
Abstract
Background:
Alzheimer’s disease (AD) is a degenerative disorder, accompanied by progressive cognitive decline, for which there is no cure. Recently, the close correlation between AD and type 2 diabetes mellitus (T2DM) has been noted, and a promising anti-AD strategy is the use of anti-T2DM drugs.
Objective:
To investigate if the novel glucagon-like peptide-1 (GLP-1)/glucose-dependent insulinotropic polypeptide (GIP) receptor agonist DA4-JC shows protective effects in the triple APP/PS1/tau mouse model of AD.
Methods:
A battery of behavioral tests were followed by in vivo recording of long-term potentiation (LTP) in the hippocampus, quantified synapses using the Golgi method, and biochemical analysis of biomarkers.
Results:
DA4-JC improved cognitive impairment in a range of tests and relieved pathological features of APP/PS1/tau mice, enhanced LTP in the hippocampus, increased numbers of synapses and dendritic spines, upregulating levels of post-synaptic density protein 95 (PSD95) and synaptophysin (SYP), normalized volume and numbers of mitochondria and improving the phosphatase and tensin homologue induced putative kinase 1 (PINK1) - Parkin mitophagy signaling pathway, while downregulating amyloid, p-tau, and autophagy marker P62 levels.
Conclusion:
DA4-JC is a promising drug for the treatment of AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a progressively neurodegenerative disease, characterized by declining cognitive function [1]. AD is the major cause of dementia-associated morbidity and mortality in elderly population [2]. The main pathological hallmarks of AD are amyloid plaques consisting of extracellular amyloid-β (Aβ) and neurofibrillary tangles containing hyperphosphorylated tau protein in neurons [3]. Although AD has been extensively studied for a long time, there is still no effective therapy for treating AD.
According to epidemiologic evidence, 70–80%of AD patients had type 2 diabetes mellitus (T2DM), which suggested that T2DM was an important risk factor for AD [4]. Growing amounts of data demonstrated that T2DM and AD shared many clinical and pathological features [5]. In view of the close association between T2DM and AD, drugs originally developed for T2DM treatment have been tested as the potential treatment for AD [6]. Lately, a clinical study demonstrated that insulin showed beneficial effects in AD patients, but chronic administration of insulin could increase insulin desensitization [7, 8]. To avoid insulin desensitization, glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP), which are incretin hormones and have similar functions as insulin, have been developed as potential treatments for AD that increase insulin sensitivity among other beneficial effects [6, 10]. Our studies with GLP-1 analogs lixisenatide, liraglutide, and CJC-1131 showed their neuroprotective effects against Aβ-induced impairments in spatial reference memory and synaptic plasticity of rats [11–13] and in APP/PS1 mouse model of AD [14–16]. Importantly, a phase II clinical trial testing liraglutide in AD patients showed protective effects by improving memory and cognition and reducing brain shrinkage as shown in MRI brain scans [17]. Our studies with GIP analogues also showed good neuroprotective effect, protecting against cognitive deficits and pathology in APP/PS1 mice by inhibiting neuroinflammation and upregulating the cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA)/cAMP-response element binding protein (CREB) growth-factor signaling pathway [18–20]. Most recently, GLP-1/GIP dual receptor agonists, simultaneously activating GLP-1 and GIP receptors, which are expressed in many brain regions, including hippocampus, have been reported to have higher efficacy as compared to single receptor agonists [21].
A novel GLP-1/GIP dual receptor agonist (DA4-JC) has been developed that contains a poly-lys cell-penetrating sequence (CPS) in order to enhance crossing of blood-brain barrier (BBB), which is important in treating diseases of central nervous system [6]. DA4-JC has showed superior neuroprotective effect to GLP-1 analogs in a mouse model of Parkinson’s disease (PD) [22] and APP/PS1 mouse model of AD [23]. However, DA4-JC has never been tested in APP/PS1/tau transgenic model of AD. In the study, we chose APP/PS1/tau (3xTg-AD) mice, expressing mutated APPSwe, PS1M146V and tauP301L. 3xTg-AD mice are characterized by not only amyloid plaques, but also neurofibrillary tangles in the brain [24]. Therefore, this study observed neuroprotective function of DA4-JC in nine-month-old 3xTgAD mice and explored possible mechanisms of drug action.
MATERIALS AND METHODS
Animals
APP/PS1/tau (3xTg-AD) male and female (28 males and 28 females) mice were purchased from Jackson Laboratory (USA). Wild-type (WT) male and female (28 males and 28 females) mice with the same genetic background (C57BL/6J) were used as control mice. Mice were housed with access to standard rodent food and water ad libitum in an independent air supply system that was maintained on a 12/12 h light-dark cycle at 23±2°C and 55±5%humidity. All animal experimental procedures were approved by the Institutional Animal Care Committee of Shanxi Medical University in Taiyuan of China.
Drug treatments
DA4-JC was synthesized by China Peptides Co, Ltd. (Shanghai, China) to 95%purity. The identity and purity of DA4-JC were confirmed by reversed-phase high performance piquid chromatography (HPLC) and characterized using matrix assisted laser resorption/ionization time of flight (MALDI–TOF) mass spectrometry. DA4-JC was stored in dry form and dissolved in sterile phosphate-buffered saline (PBS) before experiments.
Sequence of DA4-JC [25]:
YXEGTFTSDYSIYLDKQAAXEFVNWLLAGGPSSGAPPPSKKKKKK-NH2
X = aminoisobutyric acid
Eight-month-old mice were divided into four groups (half males and half females in each group): WT + PBS, WT + DA4-JC, 3xTg-AD + PBS, and 3xTg-AD + DA4-JC. Mice were injected daily with DA4-JC (10 nmol/kg, body weight) by intraperitoneal (i.p.) injection or PBS (equal volume) for consecutive 46 days. In behavioral experiments, 20 3xTg-AD mice and 20 WT mice were used, which were also used in LTP recording after behavioral experiments and follow-up Western blot test. At the same time, 12 3xTg-AD mice and 12 WT mice were used in Immunohistochemistry test; 12 3xTg-AD mice and 12 WT mice were used in Transmission electron microscope test; 12 3xTg-AD mice and 12 WT mice were used in Golgi staining test. Experimental procedure was shown in Fig. 1.

Schematic diagram of the experimental procedure. After 3 days of adaptation to the laboratory environment, the first batch of mice were injected daily with DA4-JC (i.p.) or PBS (i.p.) for 30 days before behavioral tests. The injections were kept during all behavioral tests in the first batch of mice, including open field test (OFT), new object recognition test (ORT), Y maze test (YMT), Morris water maze test (classical and reversal MWM), and conditional fear memory test. Following behavioral tests, in vivo hippocampal long-term potentiation (LTP) recording and Western blot were performed. After 3 days of adaptation to the laboratory environment, the second batch of mice were injected daily with DA4-JC (i.p.) or PBS (i.p.) for 30 days and the injections were continued for 16 days without behavioral tests. Golgi staining, immunohistochemistry (IHC), and transmission electron microscope (TEM) were simultaneously performed.
Body weight of mice
Mice were weighed to determine the injection dose. Body weight of mice was measured regularly on day1, day 7, day 14, day 21, day 28, day 35, day 42, and day 49.
Open field test (OFT)
OFT was used to assess locomotor activity of mice. The open field was a square box (40×40×40 cm) made of plastic with black walls and a white floor without lid. The bottom was divided into 16 equal squares, 4 squares in the center were defined as the central area, and rest as the peripheral area. Each mouse was placed in the center of the box and allowed to explore the open field for 5 min under moderately light condition. The spontaneous motor activity of mice was continuously recorded by a video tracking system (Ethovision 3.0 software, Noldus Information Technology, Wageningen, the Netherlands). The total distance and the percentage of time in center were automatically computed. After each trial, fecal boli in the box was removed and the floor was wiped clean with 75%ethanol.
New object recognition test (ORT)
ORT was used to evaluate the ability of mice to recognize new object. The test included familiarization session and test session. In familiarization session, two identical objects were placed in the open field with an interval of 20 cm, and the distance between each object and side wall was 10 cm. Each mouse was placed in the middle line attached to the wall on the side of non-object area. After 10 min of free exploration, mice were taken out, and test session was carried out after 6 h of rest. During test session, one of objects was taken out and replaced by a new object of the same material, but different color and size, and then mice were placed in the same position for free exploration for 10 min. The infrared camera at the central top of the open field was used to collect track of mice. The software analyzed time of mice exploring the old object and the new object and calculated new object recognition index (NOI) of mice = time to explore new object/(time to explore new object + time to explore old object)×100%. After each trial, chamber and objects were sprayed with 75%ethanol and wiped clean with toilet paper to avoid influence of odor on the next trial.
Y maze test (YMT)
The spontaneous alternation of mice was examined in Y maze to examine the spatial working memory. Y maze had three radial arms with equal angles (120°) between arms. Each arm was 30 cm long, 15 cm high, and 5 cm wide. Each mouse was placed at the junction of three arms and allowed free movement for 8 min. Entries of each mouse into each arm were recorded, and every entry that differed from the last two entries was considered a successful alternation. The alternation percentage was calculated according to the formula: [(number of alternations)/(total number of arm entries-2)]×100(%) [26].
Morris water maze (MWM) task
The classical MWM task was used to examine the spatial reference memory of mice. A circular pool (diameter 120 cm, height 50 cm) was filled with 30 cm deep of tap water (maintained at 23±2°C), and titanium dioxide was added to ensure that water was equally white throughout the pool. A white circular platform was hidden 1 cm below water level. In the hidden platform test, mice were allowed to swim in water to search for the underwater platform, which was performed four times per day for 5 consecutive days. In each trial, a mouse was placed in water facing the pool wall in one of four equal quadrants designated by computer software. The order of a mouse entry into individual quadrants was randomized by a number table. Subsequently, 24 h after completing the hidden platform test, each mouse underwent a 60 s probe trial to evaluate retention of the learned task. During the probe trial, the platform was removed, and searching behavior in the target quadrant (where the platform was located during the hidden platform test) was measured. The reversal MWM task was performed on 7th day. In the reversal MWM task, the hidden platform was moved to the opposite quadrant and mice were given four trials (60 s/trial) each day for 4 days to learn the new platform position. On 11th day, the platform was removed and a probe trial lasting for 60 s was conducted. To avoid influence of visual acuity and motor ability on searching behavior of mice, a visible platform test was also performed on 11th day, in which the platform was elevated to 1cm above water surface. Escape latency, time in target quadrant, number of platform crossing and swimming speed were recorded and analyzed with Ethovision 3.0 software (Noldus Information Technology, Wageningen, the Netherlands).
Conditional fear memory test
Conditional fear memory test was used to assess the ability of mice to remember fear. The conditional fear box was a rectangular box that could release acoustic stimulation and plantar electrical stimulation, according to system setting, and a pressure sensor was installed at the bottom of the box. When mice are free to move or receive sound and electrical stimulation, pressure curve with time could be displayed on the computer with change of plantar pressure. In this way, different activity states of mice could be measured. The whole test was divided into training session and test session. Training session: Firstly, mice were allowed to freely move in the box for 90 s, followed by sound stimulation for 30 s (frequency: 2900 Hz, intensity: 90 dB), and direct current stimulation (current intensity: 0.3 mA) was given in the last 2 s of the sound stimulation. The above acoustic stimulation and direct current stimulation were repeated for 5 times, with an interval of 90 s for each time. The software recorded activity states of mice during the whole process and counted freezing ratio of mice before and after training. Test session was conducted 24 h after the end of training session. Before the experiment stared, colored paper could be affixed to the wall of the box to change environment of the box. After the experiment began, mice were put into the box for 150 s, and baseline activity of mice in natural state was recorded by software. Subsequently, 150 s of conditioning stimulus (CS) was administered, which was acoustic stimulation without electrical stimulation (the frequency and intensity of sound wave were the same as that of training session), and activity of mice during CS was recorded by software. The freezing ratio of mice before and after CS was analyzed and compared.
In vivo long-term potentiation (LTP) recording
Mice were anesthetized with 5%chloral hydrate (0.007 ml/g, i.p.) and positioned on a stereotaxic apparatus (68016, RWD Life Science, Shenzhen, China). A small hole with 2.0 mm diameter was drilled in the skull (2.0 mm posterior from bregma and 1.5 mm from midline) for inserting a bound stimulating/recording electrode. Tips of the stimulating and recording electrode were located at the Schaffer collateral/commissural pathway and the stratum radiatum of hippocampal CA1 region, respectively. The accurate orientation of the electrode depended on the electrophysiological criteria in which a pre-synaptic stimulation always induced a negative field excitatory postsynaptic potential (fEPSP). Stimulation intensity was 30–50%of the maximum fEPSP response, and the stimulation frequency was 0.033 Hz for recording of fEPSP. Basal fEPSP was firstly recorded for 30 min. Paired-pulse facilitation (PPF) was induced by two stimuli with an interval of 50 ms to analyze pre-synaptic mechanism and interneuron activity. High-frequency stimulation (HFS) was applied to induce LTP of fEPSP. HFS protocol consisted of 3 trains of 20 stimuli at 200 Hz with an inter-train interval of 30 s. After HFS, fEPSP were recorded again for at least 1 h. Slope of fEPSP was normalized to basal fEPSP and averaged.
Immunohistochemistry (IHC)
Mice were sacrificed by cardiac perfusion-fixation with PBS and 4%paraformaldehyde (PFA). The brain tissue was carefully removed and divided into left hemisphere and right hemisphere. For the frozen section experiment, the left hemisphere was fixed in 4%PFA (pH = 7.4) for 24 h and incubated in 30%sucrose at room temperature for another 24 h. Immediately afterward, tissues were embedded in OCT frozen embedding medium (Heidelberger Strasse 17–19, Nussloch, Germany). Coronal sections were cut at 25μm on a freezing microtome (Leica, CM1850) and stored at 4°C in PBS. Then, sections were incubated with the rapid antigen repair solution (Beyotime, P0090) for 5 min at room temperature, incubated with 5%hydrogen peroxide at room temperature for 15 min and blocked with 5%goat serum (Solarbio, Beijing, China) for 30 min. Sections were incubated with primary antibody (anti-Aβ1–16, 6E10, dilution 1:500, 803105) overnight at 4°C, followed by incubation with secondary antibody (peroxidase-conjugated Affinipure goat anti-rabbit IgG (H + L), 1:200, ZSGB-BIO, Beijing, China) at 37°C for 2 h.
For the paraffin section experiment, the right hemisphere was processed into paraffin-embedded blocks. Sections were cut at 2μm on a Leica microtome (RM2255) and attached to glass slides. The paraffin was removed with xylene. Next, tissues were rehydrated in 100%, 90%, 80%, 70%, and 50%ethanol and washed twice with distilled water. Then, sections were incubated with 3%hydrogen peroxide at room temperature for 15 min and heated in a microwave oven for 3 min with citrate acid buffer (pH = 6.0). Sections were washed three times (15 min each) with PBS (0.5%TritonX-100) and blocked with 5%goat serum for 30 min. Sections were incubated with monoclonal anti-tau (phospho-T231) antibody (1:200, ab151559, Abcam, Cambridge, UK) at 37°C for 1 h, and then with secondary antibody (peroxidase-conjugated Affinipure goat anti-rabbit IgG (H + L), 1:200, ZSGB-BIO) at 37°C for 20 min. The diaminobenzidine method was used for positive area staining. We used Image Pro Plus 6.0 (Fryer, Huntley, IL) to quantify the immunopositive area of amyloid or cells. The images were thresholded to identify “stained” versus “unstained” regions. The threshold was manually set according to obvious immunopositive plaques or cells, and the threshold value did not change during the analysis, then the software will automatically analyzed the positive areas.
Western blot
Hippocampi of mice were dissected and protein was extracted (Tissue Protein Extraction Reagent; Boster, Beijing, China) and supplemented with complete protease inhibitor (Boster). Protein concentration was measured using a bicinchoninic acid protein assay kit after removing debris by low-speed centrifugation. 30μg protein sample was separated on 12%sodium dodecyl sulfate (SDS)–polyacrylamide gel. After electrophoresis, protein was transferred onto a polyvinylidene fluoride (PVDF) membrane and nonspecific binding was blocked with 5%bovine serum albumin (BSA) in Tris-buffered saline containing 0.05%Tween-20 (TBST). The membrane was incubated with the primary antibody overnight at 4°C, followed by the secondary antibody at room temperature for 2 h. The following primary antibodies (all from Abcam) were used: anti-PSD95 (ab76115), anti-SYP (ab52636), anti-PINK1 (ab23707), anti-Parkin (ab77924), anti-P62 (ab109012), GAPDH (ab226408), and β-actin (ab8226) were used as loading controls. The secondary antibodies were anti-rabbit IgG-HRP conjugate (BA1054, BOSTER, China) and anti-mouse IgG-HRP conjugate (BA1050, BOSTER, China). The membrane was rinsed with TBST and the immunocomplex was visualized using an enhanced chemiluminescence detection kit (Beyotime, Shanghai, China). The signal on the membrane was scanned with a FluorChem scanner (ProteinSimple) and quantified with Alpha View SA software (Azure C300 Biosystems, Inc, USA). Alpha View SA software was used to analyze the gray value of target protein and internal reference, and the relative gray value of target protein was calculated. The higher the gray value, the higher the protein expression level.
Transmission electron microscope (TEM)
Mice were anesthetized with isoflurane and chest cavity was opened. Saline was perfused into the heart, and then mice were infused with 4%PFA. Hippocampus was separated and cut into 1 mm3 tissues, which were fixed in glutaric acid at room temperature for 2 h and then turned to 4°C overnight. Tissues were washed with 0.1 M PBS for 3 times and fixed with 1%osmium acid for 2–3 h at room temperature. Next, tissues were washed with with 0.1 M PBS for 3 times, and placed in 50%, 70%, and 90%alcohol respectively for gradient alcohol dehydration, followed by gradient acetone dehydration in alcohol-acetone mixture (ratio of 90%acetone and 90%alcohol was 1:1), 90%and 100%acetone. Tissues were penetrated with embedding agent/acetone (1:2) at room temperature for 4h, embedding agent/acetone (2:1) overnight at room temperature and embedding agent at 37°C for 3 h. Tissues were embedded with embedding agent in the oven at 37°C overnight, at 45°C for 12 h and at 60°C for 48 h. Tissues were cut into 60–80 nm ultra-thin sections. Sections were stained with uranium dioxide acetate for 30 min, and then washed with distilled water for 3 times, followed by staining with lead citrate for 10 min, and then washed with distilled water for 3 times and dried overnight at room temperature. Finally, TEM was used for observation of synapse and mitochondria. Image J software was used to capture the area of each synapse, and the average area of each group of synapses was calculated. The number of synapses per unit area was manually marked and counted. The volume of each mitochondria was counted with Image J software, and the average volume of each group of mitochondria was calculated. The number of mitochondria per unit area was artificially manually marked and counted.
Golgi staining
Mice were injected intraperitoneally with pentobarbital sodium (1%, 50 mg/kg) and killed. Brain tissue was quickly removed and washed with distilled water. Tissue was put into immersion solution (Solution A containing potassium dichromate and mercuric chloride: Solution B containing potassium chromate = 1:1) at room temperature in dark for 14 days, and change immersion solution the next day. After two weeks, tissue was replaced in Solution C (as a trade secret of manufacturer) for 5 days, and solution was changed again the next day. Tissue was removed into dry ice to precool and cut surplus parts. Tissue was placed on the sample plat, which was placed on dry ice and quickly frozen for 10 min. Then the sample plate was immediately installed on a frozen section machine (CM1520, Leica, Germany), which was suitable for pathological and biological tissue sections. The advantages of frozen section machine were that the sample positioning and injection systems could ensure continuous thin slices (the thickness is 100μm); the powerful refrigeration system provided the strong guarantee for rapid freezing sections. After 5 min, sections of 100μm were made. Sections were attached to gelatin slices and dried in the dark. Slices were washed with distilled water twice, dyed with working liquid for 10min, and washed with distilled water twice. Dyed slices were dehydrated in 50%, 75%and 95%ethanol for 4 min at each gradient, then were dehydrated in anhydrous ethanol for 4 times (4 min each). Slices were in xylene for 3 times (4 min each), and then sealed with resin sealing agent. Finally, dendritic spines were observed under microscope. Open the Image J software, dendritic spines were manually marked and the total number of dendritic spines was calculated by the software.
Statistical analysis
All statistical analyses were performed with SPSS 13.0 and Sigma Plot 12.3 software. Escape latency in MWM task and fEPSP slope in LTP recording were analyzed using repeated-measure analysis two-way analysis of variance (ANOVA). Statistical comparison between groups of all other results were completed using two-way ANOVA. Difference between groups was considered statistically significant at p-value less than 0.05. All data were expressed as the mean±standard deviation (SD).
RESULTS
DA4-JC did not affect body weight of mice
No significant difference was found in body weight (Fig. 2, p > 0.05) of four groups. Thus, DA4-JC did not affect body weight in normal weight, non-diabetic mice.
Body weight of mice. Mice were weighed every 7 days over a period of 49 days. Body weight did not differ significantly among four groups (p > 0.05, n = 10 for each group).
DA4-JC did not change locomotor activity of 3xTg-AD mice and promoted ability of 3xTg-AD to recognize new object
As shown in Fig. 3A and 3B, there was no difference in total distance (p > 0.05), and the percentage of time spent in center (p > 0.05) among four groups; suggesting that treatment with DA4-JC did not influence spontaneous motor activity of 3xTg-AD mice. In Fig. 3D, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on NOI of mice (genotype: F (1,36) = 60.3, p < 0.001; treatment: F (1,36) = 8.884, p = 0.0051; genotype×treatment: F (1,36) = 4.821, p = 0.0346). Compared to WT + PBS group (65.17%±3.54%), NOI was obviously lower in 3xTg-AD + PBS group (52.66%±2.17%, p < 0.001). DA4-JC prevented the decreased NOI of 3xTg-AD mice (59.16%±5.47%, p = 0.0043).

DA4-JC did not influence locomotor activity of 3xTg-AD mice and promoted ability of 3xTg-AD mice to recognize new object. A, B) There was no difference in total distance and the percentage of time in center among four groups (p > 0.05, n = 10 for each group). C) Representative running traces of four groups in the open field. The green circle and the red circle represent initial position and stop position of mice, respectively. The red square frame is the defined central area. D) NOI of four groups. PBS + 3xTg-AD mice showed lower NOI than PBS + WT mice and a measurable raise was seen in DA4-JC + 3xTg-AD mice. *p < 0.05, ***p < 0.001, n = 10 for each group.
DA4-JC improved spatial working memory of 3xTg-AD mice
In Fig. 4C, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on spontaneous alternation of mice (genotype: F (1,36) = 24.85, p < 0.001; treatment: F (1,36) = 10.02, p = 0.0031; genotype×treatment: F (1,36) = 3.306, p = 0.0774). Compared with WT + PBS group (64.81%±3.18%), spontaneous alternation was significantly lower in 3xTg-AD + PBS group (54.65%±3.29%, p < 0.001). DA4-JC improved spontaneous alternation of 3xTg-AD mice (62.10%±4.21%, p = 0.007). However, total arm entries of mice did not show any significant difference among four groups (Fig. 4B, p > 0.05), suggesting that difference in spontaneous alternation among these groups were due to impaired spatial working memory rather than impaired locomotor activity.

DA4-JC improved spatial working memory of 3xTg-AD mice. A) Y maze diagram. B) There was no difference in total arm entries in four groups (p > 0.05). C) Histograms showed spontaneous alternation in four groups. PBS + 3xTg-AD mice showed lower spontaneous alternation than PBS + WT mice and a significant increase was seen in DA4-JC + 3xTg-AD mice. **p < 0.01, ***sp < 0.001, n = 10 for each group.
DA4-JC reversed impairment of spatial reference memory and cognitive flexibility of 3xTg-AD mice
In classical MWM task, as expected, escape latency of mice to find the hidden platform decreased over 5 consecutive days. As shown in Fig. 5A, no significant difference among escape latency of four groups were detected in the first 2 days; however, escape latency on training days 3, 4, and 5 was longer in 3xTg-AD + PBS group than WT + PBS group (p = 0.0038; p < 0.001; p < 0.001). DA4-JC reversed the increase of escape latency on training days 4 and 5 (p = 0.005; p = 0.0025). These results indicated that spatial reference learning disability of 3xTg-AD mice could be reversed by DA4-JC. Probe trial was performed on day 6 to assess spatial reference memory of mice. In Fig. 5B, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on time in target quadrant of mice (genotype: F (1,36) = 41.67, p < 0.001; treatment: F (1,36) = 2.619, p = 0.1143; genotype×treatment: F (1,36) = 12.24, p = 0.0013). Time in target quadrant was clearly lower in 3xTg-AD + PBS group (31.15%±2.06%, p < 0.001), compared to WT + PBS group (42.71%±3.63%). DA4-JC increased time in target quadrant of 3xTg-AD mice (37.09%±4.07%, p = 0.0048). In Fig. 5C, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on number of platform crossing of mice (genotype: F (1,36) = 34.02, p < 0.001; treatment: F (1,36) = 7.483, p = 0.0096; genotype×treatment: F (1,36) = 4.699, p = 0.0369). Number of platform crossing was obviously less for 3xTg-AD + PBS group (3.20±0.75, p < 0.001) than WT + PBS group (4.75±0.55). DA4-JC raised number of platform crossing of 3xTg-AD mice (4.15±0.58, p = 0.0072). These results indicated that DA4-JC ameliorated spatial reference memory deficit of 3xTg-AD mice.

DA4-JC reversed impairments of spatial reference memory and cognitive flexibility of 3xTg-AD mice. Classical MWM (A–D) and reversal MWM (E–I) tests were conducted at days 1–6 and 7–11, respectively. A, E) Escape latency of four groups in hidden platform test. Escape latency was significantly longer in PBS + 3xTg-AD mice on days 3, 4, 5, 9, and 10, which was reversed by DA4-JC on days 4, 5, 9, and 10. **p < 0.01, ***p < 0.001 versus PBS + WT mice; #p < 0.05, # #p < 0.01, # # #p < 0.001 versus PBS + 3xTg-AD mice, n = 10 for each group. B, F) Time in target quadrant, and (C and G) number of platform crossings in the probe trial. 3xTg-AD mice treated with DA4-JC spent more time in target quadrant and made more platform crossings than 3xTg-AD controls. **p < 0.01, ***p < 0.001, n = 10 for each group. D, I) Sample traces of four groups in probe trial on days 6 and 11, respectively. H) Swimming speed between four groups was not changed (p > 0.05).
Furthermore, cognitive flexibility of mice was assessed by following reversal MWM task. Similarly, as shown in Fig. 5E, no significant difference among escape latency of four groups were detected in the first 2 days; however, escape latency on training days 9 and 10 was longer in 3xTg-AD + PBS group than that of WT + PBS group (p < 0.001; p < 0.001). The increase in escape latency was reversed by DA4-JC on training days 9 and 10 (p = 0.0179; p < 0.001). Probe trial was performed on day 11. In Fig. 5F, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on time in target quadrant of mice (genotype: F (1,36) = 52.08, p < 0.001; treatment: F (1,36) = 5.659, p = 0.0228; genotype×treatment: F (1,36) = 9.261, p = 0.0044). Time in target quadrant of 3xTg-AD + PBS group (32.75%±4.62%) was significantly lower than WT + PBS group (44.66%±3.26%, P < 0.001). After DA4-JC treatment, time in target quadrant of 3xTg-AD mice was increased (39.04%±3.16%, p = 0.0026). In Fig. 5G, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on number of platform crossing of mice (genotype: F (1,36) = 52.06, p < 0.001; treatment: F (1,36) = 7.323, p = 0.0103; genotype×treatment: F (1,36) = 5.673, p = 0.0226). Number of platform crossing was evidently less in 3xTg-AD + PBS group (3.42±0.42, p < 0.001), compared with WT + PBS group (4.94±0.61). DA4-JC reversed the decreased number of platform crossing of 3xTg-AD mice (4.23±0.55, p = 0.0051). These data indicated that PBS-treated 3xTg-AD mice were slower to extinguish memory of original target and new target; DA4-JC improved cognitive flexibility of 3xTg-AD mice.
Visible platform test was performed on day 11 for mice to exclude visual or motor dysfunction. As shown in Fig. 5H, swimming speed did not differ significantly among all groups (p > 0.05), thereby suggesting that altered escape latency, time in target quadrant and number of platform crossing were due to difference in spatial reference learning and memory, and cognitive flexibility impairments rather than visual or motor ability of mice.
DA4-JC enhanced ability of 3xTg-AD mice to remember fear
In training session, freezing ratio of WT + PBS mice and 3xTg-AD + PBS mice increased with increase of reinforcement times (p > 0.05, Fig. 6B, C). In test session (Fig. 6E, F), two-way ANOVA showed that genotype and treatment had significant main effect and interaction on freezing ratio of mice (genotype: F (1,36) = 43.13, p < 0.001; treatment: F (1,36) = 1.499, p = 0.2287; genotype×treatment: F (1,36) = 5.457, p = 0.0252). Compared to WT + PBS group (82.55%±3.43%), freezing ratio was clearly lower in 3xTg-AD + PBS group (69.88%±5.14%, p < 0.001). DA4-JC increased freezing ratio of 3xTg-AD mice (74.95.71%±4.92%, p = 0.0097).

DA4-JC enhanced ability of 3xTg-AD mice to remember fear. A) Diagram of fear memory test. B) Plots showed change in freezing ratio during training session between four groups. C) Histograms showed freezing ratio in four groups before 1st reinforcement and after 5th reinforcement in training session. D) Representative original traces of freezing ratio of four groups in test session. E) Plots showed change of freezing ratio of four groups in pre-CS (white area) and during CS (gray area). F) Histograms showed a significant decrease in freezing ratio of PBS + 3xTg-AD mice during CS, and a significant raise was seen in DA4-JC + 3xTg-AD mice. **p < 0.01, ***p < 0.001, n = 10 for each group.
DA4-JC improved hippocampal synaptic plasticity of 3xTg-AD mice
Considering close association of spatial reference memory with synaptic plasticity in hippocampus, we further examined hippocampal LTP in CA1 region by recording in vivo fEPSP. After stably recording basal fEPSP for 30 min, paired pulses and HFS were applied to Schaffer collateral respectively to induce PPF and LTP of fEPSP. We did not find any difference in percentage of fEPSP2/fEPSP1 among four groups (Fig. 7B, p > 0.05), indicating that presynaptic neurotransmitter release was not changed. In Fig. 7C and 7D, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on fEPSP slope of mice at 30 min post-HFS (genotype: F (1,20) = 117.3, p < 0.001; treatment: F (1,20) = 0.6897, p = 0.4161; genotype×treatment: F (1,20) = 20.02, p < 0.001) and 60 min post-HFS (genotype: F (1,20) = 233.1, p < 0.001; treatment: F (1,20) = 20.27, p < 0.001; genotype×treatment: F (1,20) = 82.64, p < 0.001). fEPSP slope did not show significant difference immediately after HFS between four groups, but a significant reduction of fEPSP slope was found at 30 min and 60 min post-HFS in 3xTg-AD + PBS group (140.70%±6.28%, p < 0.001; 124.48%±2.36%, p < 0.001), compared to that of WT + PBS group (176.78%±4.44%; 174.60%±3.86%). DA4-JC rescued the reduced fEPSP slope of 3xTg-AD mice at 30 min and 60 min post-HFS (149.29%±5.12%, p = 0.0078; 152.45%±8.43%, p < 0.001). DA4-JC could improve hippocampal synaptic plasticity of 3xTg-AD mice.
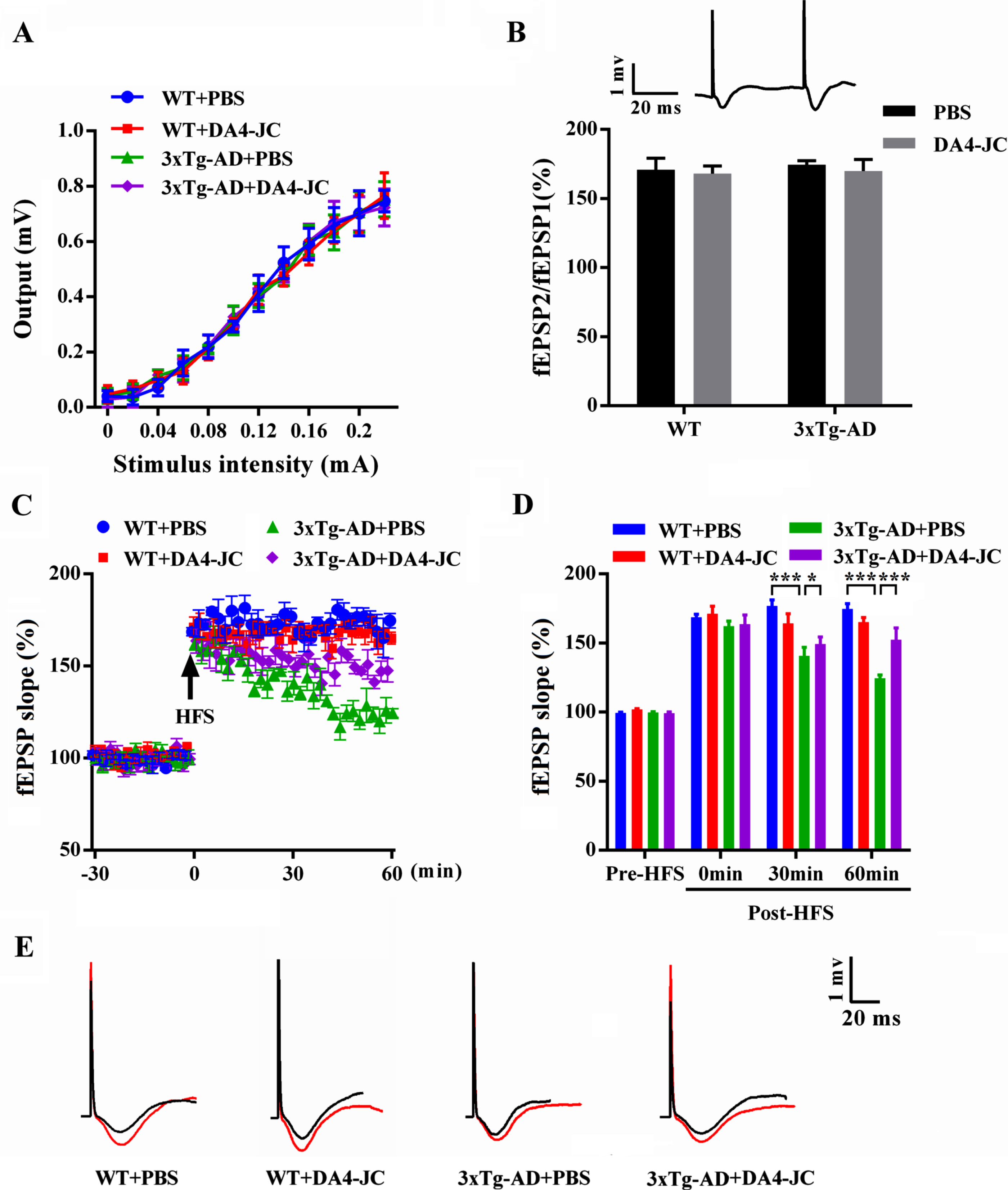
DA4-JC improved hippocampal synaptic plasticity in 3xTg-AD mice. A) There was no difference in I/O curve of four groups (p > 0.05). B) Histograms showed there was no difference in PPF (fEPSP2/fEPSP1) of four groups (p > 0.05). Inset: a sample trace of paired fEPSP, with an increase in amplitude of the second fEPSP. C) Plots represented time course of fEPSP before and after HFS in four groups. D) Histograms showed fEPSP slope in pre-HFS and 0 min, 30 min, and 60 min post-HFS. Compared to WT + PBS group, significant reduction of fEPSP slope was found at 30 min and 60 min post-HFS in 3xTg-AD + PBS group, which was rescued by DA4-JC. *p < 0.05, ***p < 0.001, n = 6 for each group. E) Sample traces of fEPSP before (black line) and after (red line) HFS in four groups.
DA4-JC increased levels of PSD95 and SYP in 3xTg-AD mice
In Fig. 8B, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on PSD95 level of mice (genotype: F (1,20) = 191.6, p < 0.001; treatment: F (1,20) = 7.169, p = 0.0145; genotype×treatment: F (1,20) = 1.648, p = 0.2139). Compared with WT + PBS group (1.00±0.04), level of PSD95 in 3xTg-AD + PBS group was lower (0.55±0.08, p < 0.001). DA4-JC prevented the decreased level of PSD95 in 3xTg-AD mice (0.67±0.07, p < 0.001). In Fig. 8C, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on SYP level of mice (genotype: F (1,20) = 345.9, p < 0.001; treatment: F (1,20) = 26.75, p < 0.001; genotype×treatment: F (1,20) = 11.16, p = 0.0033). Level of SYP was lower in 3xTg-AD + PBS group (0.50±0.05, p < 0.001) than WT + PBS group (1.00±0.07). After DA4-JC treatment, level of SYP in 3xTg-AD mice was increased (0.69±0.03, p < 0.001).
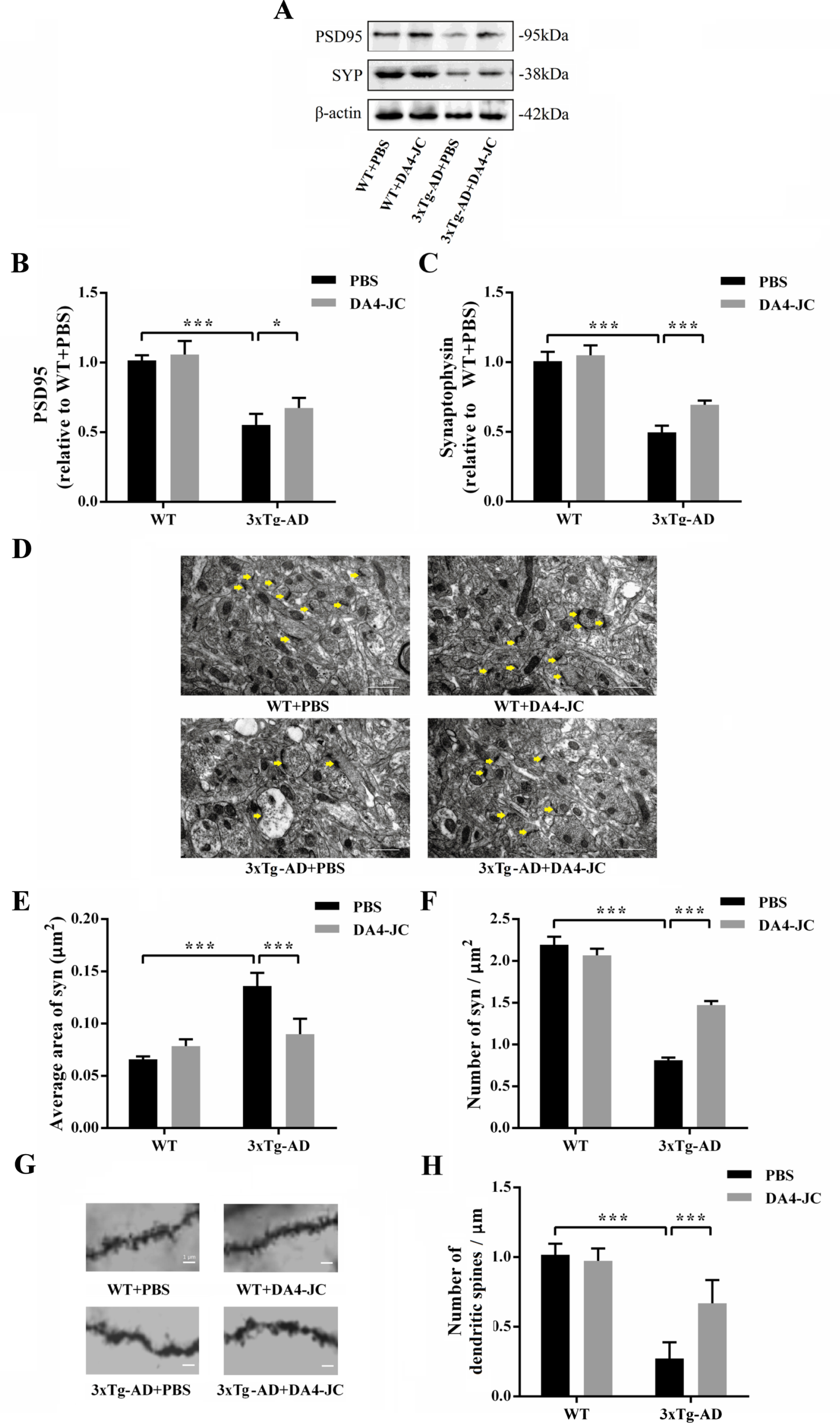
DA4-JC increased levels of PSD95 and SYP, and normalized synapse and dendritic spines numbers of 3xTg-AD mice. A) Representative western blotting images of PSD95, SYP, and β-actin in four groups. B, C) Histograms showed decrease in levels of PSD95 and SYP in PBS + 3xTg-AD mice was reversed by DA4-JC. D) Typical electron micrograph of synapse in four groups. The distribution of high electron density in synapse of hippocampus was uneven and synaptic structure was abnormal in 3xTg-AD + PBS mice. E) Histograms showed that average area of synapse was larger in 3xTg-AD + PBS mice, which was normalized by DA4-JC. F) Histograms showed that numbers of synapses were lower in 3xTg-AD + PBS mice, which was increased by DA4-JC. G) Typical micrograph of dendritic spines of hippocampus in four groups. H) Histograms showed that number of dendritic spines was less in 3xTg-AD + PBS mice, while DA4-JC prevented this reduction. *p < 0.05, ***p < 0.001, n = 6 for each group.
DA4-JC normalized synapse and dendritic spines numbers of 3xTg-AD mice
In Fig. 8E, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on average area of synapse of mice (genotype: F (1,20) = 93.49, p < 0.001; treatment: F (1,20) = 15.76, p = 0.0008; genotype×treatment: F (1,20) = 48.41, p < 0.001). Average area of synapse was larger in 3xTg-AD + PBS group (0.1360±0.0126, p < 0.001), compared with WT + PBS group (0.0658±0.0027), which was prevented by DA4-JC (0.0899±0.0148, p < 0.001). In Fig. 8F, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on number of synapse of mice (genotype: F (1,20) = 1248, p < 0.001; treatment: F (1,20) = 90.75, p < 0.001; genotype×treatment: F (1,20) = 197.9, p < 0.001). Number of synapses was less in 3xTg-AD + PBS group (0.8106±0.0341, p < 0.001) than WT + PBS group (2.1945±0.0950). After DA4-JC treatment, number of synapses was increased in 3xTg-AD mice (1.4716±0.0492, p < 0.001). In Fig. 8H, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on number of dendritic spines of mice (genotype: F (1,20) = 120.2, p < 0.001; treatment: F (1,20) = 13.61, p = 0.0015; genotype×treatment: F (1,20) = 21.11, p < 0.001). Compared to WT + PBS group (1.0172±0.0788), number of dendritic spines in 3xTg-AD + PBS group was less (0.2730±0.1161, p < 0.001). DA4-JC reversed the decreased number of dendritic spines in 3xTg-AD mice (0.6693±0.1655, p < 0.001).
DA4-JC alleviated hippocampal Aβ and tau pathology of 3xTg-AD mice
In Fig. 9C, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on Aβ area of mice (genotype: F (1,20) = 1062, p < 0.001; treatment: F (1,20) = 179.4, p < 0.001; genotype×treatment: F (1,20) = 151.5, p < 0.001). Aβ area was more in 3xTg-AD + PBS group (7.03%±0.38%, p < 0.001) than WT + PBS group (1.03%±0.17%). Aβ area was decreased in 3xTg-AD mice by DA4-JC treatment (3.59%±0.40%, p < 0.001). In Fig. 9D, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on p-tau area of mice (genotype: F (1,20) = 113.3, p < 0.001; treatment: F (1,20) = 32.09, p < 0.001; genotype×treatment: F (1,20) = 6.172, p = 0.022). Compared with WT + PBS group (11.62%±0.85%), p-tau area was more than that of 3xTg-AD + PBS group (17.38%±0.86%, p < 0.001). DA4-JC prevented the increased of p-tau area in 3xTg-AD mice (13.80%±0.73%, p < 0.001).
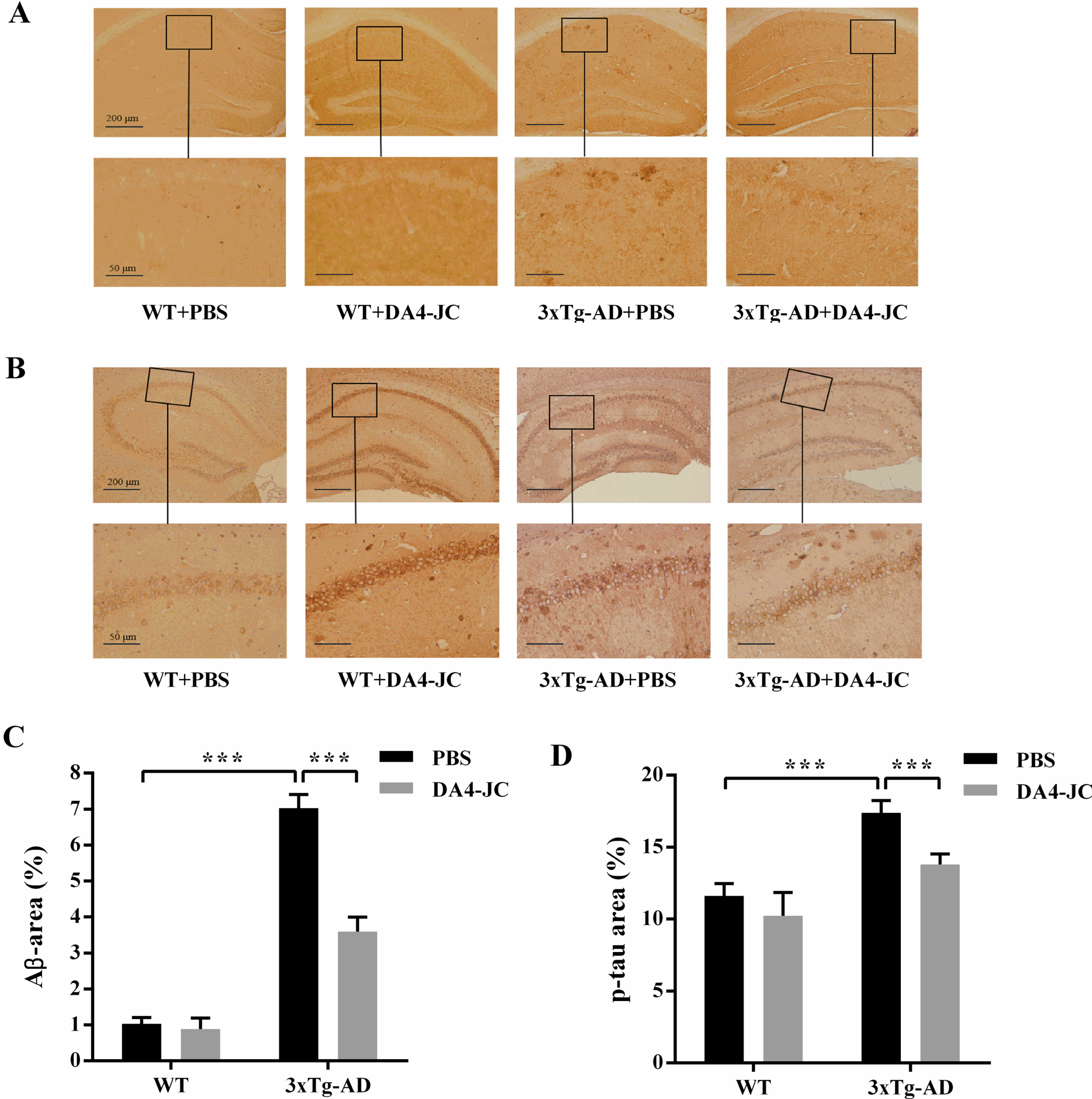
DA4-JC alleviated hippocampal Aβ and tau pathology of 3xTg-AD mice. A) Representative immunohistochemical images of amyloid plaques in four groups. B) Representative immunohistochemical images of neurofibrillary tangles in four groups. C) Histograms showed the percentage of Aβ area in hippocampus was significantly higher in PBS + 3xTg-AD mice than PBS + WT mice and a significant decrease was seen in DA4-JC + 3xTg-AD mice. D) Histograms showed the percentage of p-tau area in hippocampus was significantly higher in PBS + 3xTg-AD mice than PBS + WT mice and a measurable decrease was seen in DA4-JC + 3xTg-AD mice. ***p < 0.001, n = 6 for each group.
DA4-JC normalized mitochondria numbers of 3xTg-AD mice
In Fig. 10B, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on average volume of mitochondria of mice (genotype: F (1,20) = 92.52, p < 0.001; treatment: F (1,20) = 38.35, p < 0.001; genotype×treatment: F (1,20) = 5.856, p = 0.0252). Average volume of mitochondria was smaller in 3xTg-AD + PBS group (0.0810±0.0086, p < 0.001), compared to WT + PBS group (0.1416±0.0156). After DA4-JC treatment, average volume of mitochondria was larger in 3xTg-AD mice (0.1244±0.0111, p < 0.001). In Fig. 10C, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on number of mitochondria of mice (genotype: F (1,20) = 284.1, p < 0.001; treatment: F (1,20) = 27.42, p < 0.001; genotype×treatment: F (1,20) = 48.63, p < 0.001). Number of mitochondria in 3xTg-AD + PBS group was more (1.3379±0.1075, p < 0.001) than WT + PBS group (0.5766±0.0519). DA4-JC decreased number of mitochondria in 3xTg-AD mice (0.9479±0.0872, p < 0.001).

DA4-JC normalized mitochondria size and numbers of 3xTg-AD mice. A) Typical electron micrograph of mitochondria in four groups. Mitochondria of hippocampal neurons swelling and cavitation, and broken mitochondria cristae could be seen in 3xTg-AD + PBS mice. B) Histograms showed average volume of mitochondria was smaller in 3xTg-AD + PBS mice, which was normalized by DA4-JC. C) Histograms showed number of mitochondria was more in 3xTg-AD + PBS mice. ***p < 0.001, n = 6 for each group.
DA4-JC increased levels of PINK1 and Parkin and decreased P62 expression in 3xTg-AD mice
In Fig. 11B, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on PINK1 level of mice (genotype: F (1,20) = 568, p < 0.001; treatment: F (1,20) = 112.9, p < 0.001; genotype×treatment: F (1,20) = 51.65, p < 0.001). Level of PINK1 was decreased in 3xTg-AD + PBS group (0.24±0.05, p < 0.001), compared with WT + PBS group (1.00±0.07). DA4-JC boosted level of PINK1 in 3xTg-AD mice (0.68±0.04, p < 0.001). In Fig. 11C, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on Parkin level of mice (genotype: F (1,20) = 272.4, p < 0.001; treatment: F (1,20) = 21.38, p = 0.0002; genotype×treatment: F (1,20) = 49.82, p < 0.001). Level of Parkin was lower in 3xTg-AD + PBS group (0.32±0.05, p < 0.001) than WT + PBS group (1.00±0.09). Level of Parkin in 3xTg-AD mice was increased by DA4-JC treatment (0.66±0.07, p < 0.001). In Fig. 11D, two-way ANOVA showed that genotype and treatment had significant main effect and interaction on P62 level of mice (genotype: F (1,20) = 351.00, p < 0.001; treatment: F (1,20) = 10.13, p = 0.0047; genotype×treatment: F (1,20) = 41.30, p < 0.001). Compared to WT + PBS group (1.00±0.09), level of P62 in 3xTg-AD + PBS group was higher (1.69±0.04, p < 0.001). DA4-JC decreased level of P62 in 3xTg-AD mice (1.43±0.06, p < 0.001).

DA4-JC increased levels of PINK1 and Parkin and decreased P62 expression in 3xTg-AD mice. A) Representative western blotting images of PINK1, Parkin, P62, and GAPDH in four groups. B, C) A decrease in levels of PINK1 and Parkin in PBS + 3xTg-AD mice was reversed by DA4-JC. D) The increase in level of P62 in PBS + 3xTg-AD mice was reversed by DA4-JC. ***p < 0.001, n = 6 for each group.
DISCUSSION
Previous studies demonstrated that DA4-JC is more effective at the same dose than single receptor agonists lixisenatide and liraglutide, most likely because of co-stimulation of GLP-1 and GIP receptors [27]. By adding a poly-lys CPS, DA4-JC can readily cross the BBB at a much higher rate than liraglutide, semaglutide, or other dual agonists [28]. Higher expression levels of glial cell line-derived neurotrophic factor (GDNF), which promotes neuron survival, is induced by DA4-JC, compared to other dual receptor agonists [27, 29]. Therefore, we decided to test DA4-JC in the 3xTg-AD mouse model. Importantly, GLP-1, GIP, and GLP-1/GIP analogs do not promote insulin secretion when blood glucose level is in normal range and can be given to non-diabetic AD patients [30]. Our experiment showed that DA4-JC treatment did not affect body weight in both 3xTg-AD and WT mice, which was in line with the above point, suggesting that long-term treatment with DA4-JC is safe for AD patients without T2DM. DA4-JC showed clear improvements in a range of behavioral tests. In new object recognition test (ORT), spatial working memory test (YMT), spatial reference memory test (classical MWM test), cognitive flexibility test (reversal MWM test) and conditional fear memory test, 3xTg-AD mice were impaired, while DA4-JC treatment reversed these impairments.
Hippocampal LTP has been thought as a primary cellular mechanism, closely related to cognition [31]. We found that DA4-JC alone did not affect basic synaptic transmission, but DA4-JC treatment could significantly prevent hippocampal LTP deficit in 3xTg-AD mice. At the same time, we also tested synaptic proteins, such as post-synaptic density protein 95 (PSD95) and synaptophysin (SYP). PSD95, is a member of membrane-associated guanylate kinase and the major scaffolding protein in post-synaptic density (PSD) and a potent regulator of synaptic strength and plasticity [32]. SYP is an abundant membrane protein of synaptic vesicles enriched in presynaptic nerve terminals and regarded as an accurate index of neuronal synaptic density [32]. We found a reduction in expression levels of PSD95 and SYP in the hippocampus of 3xTg-AD mice, which was normalized by DA4-JC. In addition, numbers of synapses and dendritic spines were much reduced in 3xTg-AD mice, which were normalized by DA4-JC, too. Furthermore, treatment with DA4-JC markedly reduced levels of Aβ and p-tau in the hippocampus of 3xTg-AD mice. This may account for improvement of cognitive behavior in 3xTg-AD mice treated with DA4-JC.
Recent studies have found that autophagy plays a key role in AD [33]. Autophagy is essential for cell survival, and autophagy dysfunction is associated with neurodegenerative diseases [34]. Autophagy is impaired in AD [35]. Acting as a adaptor protein for autophagy, P62 level was increased when autophagy is inhibited [36], which was consistent with our experiment data, showing higher expression of P62 in 3xTg-AD mice than that of WT mice. Mitophagy, a type of selective autophagy, is a mechanism of selective removal of damaged mitochondria and timely removal of damaged mitochondria plays an important role in maintaining normal physiological function of cells [37]. Before cognitive deficit occurring in both AD patients and animal models, accumulation of dysfunctional mitochondria is a common feature of neurons in AD [38]. The study shows that impaired mitophagy can trigger accumulation of Aβ and tau protein, by aggravating oxidative damage, resulting in synaptic dysfunction and cognitive impairment [39]. Therefore, mitophagy is of great significance to AD [40]. At present, there are five main mitophagy pathways reported in mammals, and PINK1/Parkin pathway is the focus of current research [41]. Phosphatase and tensin homologue (PTEN) induced putative kinase 1 (PINK1) is a serine/threonine protein kinase, located in the outer membrane of mitochondria. Phosphorylation of PINK1 activates Parkin, and Parkin translocates from cytoplasm to mitochondria, resulting in promoting ubiquitination of mitochondrial outer membrane proteins and activation of ubiquitin-proteasome system (UPS), mediating autophagosomes coating mitochondrion, and then initiating mitochondrial autophagy [36]. Increasing expression of PINK1 and Parkin could promote clearance of damaged mitochondria in AD mice, alleviating synaptic loss and cognitive impairment [42, 43]. Our results displayed that DA4-JC could down-regulate p62 level and up-regulate PINK1/Parkin pathway, which well explained DA4-JC attenuating cognitive decline and pathology in 3xTg-AD mice. In addition, because of abnormal mitochondria division, average volume of mitochondria was smaller and the number of mitochondria was more in 3xTg-AD mice than that of WT mice, reflecting mitophagy dysfunction in AD, which was relieved by DA4-JC. The results found were in agreement with our previous findings, testing DA4-JC in different animal models of neurodegeneration. In 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) mouse model of PD, we found that DA4-JC protected the brain from the toxic effects of MPTP. Motor activity was improved, synaptic activity and dopamine synthesis normalized, and dopaminergic neurons were protected. Furthermore, chronic inflammation response observed in PD was much reduced by the drug. Importantly, expression of key growth factors such as GDNF was normalized, too [44]. In the streptozocin (STZ) icv rat model of AD, DA4-JC protected the brain from the toxic effects of STZ and improved spatial learning, reduced chronic inflammation response, reduced levels of phosphorylated tau, mitochondrial mitophagy signaling proteins expression such as BCL2-associated X protein (BAX) and B-cell lymphoma-2 (Bcl-2) were normalized, and improved insulin signaling as shown in protein kinase B (Akt) and insulin receptor substrate-1 (IRS-1) phosphorylation [25]. In APP/PS1 mouse model of AD, we found that DA4-JC was more effective than liraglutide in protecting memory formation, reducing amyloid plaque load, normalizing hippocampal LTP, and reducing levels of pro-inflammatory cytokines [23].
The present study adds important information on the underlying mechanism of the drug effects. We show for the first time that DA4-JC is protective in the 3xTg-AD mouse model and protects memory, normalizes synaptic numbers in hippocampus, size and numbers of mitochondria, and reduces levels of amyloid and phosphorylated tau in the brain. As the GLP-1 analogue liraglutide has already shown protective effects on cognition and memory and on the continuous shrinking of the brain in a phase II clinical trial of AD patients [17], and that DA4-JC is more effective than liraglutide [23] and can cross the BBB at a much enhanced rate compared to liraglutide [28], we conclude that DA4-JC has great potential to be more effective to treat AD patients than liraglutide. Clinical trials are planned to test the efficacy of DA4-JC in patients.
Footnotes
ACKNOWLEDGMENTS
This work was supported by National Natural Science Foundation of China (No. 31600865, 31700918); Open Fund from Key Laboratory of Cellular Physiology (Shanxi Medical University), Ministry of Education, China (No. KLMEC/SXMU-202013); Shanxi “1331 Project” Key Subjects Construction (1331KSC); Research Project supported by Shanxi Scholarship Council of China (No. 2020-083).