
Abstract
Background:
Women have a two-fold higher risk than men to Alzheimer’s disease (AD) at midlife. Larger brain tau burden was consistently shown in older women than age-matched men. The biological basis for this gender disparity remains elusive.
Objective:
We sought to know whether tau expression and phosphorylation physiologically differ between males and females.
Methods:
We used western blots and immunohistochemistry to compare the levels of total tau and phosphorylated tau in the hippocampus and entorhinal cortex (EC) between sexes in Wistar rats at 40 days, and 8 and 20 months of age.
Results:
We detected no statistically significant difference in total tau, 3R-tau, and 4R-tau between sexes. However, female rats exhibited lower levels of tau unphosphorylated at the Tau-1 site at 40 days of age. At 8 months of age, females showed higher levels of tau phosphorylated at Ser190, Ser387, and Ser395 (Ser199, Ser396, and Ser404 of human tau, respectively) than males in EC. At 20 months of age, both brain regions of female rats consistently showed higher levels than males of tau phosphorylated at Ser253, Ser387, PHF-1 (Ser387/395), and Ser413 sites, which correspond to Ser262, Ser396, Ser396/404, and Ser422 of human tau, respectively.
Conclusion:
Rats of both sexes have comparable levels of total tau, 3R-tau, and 4R-tau, whereas females exhibit higher levels of tau phosphorylated at multiple sites that are implicated in AD tau pathology, indicating a sexual dimorphism of tau phosphorylation that may potentially underlie the disparity in brain tau burden and risk for AD between sexes.
Keywords
INTRODUCTION
Sex and gender are considered genetic modifiers of biology and disease [1]. Owing to different levels of sex hormones along with their age-associated changes in the body and the male-specific Y-chromosome genes, men and women differ in many aspects of physiology and behavior, and exhibit disparity in many chronic diseases including neurodenegerative disorders [1, 2]. Sex differences are seen in prevalence, clinical manifestation, and treatment response in Alzheimer’s disease (AD), Parkinson’s disease, and amyotrophic lateral sclerosis [3–5].
As the most common cause of dementia, AD is a devastating, age-related neurodegenerative disorder that currently afflicts approximately over 50 million people worldwide, a figure that is projected to be 152 million by 2050 [6]. Whereas the etiology of sporadic form of AD, which represents over 95%of all cases, remains largely unknown, studies have identified a variety of genetic and environmental risk factors for the disease, which include aging, presence of apolipoprotein E type 4 allele (APOE ɛ4), Down syndrome, diabetes, and traumatic brain injury [7, 8]. Importantly, the disease appears to affect men and women with disparity. Epidemiological studies have consistently indicated that women exhibit a higher incidence of AD during aging and a two-fold higher lifetime risk for the disease than men at midlife [9–13]. Females also exhibit more rapid progression of mild cognitive impairment [14] and greater severity in clinical manifestation and deterioration of dementia [15, 16], along with more pronounced gray matter atrophy than men [17].
AD is histopathologically hallmarked by extracellular deposition of aggregated amyloid-β peptides as plaques and intraneuronal accumulation of aggregates of microtubule-associated protein tau as neurofibrillary tangles, accompanied by massive neuronal degeneration. Neurofibrillary tau pathology in AD brain is made up of hyperphosphorylated tau [18, 19], and this is also the case in all other tauopathies without exception [20, 21]. The burden of neurofibrillary tau pathology in the brain is positively corelated with the severity of dementia [22, 23], and an individual with familial AD who exhibited robust amyloid-β load did not show cognitive impairment until marked tau burden was detectable in the brain [24]. Thus, the accumulation of hyperphosphorylated aggregated tau is implicated in pathophysiological mechanisms of AD [25, 26].
The association of sex with tau pathology was suggested recently. Clinical neuroimaging studies consistently showed that older women, both clinically normal and cognitively impaired, exhibited heavier tau burden and higher tau accumulation rate in the brain than men [27–33]. These findings are in line with neuropathological studies indicating more robustly accelerated neurofibrillary and amyloid pathologies in women than men [34, 35]. Analysis of large data sets revealed that women showed higher levels of total tau in the cerebrospinal fluid (CSF) than men [35]. Among cognitively normal older women, APOE ɛ4 carriers showed ∼1.5-fold higher total tau level in the cerebrospinal fluid than APOE ɛ3 homozygotes, along with a significant reduction in functional brain connectivity; by contrast, their male counterparts showed minimal differences [36].
Gender difference in the expression and pathological alteration of tau has been implicated in transgenic mouse models of tauopathy. In two different transgenic mouse lines—JPNL3 and rTg4510—that overexpress P301L-mutated human tau by 1- and 13-fold higher than the endogenous murine tau, respectively, females consistently have higher levels of total tau and aggregated tau in the brain and develop tau pathology earlier than males [37, 38]. These studies suggest that gender may exert an effect on tau transgene expression and tau protein aggregation. However, it remains unknown whether females and males have different levels of brain tau under physiological conditions in wild-type animals that do not carry tau transgenes. In the present study, we set out to address this question by analyzing the levels of total tau and phosphorylated tau in the hippocampus and entorhinal cortex (EC) in wild-type rats at 40 days, and 8 and 20 months of ages.
MATERIALS AND METHODS
Animals
Wistar rats of both sexes at 40 days, and 8 and 20 months of age were used; these ages represent puberty, adult, and late midlife, respectively [39, 40]. Rats originally purchased from the Charles River Laboratories were bred and housed in local animal colony, with a 12/12 h light/dark cycle and ad libitum access to food and water. This study was carried out in accordance with the PHS Policy on Human Care and Use of Laboratory animals and approved by the Institutional Animal Care and Use Committee at New York State Institute for Basic Research in Developmental Disabilities.
Dissection of the hippocampus and EC
For biochemical studies, rats were euthanized by asphyxiation in a CO2 chamber and their brains were immediately taken out and submerged in ice-cold phosphate-buffered saline (PBS) until further dissection. Bilateral hippocampi and entorhinal cortices (Fig. 1A), the most vulnerable and the earliest brain regions affected by tau pathology in AD [41] and consistently shown to exhibit higher levels of tau burden in females in brain imaging [27, 43], were dissected in ice-cold PBS and separately stored at −80°C until used. Because the normal brain tau is rapidly dephosphorylated during postmortem delay and this can critically affect the assessment of tau phosphorylation [44], the time spent for asphyxiation and brain dissection were recorded. No statistically significant difference in postmortem interval was evident in tissue dissection between females and age-matched males (Fig. 1B, C).

Dissection of rat brain regions. A) Schematic diagrams showing entorhinal cortex and the hippocampus dissected for biochemical studies. B, C) Bar charts with scattered dots showing the time spent for brain dissection and total time spent for both asphyxiation and brain dissection, respectively. Data are expressed as mean ± SEM (n = 5-6 rats/group) and analyzed with unpaired Student t test. No statistical significance was detected between females and age-matched males. M and F represent males and females, respectively; this also applies to all the figures hereafter.
Preparation of brain homogenates
The hippocampus and EC tissues were homogenized in cold buffer consisting of 50 mM Tri–HCl, pH 7.2, 8.5%sucrose, 2.0 mM EDTA, 2.0 mM EGTA, 10 mM β-mercaptoethanol, 1.0 mM sodium orthovanadate, 100 mM sodium fluoride, 20 mM glycerol-phosphate, 50 nM okadiac acid, 5.0 mM benzamidine, 0.5 mM 4-(2-aminoethyl)benzenesulfonyl fluoride hydrochloride, and 10μg/ml of each of aprotinin, leupeptin, and pepstatin. The homogenate was mixed at 1:1 ratio (v/v) with 2× Laemmli buffer, followed by boiling for 5 min. Protein concentration was quantified by using the modified Lowry assay.
Western blotting
Western blotting was carried out as described previously [45]. Briefly, the same amount of brain homogenate protein was subjected to 10%–12.5%SDS-PAGE and electro-transferred onto the Immobilon-P membrane (Millipore, Bedford, MA). The membrane was sequentially blocked with 10%fat-free milk in Tris-buffered saline (TBS), incubated with primary antibody (Table 1), washed in 0.1%Tween-20 in TBS (TBST), incubated with species-matched peroxidase-conjugated secondary antibody (Jackson ImmunoResearch, West Grove, PA), and washed in TBST. The blots were then developed with an ECL kit (Pierce, Rockford, IL), and exposed to the HyBlot CL® autoradiography film (Denville Scientific, Holliston, MA). Specific immunoactivity was quantified by using the Multi Gauge V3.0 densitometric analysis software (Fuji Photo Film Co., Ltd.). The arbitrary units, with the background being subtracted, were used for the computation of relative immunoreactivity, which was a ratio calculated by dividing the arbitrary units of individual samples with the average of those for the age-matched males. The relative immunoreactivity was subjected to statistical analysis.
Primary antibodies employed in this study
All phosphorylation sites are numbered according to human tau441, i.e., the longest isoform of human brain tau. G, goat; M, mouse; Mono-, monoclonal; Poly-, polyclonal; R, rabbit.
Immunohistochemistry
For immunohistochemical studies, rats were deeply anesthetized by intraperitoneal injection of Avertin (600 mg/kg body weight), and transcardially perfused with PBS followed by 4%phosphate-buffered paraformaldehyde. The brain was carefully dissected out from the cadaver and submerged in the same fixative overnight at 4°C, dehydrated for cryopreservation in 30%sucrose, and cut into 40μm-thick, serial sagittal sections on a freezing microtome. Three sections, with 120μm intervals and at ∼1.6 mm laterally, were subjected to immunofluorescence staining. Brain sections of 40-day-old rats were immunostained with the antibody Tau-1 (1:1,000), and those of 20-month-old rats were probed with PHF-1 (1:1,000) and AT8 (1:1,000).
After blocked with 10%normal goat serum, sections were incubated with the primary antibody overnight at 4°C, washed in PBS, and then incubated with Alexa Fluor 555-conjugated goat anti-mouse IgG (1:1,000, Life Technologies, Rockford, IL) plus TO-PRO 3 iodide (1:1,000, Thermo Fisher Scientific, for nuclear counterstain) for 2 h at room temperature. Sections were then washed in PBS and mounted on pre-cleaned microscopic slides with anti-fading mounting medium. Omission of the primary antibody was included as a control procedure for nonspecific binding of the secondary antibodies. In addition, the specificity of tau antibodies were further verified by parallel immunostaining of brain sections from a tau knockout mouse [46].
The stained sections were viewed under a Nikon EZ C1 confocal microscope. Three random visual fields at the 100× objective lens for each section—in the CA1 stratum radiatum and the fimbria for the juvenile and 20-month-old rats, respectively–were scanned/photomicrographed at the best focal plane for morphometric analysis. The settings for the laser scanning, including power of the exciting laser, gain and scanning resolution, were kept constant throughout the acquisition of all fluorescent images. The images were then analyzed by using the NIH ImageJ for Windows. Signals were automatically thresholded with the Moments algorithm, and the area fraction was measured and expressed as percent area occupied. The measurements of all the images for each animal were averaged for subsequent statistical analysis.
Statistical analysis
The GraphPad Prism 8.0 software was used for statistical analysis. The Kolmogorov–Smirnov test was used to examine the normality of data distribution. Data were computed as mean±standard error of the mean (SEM), and analyzed with unpaired Student t test for the comparison between sexes at the same age when they passed both the normality test and the F test for equal variance, or otherwise with Welch’s corrected t test (for unequal variance) or non-parametric Mann-Whitney U test. p < 0.05 was considered statistically significant.
RESULTS
Male and female rats do not differ in the level of total tau or tau isoforms
A neuroimaging study reported that clinically normal women exhibited higher entorhinal cortical tau than men [27]. The expression level of tau plays an important role in its pathogenesis, and we recently found that the rat hippocampus and EC exhibit higher levels of total tau than other cortical and subcortical brain regions [45]. In the present study, we first asked whether the level of total tau in females differs from that in males. We probed the western blots of the hippocampal and EC tissues with 92e and 134d, two well-characterized rabbit polyclonal antibodies against total tau [47–49], and Tau-5, a commercially available mouse monoclonal pan-tau antibody. The results consistently showed no statistically significant difference between female and male rats across all three ages tested (Fig. 2).
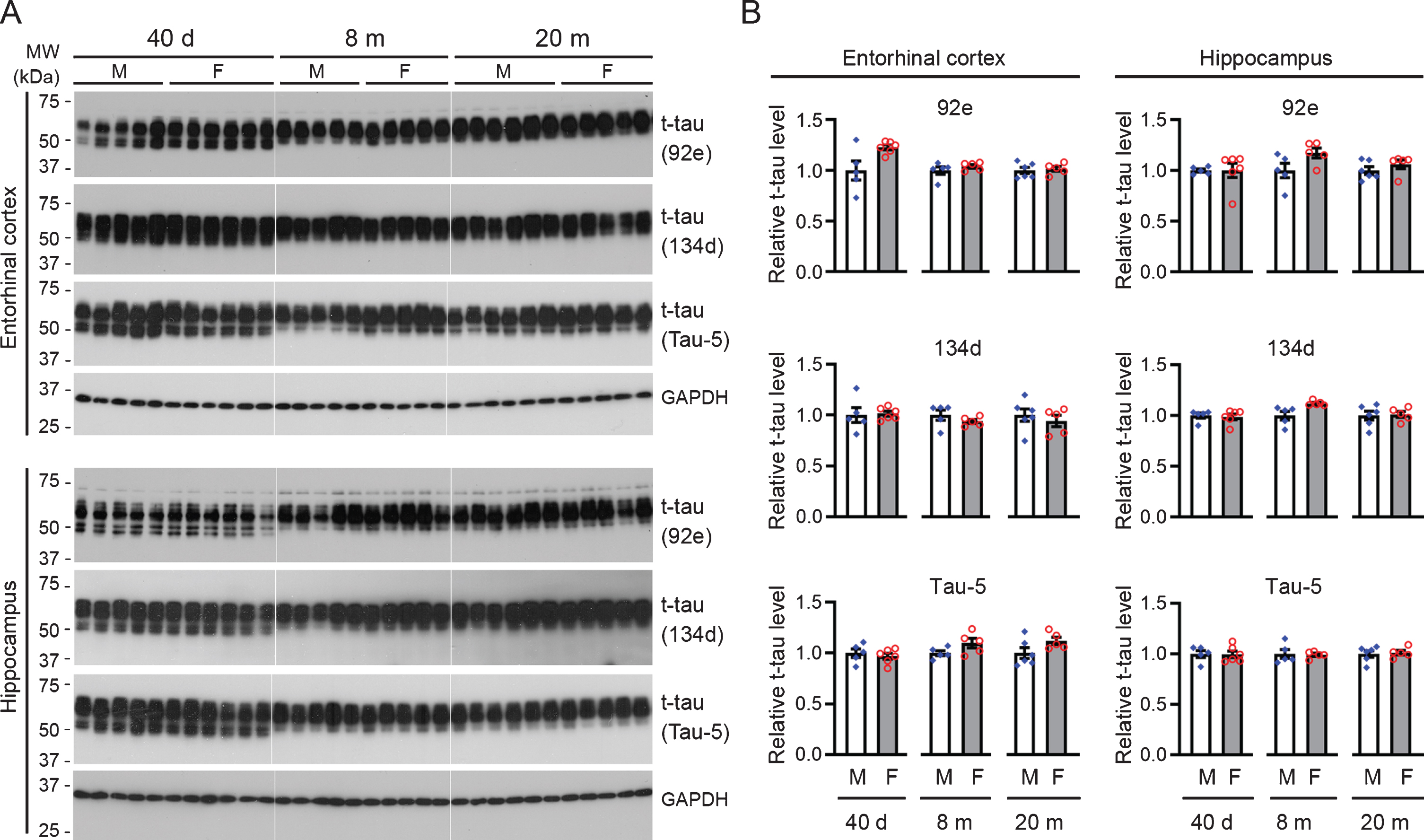
Male and female rats do not differ in total tau level in entorhinal cortex or in the hippocampus. A) Representative western blots probed separately with two rabbit polyclonal antibodies recognizing total tau (92e and 134d) and a mouse monoclonal antibody against total tau (Tau-5). GAPDH served as a loading control. B) Bar charts with scattered dots showing densitometric quantification. Data were converted as relative levels using the average of the males as 1, and this applies to the data presented in figures hereafter unless otherwise indicated. Data are expressed as mean ± SEM (n = 5-6 rats/group) and analyzed with unpaired Student t test. No statistical significance was detected between female and age-matched male rats.
Based on the presence or absence of the second microtubule-binding repeat domain of the molecule, tau is categorized into 4R-tau and 3R-tau isoforms, which are developmentally regulated and hold a balance under physiological condition in the adult brain [50–52]. Shift of the balance, toward an increase in 3R-tau for instance, may lead to the development of tau pathology [53]. In the present study, we examined the expression levels of 3R-tau and 4R-tau in the hippocampus and EC. We found that 3R-tau was robustly expressed in both the hippocampus and EC at 40 days of age and remained at a low level from younger adulthood to late midlife (Fig. 3). However, there was no detectable difference in 3R-tau, nor in 4R-tau, between female rats and age-matched males at all three ages tested, except for a trend of higher 4R-tau level in females than males in the hippocampus (p = 0.056) and EC (p = 0.062) at 20 months of age (Fig. 3). These data suggest that female and male rats have overall comparable levels of total tau, 3R-tau, and 4R-tau.
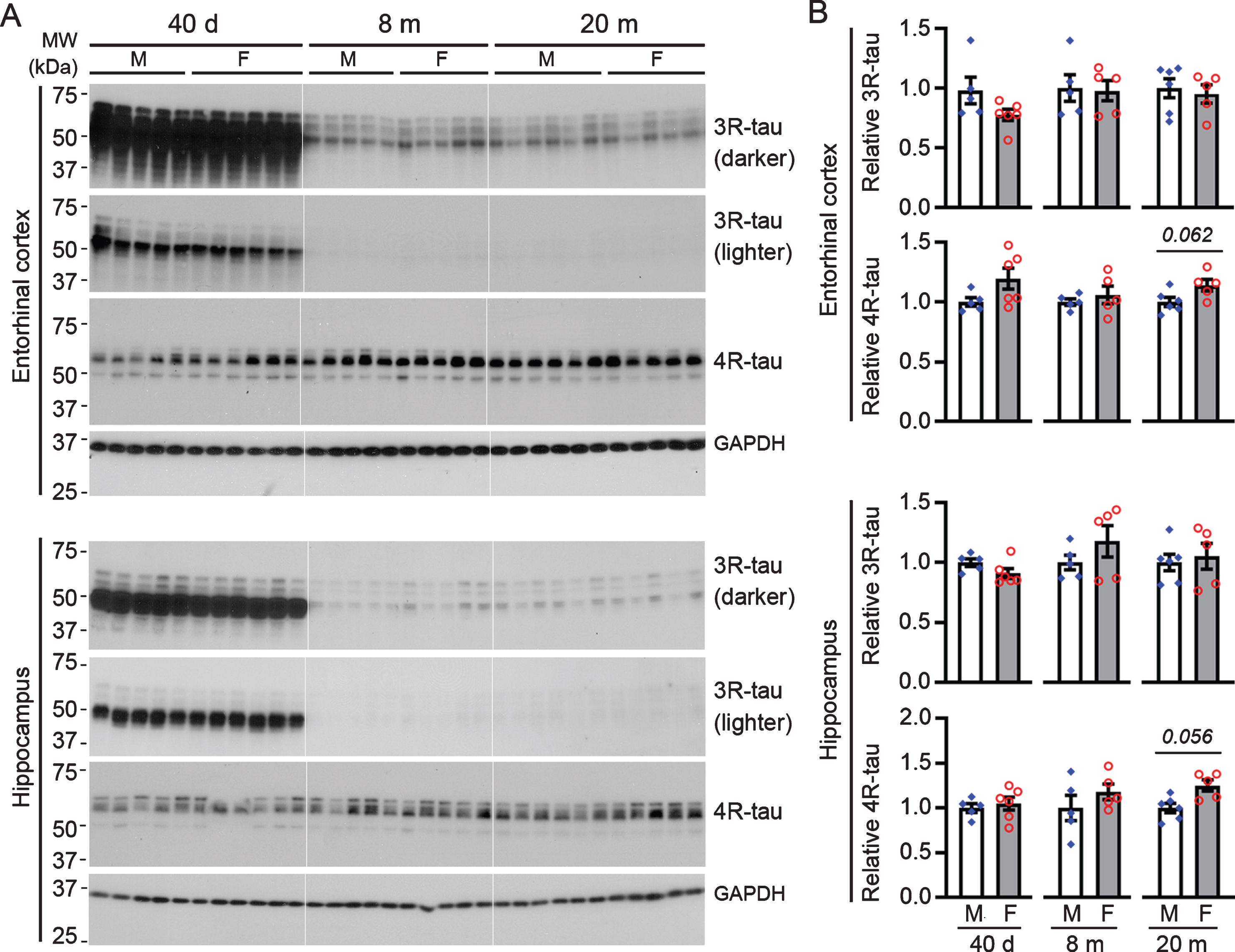
Male and female rats do not differ significantly in 3R-tau or 4R-tau levels in the hippocampus or entorhinal cortex. A) Representative western blots. B) Bar charts with scattered dots showing densitometric quantification. Data are expressed as mean ± SEM (n = 5-6 rats/group) and analyzed with unpaired Student t test. No statistical significance, except for a trend in 4R-tau at 20 months of age, was detected between female and age-matched male rats.
Female rats exhibit higher levels of phosphorylated tau than age-matched males
Tau pathology in all neurodegenerative tauopathies, including AD, is made up of hyperphosphorylated tau [20], and hyperphosphorylation of tau has been implicated in its aggregation [54–57]. Therefore, we asked whether female rats would have a higher level of phosphorylated tau than their male counterparts. To this end, we probed western blots with a set of phosphorylation-dependent antibodies that recognize specific tau epitopes only when they are phosphorylated at a particular site(s), except for Tau-1 that recognizes tau unphosphorylated at Ser186/189/190/193 of rat tau, which corresponds to Ser195/198/199/202 of human tau.
At 40 days of age, rats of both sexes exhibited comparable levels of tau phosphorylated at Ser190, Thr208, Ser253, Ser387, Ser395, and Ser413 in both EC and hippocampus. However, female rats at this age showed a markedly lower level of tau recognized by Tau-1 (Figs. 4 5). These data suggest that female young rats had a higher level of tau that is phosphorylated at Ser186/189/190/193 than the male animals.
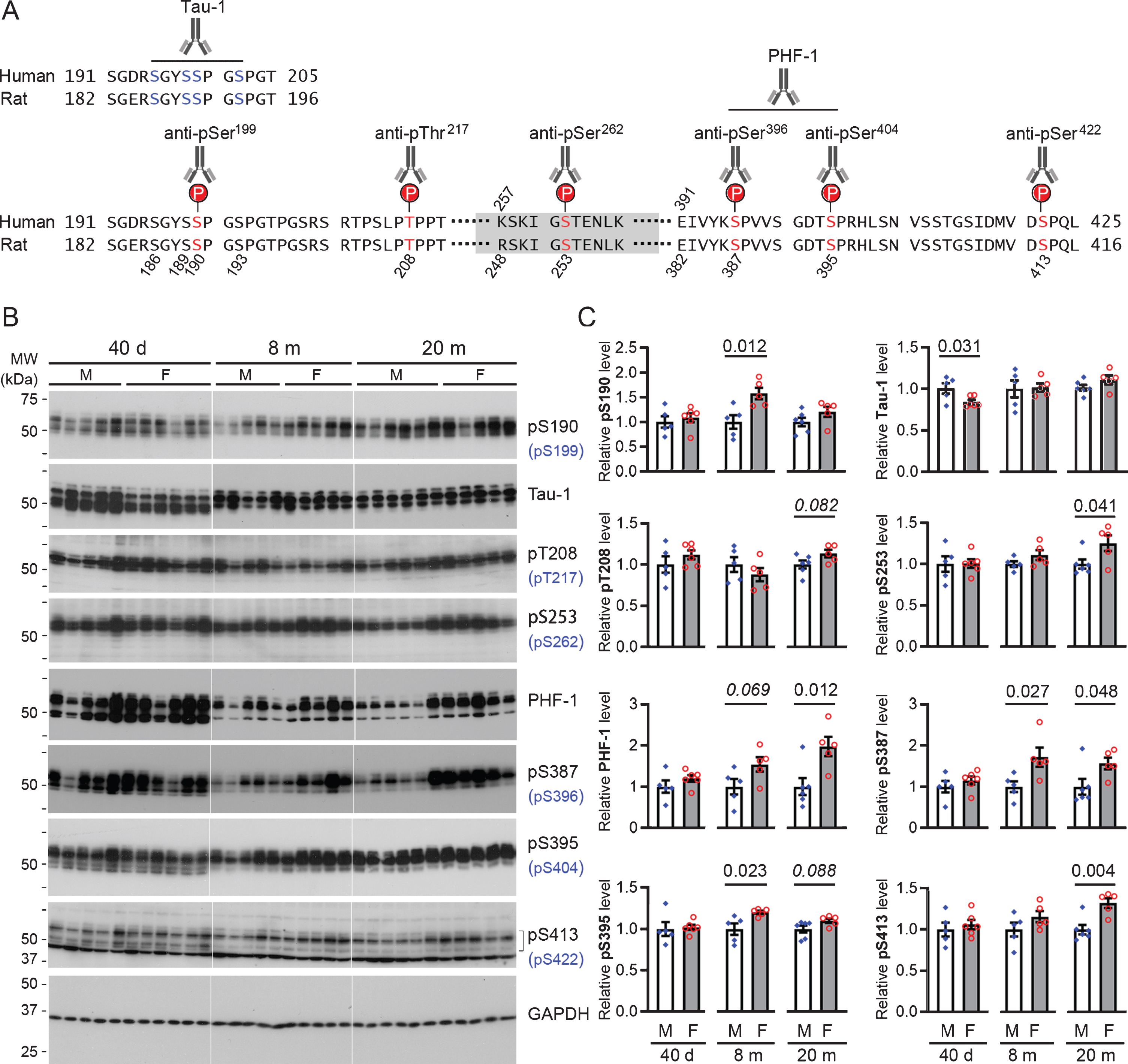
Female rats show higher levels of phosphorylated tau at multiple sites in entorhinal cortex than their male counterparts. A) Schematic diagrams showing the epitopes recognized by phosphorylation-dependent tau antibodies used for western blots. Relevant regions of the amino acid sequences of the longest isoforms of human brain tau (tau441) and rat brain tau (432 amino acid long) are aligned to depict the epitopes. B) Representative western blots probed with indicated phospho-tau antibodies. Epitopes shown in brackets are the equivalent epitope on human tau. C) Bar charts with scattered dots showing densitometric quantification. Data are expressed as mean ± SEM (n = 5-6 rats/group) and analyzed with unpaired Student t test. The p-value is shown wherever a statistical significance or a trend (italic) was detected; this also applies to all the figures hereafter.
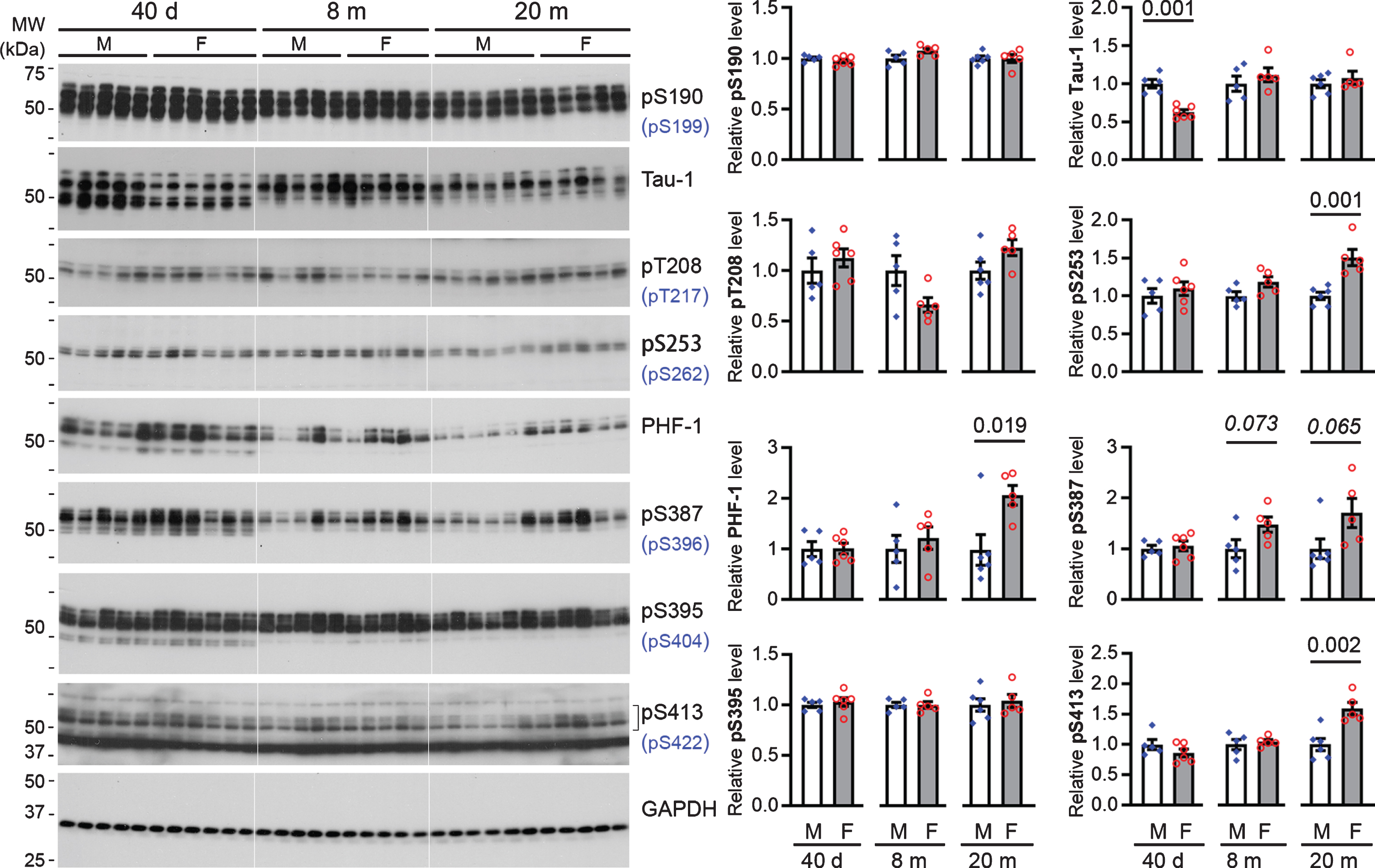
The hippocampus of female rats exhibits higher levels of tau phosphorylated at multiple sites than their male counterparts. A) Representative western blots probed with indicated anti-phospho-tau antibodies. Epitopes shown in brackets are the equivalent epitope on human tau. B) Bar charts with scattered dots showing densitometric quantification. Data are expressed as mean ± SEM (n = 5-6 rats/group) and analyzed with unpaired Student t test.
At 8 months of age, the EC in female rats was found to have moderately higher levels of tau phosphorylated at Ser190, Ser387, and Ser395−corresponding to Ser199, Ser396, and Ser404 of human tau, respectively−than in male rats (Fig. 4); only a similar trend was seen for Ser387 (p = 0.073) phosphorylated tau in the hippocampus (Fig. 5). However, when rats reached the age of 20 months, both EC and the hippocampus of female rats consistently showed higher levels of tau phosphorylated at Ser253, Ser387, PHF-1 (Ser387/395), and Ser413, which correspond to Ser262, Ser396, Ser396/404, and Ser422 of human tau, respectively, than male animals; a similar trend was also observed for tau phosphorylated at Thr208 and Ser395 (p = 0.082 and 0.088, respectively) in the EC (Figs. 4 5).
We next performed immunofluorescence staining of rat brain sections at two representative ages, 40 days and 20 months (Fig. 6A-E), to verify the main findings of the western blot analysis. The antibodies used for western blots were screened, by using brain sections from a tau knockout mouse, for their specificity in immunohistochemistry, and three monoclonal antibodies that showed no detectable immunoreactivity in tau null mouse brain, Tau-1, PHF-1, and AT8, were selected (Fig. 6F).
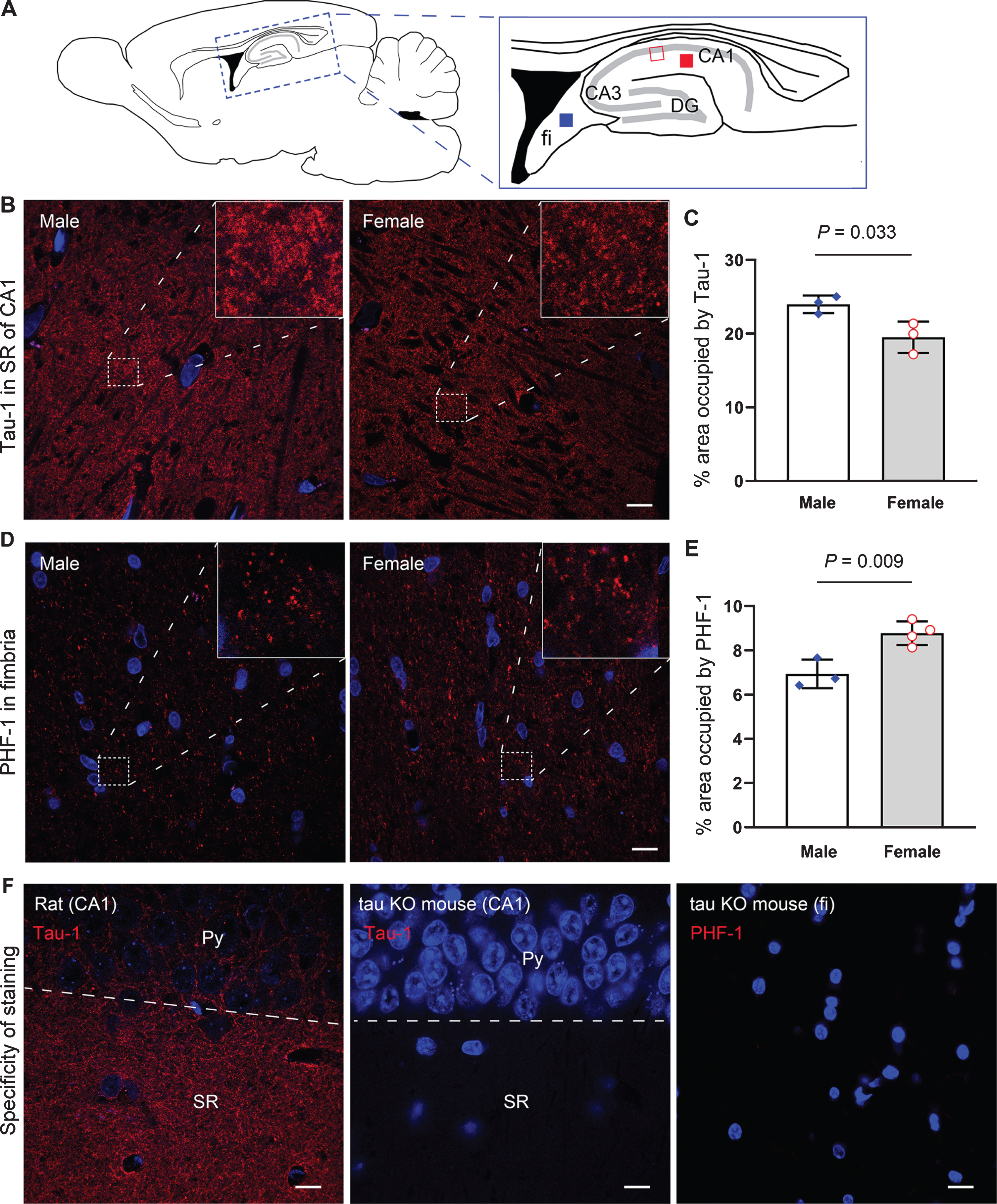
Immunofluorescence staining showing higher levels of phosphorylated tau in the hippocampus of female than aged-matched male rats. A) Schematic diagrams showing the locations for photomicrographs and morphometric analysis. The red and blue filled squares and the red open square indicate the areas shown in (B), (D), and (F), respectively. B, C) Representative photomicrographs and quantification of Tau-1 immunostaining in the CA1 stratum radiatum (SR) in female versus male rats at 40 days of age, respectively. D, E) Representative photomicrographs and quantification of PHF-1 immunostaining in the fimbria (fi) in female versus male rats at 20 months of age. F) The specificity of antibodies used. Neither of Tau-1 and PHF-1 exhibited specific staining in tau knockout (KO) mouse brain. TO-PRO 3 iodide (blue) was used to counterstain the nuclei of cells. Data are expressed as mean ± SEM (n = 3-4 rats/group) and analyzed with unpaired Student t test. Scale bar = 10μm. Py, pyramidal cell layer.
Tau-1 immunoreactivity was seen to abundantly distribute in the neuropil areas of the rat brain (Fig. 6B). Morphometric analysis showed that 40-day-old female rats exhibited ∼19%less Tau-1 immunoreactivity in the stratum radiatum of the hippocampal CA1 subfield as compared to males at the same age (Fig. 6C). PHF-1 immunostaining signals, however, was found to sparsely distribute in fimbria and CA3 mossy fiber area (Fig. 6D). Quantification of the PHF-1 staining showed ∼26%more immunoreactivity in fimbria in female rats than male animals at 20 months of age (Fig. 6E). No specific immunoreactivity to AT8 was detected in rat brain sections by immunofluorescence staining.
Male and female rats do not differ in levels of gliosis or presynaptic markers
Synaptic changes and gliosis are common features in neurodegeneration [58, 59]. To test whether the higher level of phosphorylated tau in 20-month-old females was related to synaptic degeneration and gliosis, we performed western blotting with antibodies recognizing synaptophysin (a presynaptic marker), glial fibrillary acid protein (GFAP), an astroglial marker, and ionized calcium-binding adaptor molecule 1 (Iba-1), a marker of residential microglia. No statistically significant difference between female and male rats across all three ages was detected for these three markers, despite a modest increase in GFAP expression in 20-month-old rats compared to those at younger ages and a slightly lower level of Iba-1 in 8-month-old females than age-matched males (Fig. 7).

Female rats show similar levels of glial and presynaptic markers to their male counterparts. A) Representative western blots probed with antibodies against indicated markers. B) Bar charts with scattered dots showing densitometric quantification. Data are expressed as mean ± SEM (n = 5-6 rats/group) and analyzed with unpaired Student t test. No statistical significance, except for Iba-1 at 8 months of age, was detected between female and age-matched male rats. Syp, synaptophysin.
DISCUSSION
The biological basis for higher brain levels of pathological tau in old females than their male counterparts remains to be understood. Here we found, for the first time, that female rats exhibit higher levels of phosphorylated tau at multiple sites than age-matched males, despite their comparable levels of total tau, 3R-tau, and 4R-tau. These findings add to mechanistic insights into the disparity between males and females regarding the tau pathology load in the aging brain.
The level of total tau is important in tau pathogenesis; reduction in tau expression was shown to reverse tau deposition in mouse models of tauopathy [60, 61]. The expression levels of total tau appear to be an important determinant of the regional susceptibility of the human brain to the development of tau pathology, which is evidenced by the relative sparing of the cerebellum [62] that conspicuously exhibits the lowest level of total tau among other brain regions [45, 63]. In the present study, we used well-characterized antibodies against total tau in western blots and found that there was no detectable difference in total tau levels in the hippocampus and EC between male and female rats. In addition, we measured the levels of 3R-tau and 4R-tau, imbalance of which is also implicated in tau pathogenesis [53], and the results showed no statistically significant difference between the sexes, except for a trend of more 4R-tau in late midlife females. Given that the adult rat brain expresses predominantly 4R-tau [64], a similar trend in total tau would be expected albeit not evident in the present study. This could be the consequence of a relatively small sample size, or rather small difference in total tau between sexes. Nevertheless, our data suggest that physiologically, male and female rats may have overall comparable levels of total tau and its alternative splicing isoforms from puberty to late midlife.
Tau can be modulated by a variety of post-translational modifications, among which phosphorylation appears to be involved at the largest number of sites on the tau molecule [65, 66]. Indeed, tau is a soluble phosphoprotein that can be phosphorylated at 85 putative phosphorylation sites—45 serine, 35 threonine, and 5 tyrosine residues—on its longest isoform in the human brain [65]. Among these sites, two types pertaining to the microtubule-binding activity of tau, the primary function of tau identified in vitro, have been mostly studied [67]. The first one is the KXGS motifs in the microtubule-binding repeats (Ser262/324/356), phosphorylation at which, particularly at Ser262, strongly inhibits tau-microtubule interaction; the second type is the Ser/Thr-Pro motifs within the repeat-flanking domains, phosphorylation at which also alters the affinity of tau for microtubules albeit at a lesser extent [67–69]. In the present study, we analyzed both Ser253 (equivalent of Ser262 for human tau441) and the main proline-directed Ser/Thr sites, i.e., Ser190, Ser193, Thr208, Ser387, Ser395, and Ser413, which correspond to Ser199, Ser202, Thr217, Ser396, Ser404, and Ser422 of human tau411, respectively. Phosphorylation of tau on these sites was reported in both developing and adult rat brains [70], and the human tau protein is hyperphosphorylated at all these sites in AD tau pathology [71, 72].
In the brain of young adult (10 weeks of age) rats, tau was reported to be slightly to moderately phosphorylated at Thr172, Ser190, Ser193, Thr222, and Ser395, whereas the vast majority of adult rat tau was found unphosphorylated at both Ser387 and Ser391 [70]. In the present study, we detected a significant amount of tau phosphorylated at Ser387 at 40 days, and at 8 and 20 months of age, with a higher level in adult females than males. This discrepancy may arise from different ages of animals tested, as old rodents exhibit higher levels of phosphorylated tau than young adults, including those at the site Ser387 that corresponds to Ser396 of human tau [73, 74]. These findings are in line with a previous study showing higher phospho-tau levels in females than males in P301L-tau transgenic mice [75].
Tau phosphorylation is developmentally regulated [70, 76] and exhibits regulatory roles in neurodevelopment, synaptic activity and other physiological functions [77–79], probably by dynamically altering its binding activity to microtubules or interaction with Fyn and PSD-95/NMDA receptor complex [67, 79]. Hyperphosphorylation of non-mutated tau was shown to induce its self-aggregation in vitro [54, 57], to increase its propensity of being captured by pathological tau seeds [55], and to directly initiate its aggregation and insolubilization in the brain of adult wild-type mice [80]. In addition, a meta-analysis of transcriptome-wide interaction studies has linked the higher risk for AD in women than men with tau phosphorylation process [81]. The absence of AT8 immunoreactivity in immunohistochemical staining at 20 months of age in rats of both sexes indicates a non-aggregated state of the tau protein at midlife in Wistar rats. However, the higher levels of phosphorylated tau in females than males may endow the brain tau in old females with a higher propensity of undergoing pathological alterations under certain unfavorable or disease conditions, for instance, being recruited in the presence of seeding-competent oligomeric tau [55].
The sex-phosphorylated tau dimorphism may be attributed to different levels of sex hormones between males and females [82]. Lower testosterone level was found to be related to higher CSF phospho-tau level among APOE ɛ4 carriers regardless of sex [83]. Intriguingly, the effect of estrogen on tau phosphorylation appears to depend on the differentiation state of neurons [31, 85], as the hormone per se could exert influence on neuronal differentiation [86, 87]. Treatment of mature neurons differentiated from SH-SY5Y neuroblastoma cells with 17-β-estradiol significantly increased tau phosphorylation [85]. Nonetheless, the exact mechanisms underlying the sex dimorphism regarding phosphorylated tau remained to be explored in future studies.
The increase in GFAP expression is an indication of astrogliosis associated with aging [88]. In the present study, we found that rats at 20 months of age exhibited a moderate yet consistent increase in GFAP expression in both EC and the hippocampus as compared to 8-month-old animals. Although the age of 20 months in the commonly used strains of laboratory rats is usually considered the late midlife related to their lifespan [40, 89], the increased GFAP expression in rats of both sexes suggests that aging-associated changes may have started at this age in Wistar rats. Nonetheless, the absence of statistically significant difference in GFAP expression between sexes at the same age indicates that the sexual dimorphism in phosphorylation of brain tau may not be associated with differential levels of astrogliosis at 20 months of age. Likewise, no increased Iba-1 expression was detected in 20-month-old females, suggesting that there might not be microgliosis associated with higher level of phosphorylated tau in female rats at this age.
In short, we found that the hippocampus and EC exhibit higher levels of phosphorylated tau in female than male wild-type rats, despite their comparable levels of total tau and its alternative splicing isoforms. This sex dimorphism may predispose females to greater susceptibility than males to the development of tau pathology.
Footnotes
ACKNOWLEDGMENTS
This work was supported by funds from the New York State Office for People with Developmental Disabilities and the Alzheimer’s Association (2016-NIRG-397030). We thank Drs. Lester I. Binder and Peter Davies for their generous gifts of Tau-1 and PHF-1 antibodies, respectively. We also thank Ms. Yunn Chyn Tung for her technical support in western blotting.