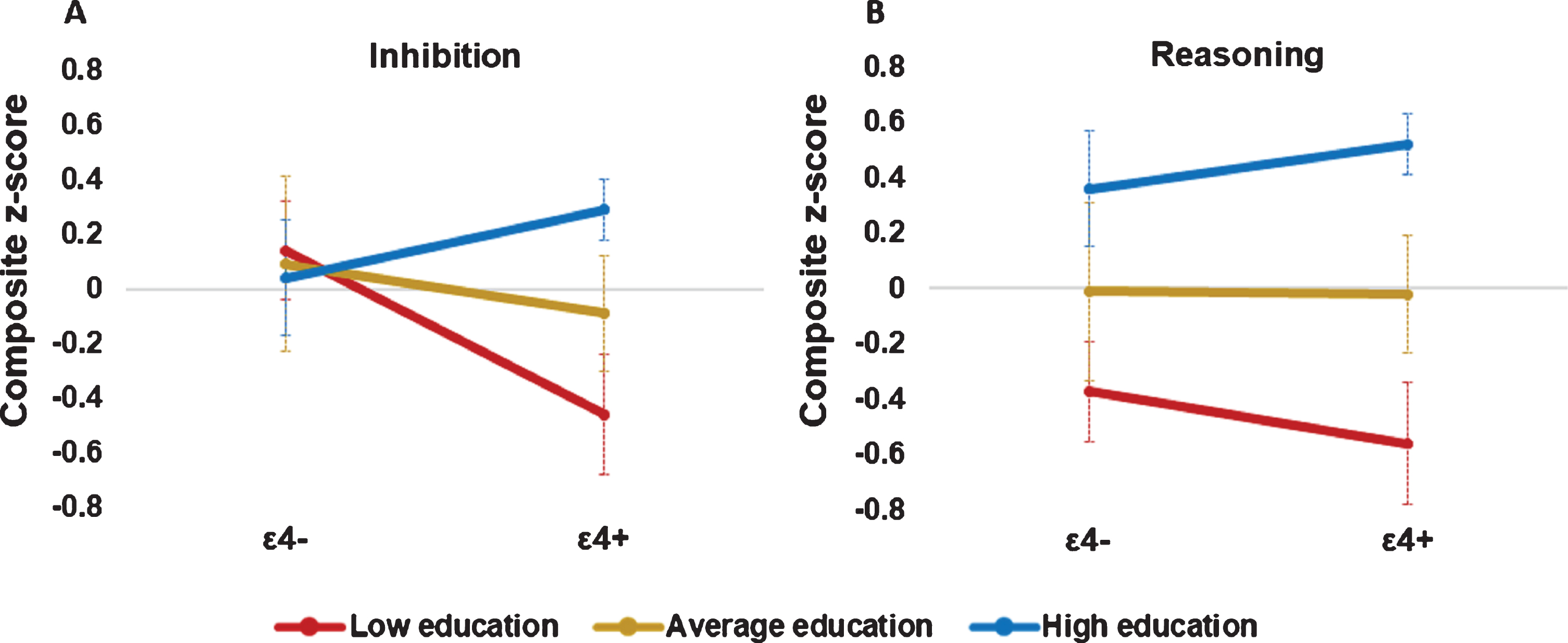
A) Moderation effect of educational attainment on the relationship between the apolipoprotein E genotype (ɛ4 carriers: ɛ4+; noncarriers: ɛ4–) and performance on the inhibition composite index. B) Main effect of educational attainment on the reasoning composite index. Lines represent means of inhibition for high (i.e., one standard deviation above the mean), average (i.e., mean), and low (i.e., one standard deviation below the mean) levels of educational attainment among older adults. Error bars denote the standard error of the mean.
In the regression analysis for reasoning ability, the total model was significant (F (5,62) = 8.09, p < 0.001) and explained 39%of the variance in the reasoning composite index. In this model, after we controlled for participants’ age and sex, educational attainment was a significant independent predictor of reasoning ability (b = 0.13, 95%CI [0.06, 0.20], t = 3.59, p < 0.001; Fig. 2B), while APOE genotype was not (p > 0.05). The APOE×education interaction term was not statistically significant (p > 0.05) in the model.
DISCUSSION
The present study investigated the cognitive phenotype of APOE and the interactions between educational attainment and APOE ɛ4 allele with a focus on EFs. To overcome the methodological drawbacks of previous studies, we measured four EFs (i.e., shifting, working memory, inhibition, and reasoning) using a relatively comprehensive battery, and we carefully matched groups for demographic and clinical variables. The results revealed that although no distinct cognitive phenotypes associated with APOE status were observed in EF components, a gene–education interaction was observed in the inhibition domain; a higher education level increased the domain-specific cognitive function of ɛ4 carriers to the same level as ɛ4 noncarriers. The regression-based moderation analyses with the entire cohort also yielded the same findings, indicating that educational attainment was a significant moderator accounting for the relationship between the APOE ɛ4 allele and inhibition performance, even when the effects of age and sex had been controlled for. For older adults with 12 years of education or fewer, the relationship between the APOE ɛ4 genotype and inhibition performance became increasingly negative. A main effect of education on the reasoning index was also observed.
The scores for EFs and other cognitive measures did not differ between APOE ɛ4 carriers and noncarriers; thus, our hypothesis was not supported. Our findings echoed those from meta-analysis studies, which have indicated that no consistent association exists between the APOE ɛ4 genotype and EFs [1, 5]. We identified a significant interaction effect between APOE ɛ4 status and educational attainment level. Therefore, negative effects associated with the APOE ɛ4 allele on EFs may be negated or inconsistently observed if participants are not grouped according to educational attainment level, which could partially explain the inconsistent findings among relevant studies.
The present study extended the literature on the effect of high educational attainment level in buffering the negative effect of APOE ɛ4 on memory and language to the domain of EF. Notably, we det-ermined that the gene–environment interaction was only manifested in the ability to inhibit prepotent responses and not shifting, working memory, or abstract reasoning functions. Evidence suggests that performance on inhibition tasks continues to improve until young adulthood (early twenties), partly related to the ongoing metacognitive development [39, 40] and increased myelination of frontostriatal connections [41]. By contrast, mental set switching for complex shifting tasks and working memory at a moderately difficult level (e.g., six locations used for the SSTM task used in the present study) typically reach adult levels by middle adolescence [42]. These findings echo the results of a longitudinal twin study [43] that investigated genetic and environmental contributions to stability and changes in EFs from late adolescence to young adulthood (17 to 23 years old). Their results revealed that the contribution of nonshared environmental influences (e.g., changes related to education) to changes in performance on inhibition measures (15%to 34%) over time was greater than that to working memory and shifting abilities (0%to 15%) [43]. These findings accord with our findings and suggest that continuous education throughout young adulthood (e.g., attending college) may benefit the continuous development of inhibition ability to a greater degree, where a gene–education interaction would be most likely occur, than that of shifting or working memory functions. Notably, studies have indicated that inhibition is the most vulnerable component among different EF components in patients with AD [44] or MCI [45] compared with healthy controls. Our findings on the poor performance of ɛ4 carriers with relatively low education for the inhibition aggregate variable but not other aggregate variables indicate that inhibition may be a crucial explanatory factor for the cascades caused by APOE ɛ4 in the trajectories to AD development.
Studies on neural mechanisms and brain function have provided parallel evidence that explains our findings on the unique effect of the gene–environment interaction on inhibition function. Several studies have indicated that inhibition relies heavily on the right inferior frontal and associated networks [46]. Moreover, Robertson [47] proposed a noradrenergic theory to explain the underlying mechanism of cognitive reserve. The theory posited that cognitive reserve develops through repeated noradrenaline activation, and the neurocognitive correlates of noradrenergic activity, including arousal, sustained attention, response to novelty, and awareness, are located in the right hemisphere [48]. The present findings indicate that highly educated older adults could buffer the detrimental effect of the APOE ɛ4 genotype on functions that rely heavily on the right frontal regions. Consistent with this evidence, one study [49] reported differential effects between APOE status and educational attainment level on regional metabolism among less-educated APOE ɛ4 carriers but not more-educated ɛ4 carriers, suggesting reduced metabolism in the right prefrontal areas compared with noncarriers. However, related studies are scarce, and further imaging and behavioral studies are required to confirm the brain–behavior connection.
A main effect of educational attainment on abstract reasoning ability was observed in the present study, which accords with studies that reported that a higher cognitive reserve, calculated as the sum index of the scores on the vocabulary test, educational level, and occupation type, was associated with better abstract reasoning ability [50]. People with lower educational levels were reported to have a greater decline in reasoning ability [51]. Furthermore, reasoning measures have frequently been used to estimate scores on an intellectual battery that is highly correlated with education [52, 53]. Overall, our findings regarding the reasoning index indicate a close relationship between educational effects and abstract reasoning ability in older adults.
We used the number of years of formal education completed in early life as a variable of interest. Studies have indicated that receiving a college-level education in late adulthood improves performance on language-related tests (i.e., vocabulary and comprehension tests and the Boston Naming Test) but not EFs [54, 55]. A high educational level is one of the most frequently studied protective factors for cognitive aging and AD [56], and findings have demonstrated that the positive effect of educational attainment remains even after considering the influence of socioeconomic status or access to resources [23]. However, the effects of the timing of formal education on different cognitive functions, the underlying neural mechanisms of structuring and restructuring of the brain, and network development remain unclear. Education received during different periods of life likely exerts different effects on various cognitive domains and brain networks, depending on the developmental stage of the individual and the content of the education. Further studies are thus warranted.
Notably, despite the existence of a large body of research on EFs, both a universal definition and a standardized form of measurement remain unavailable [15]. The measures selected to represent each of the four EF indexes in the present study were based on those in studies that employed methods such as confirmatory factor analysis [13–15, 57–60]. Our results concerning the gene–education interaction in inhibition function but not in other EF indexes are consistent with findings concerning neural mechanisms and developmental dynamics related to gene–environment interactions in the same EFs, thereby supporting the decision to separate these EF indexes to examine gene–environment interactions. However, caution should be exercised when interpreting our findings because each index does not necessarily represent an orthogonal construct, particularly in the medium to large correlations often observed between the latent variables of different EFs [14, 59]. No EF measure requires only one EF [57, 61], and the factor structures identified in terms of EFs in young adults may not be the same as those identified in older adults, even when identical tests are employed [57]. Moreover, diverse models have also been proposed to account for the complex intrafactor relationship within an EF construct. For example, studies have identified three subcomponents underlying the response inhibition construct: action withholding, interference resolution, and action cancellation, which indicated the involvement of both common and distinct neural substrates [62, 63]. Overall, these findings highlight the complexity of EFs and the importance of considering factors such as participant characteristics (e.g., genetic makeup and environmental contribution) and the specific cognitive processes supporting a task rather than the task itself. Future research is required to obtain a thorough understanding of the complex nature–nurture interactions of EFs [57].
Despite the novel findings of the present study, some limitations should be noted. First, the present study had a relatively small sample size, especially in the two low educational attainment subgroups. However, our results displayed large effect sizes (all Hedges’ g > 0.9). In post hoc power analysis conducted, a statistical power of 0.93, well exceeding the standard requirement of 0.8, was obtained according to the estimated power curve for the final sample size of 68 and a mean Cohen’s f effect size of 0.5. Additionally, both the group-level analyses and regression-based moderation analyses yielded the same results, indicating the robustness of the findings. The comparable results demonstrated that our sample size was sufficient for adequate power. Second, the heterogeneity in EF performance caused by other genetic factors could not be excluded. Genetic factors, such as COMT, BDNF, and TOMM40, have been reported to contribute to individual differences in cognitive aging [64]. The possible gene–gene interaction in cognitive aging should be considered in future research. Third, this study was cross-sectional, which precludes determining the direction of observed associations. Furthermore, despite our relatively stringent methods, the effect of incipient AD cannot be fully ruled out.
In conclusion, the present study was the first to reveal the benefits of advanced education on modifying potentially negative cognitive effects on EFs among older APOE ɛ4 carriers, which offers a favorable outlook for people with a genetic predisposition for AD. This study also highlights the unique role of inhibition function as a factor that may help to elucidate the cognitive cascades related to APOE ɛ4 that increase the risk of developing AD. Aging is associated with increased individual variability in cognitive functions, and our findings highlight the importance of considering factors of both nature and nurture to understand the complex process of cognitive aging.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Professors Yi-Yu Yeh and Chi-Hung Juan for their helpful comments on an early draft of this paper.
This work was supported by the Ministry of Science and Technology, Taiwan (grant numbers 109-2629-H-002-001-MY3, 108-2410-H-002-106-MY2, and 105-2420-H-002-003-MY2 to Y.L.C.).