
Abstract
Background:
Geriatric surgical patients are at higher risk of developing postoperative neurocognitive disorders (NCD) than younger patients. The specific mechanisms underlying postoperative NCD remain unknown, but they have been linked to genetic risk factors, such as the presence of APOE4, compared to APOE3, and epigenetic modifications caused by exposure to anesthesia and surgery.
Objective:
To test the hypothesis that compared to E3 mice, E4 mice exhibit a more pronounced postoperative cognitive impairment associated with differential DNA methylation in brain regions linked to learning and memory.
Methods:
16-month-old humanized apolipoprotein-E targeted replacement mice bearing E3 or E4 were subjected to surgery (laparotomy) under general isoflurane anesthesia or sham. Postoperative behavioral testing and genome-wide DNA methylation were performed.
Results:
Exposure to surgery and anesthesia impaired cognition in aged E3, but not E4 mice, likely due to the already lower cognitive performance of E4 prior to surgery. Cognitive impairment in E3 mice was associated with hypermethylation of specific genes, including genes in the Ephrin pathway implicated in synaptic plasticity and learning in adults and has been linked to Alzheimer’s disease. Other genes, such as the Scratch Family Transcriptional Repressor 2, were altered after surgery and anesthesia in both the E3 and E4 mice.
Conclusion:
Our findings suggest that the neurocognitive and behavioral effects of surgery and anesthesia depend on baseline neurocognitive status and are associated with APOE isoform-dependent epigenetic modifications of specific genes and pathways involved in memory and learning.
Keywords
INTRODUCTION
Adults aged sixty-five years and older are the fastest-growing segment of the United States population, and this population accounts for a large portion of all surgeries performed in the United States each year. The development of a postoperative neurocognitive disorder (NCD) is one of the most common postoperative complications in this age group [1, 2]. Recent large epidemiological studies and prospective longitudinal investigations have shown that these older patients are also at higher risk of developing Alzheimer’s disease (AD) [3 –6]. Animal and biomarker investigations also suggest a link between exposure to surgery and anesthesia and AD pathology [7 –14]. However, there is conflicting literature around the hypothesis that surgery and anesthesia lead to an increased risk of AD [15]. Some suggest it is unclear whether exposure to anesthesia and surgery per se increases the risk of developing AD, whether it exacerbates existing co-morbidities that predispose to AD, or if the association simply reflects the natural cognitive trajectory of patients undergoing these procedures.
While the etiology of AD is not fully understood, environmental factors are thought to interact with genetic factors to modulate its development and progression; one such environmental factor may be exposure to surgery and anesthesia. However, the mechanisms underlying the development of postoperative NCD after surgery and anesthesia are uncertain.
Epigenetic mechanisms, chemical modification of the DNA and chromatin that do not affect the DNA sequence are responsible for modulating gene expression in different stages of development, including aging. In particular, alterations of DNA methylation have been implicated in several neurodegenerative diseases, including AD [16]. Although exposure to surgery and anesthesia is associated with postoperative NCD in older adults and causes memory impairment in aged rodent models [17, 18], there is little known regarding the role of DNA methylation in postoperative NCD. Moreover, whether surgery and anesthesia alter DNA methylation globally or in a more targeted way is unclear.
One gene associated with both AD and postoperative NCD is apolipoprotein E (APOE) [19 –21]. In humans, there are three major APOE isoforms: E2, E3, and E4. Compared to E3, E4 increases the risk of developing AD and NCD in older adults [22 –25]. However, as mentioned above, the long-term cognitive decline after surgery in APOE4 carriers versus non carriers may simply reflect the natural cognitive trajectory in APOE4 carriers, or the effects of comorbidities on cognition [26, 27]. In the current study, we used an animal model to test the hypothesis that exposure to surgery and anesthesia causes a more pronounced cognitive injury in aged E4 mice than aged E3 mice. Therefore, we tested the hypothesis that exposure to surgery and anesthesia causes a more pronounced cognitive injury in aged E4 mice than aged E3 mice. We further explored whether differences in postoperative neurocognitive injury in the E3 and E4 mice were associated with specific differences in DNA methylation patterns in the hippocampus, a brain region associated with learning and memory that is heavily affected in AD [28].
MATERIALS AND METHODS
Animals, anesthesia, and surgery
All experiments complied with the NIH Guide for the Care and Use of Laboratory Animals, and protocols were approved by Oregon Health & Science University Animal Care and Use Committee.
Sixteen-month-old male and female humanized targeted replacement mice expressing E3 or E4 under control of the mouse APOE promoter were assigned to either surgery + general anesthesia (n = 24 mice; 12 mice/genotype) or sham procedure (n = 24 mice; 12 mice/genotype) [29, 30]. There was an equal distribution of males and females in each group. All mice were singly housed prior to random assignment to surgery or sham group. Before surgery, the mice were positioned on a heating pad (37°C) using paper tape. Mice were actively warmed using a water circulation heating blanket, and temperature, respiration rate, and mucous membrane color were monitored throughout surgery. Isoflurane was administered via nose cone at 4% concentration for induction and 1.5–2% in air during the surgical procedure. The surgery, a laparotomy, was performed under sterile conditions and consisted of opening and suturing the abdomen closed under 30 min of general isoflurane anesthesia. Adequacy of anesthesia was determined by the absence of response to toe pinch or absence of blink reflex. For postoperative pain control, mice were injected with 0.05 mg/kg buprenorphine immediately postoperatively, and twice daily for 3 days postoperatively. Mice were provided acetaminophen-treated drinking water for 24 h in advance of the surgical procedure and for 3 days postoperatively. Control mice were handled similarly but did not undergo surgery and anesthesia and did not receive buprenorphine or acetaminophen (Fig. 1).

Mice were randomized into two groups. The sham mice were exposed to oxygen and did not receive surgery. The surgery + general anesthesia group underwent a laparotomy and thirty minutes of isoflurane general anesthesia. This figure was created with BioRender.com.
Beginning on postoperative day 4 or 5, animals were tested in the open field for 2 days, followed by a 2-day novel object recognition paradigm. In the second week, animals were tested in a 5-day water maze paradigm. During the third and final week of testing, animals underwent fear conditioning habituation on Tuesday, contextual and cued fear conditioning recall on Wednesday, and were euthanized on Thursday (Fig. 2) via cervical dislocation. Brains were quickly removed and dissected, isolating hippocampi as described by Spijker [31]. Dissected tissues were immediately frozen in liquid nitrogen and stored at –80°C for DNA methylation analysis.
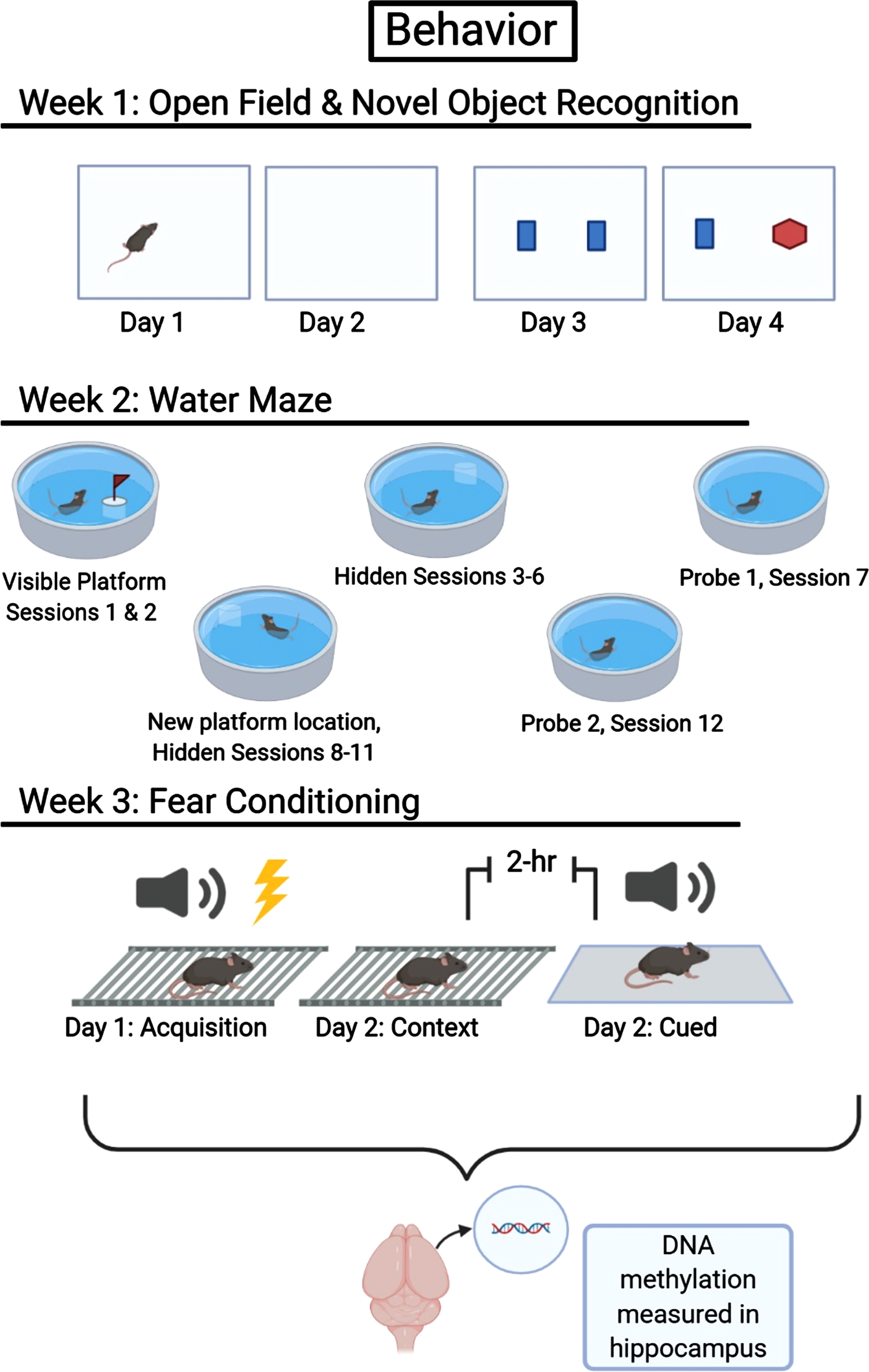
Timeline for behavioral testing and DNA methylation analysis. This figure was created with BioRender.com.
Behavioral testing
Open field and novel object recognition
The open-field test is used to assay general locomotor activity levels, anxiety, and exploratory behavior. Mice were placed in an open field (41 cm×41 cm, white walls and floors) for 10 min/day, on 2 consecutive days. Total distance moved and time spent in the center of the arena (20 cm2) were tracked and analyzed using Ethovision XT software (Noldus, Wageningen, the Netherlands). The time spent avoiding the more anxiety-provoking center of the open field was used to determine anxiety-like behavior.
The novel object recognition test is used to investigate aspects of learning and memory. Hippocampal function is important in this novel object recognition task that includes a 24 h interval between training and testing [32]. Following habituation to the open field, on the third day, mice were exposed to 2 identical objects (small orange hexagonal prisms, 5 cm×3 cm) placed in the center of the arena, 10 cm apart. On day 4, one of the familiar objects was replaced with a novel object of distinct shape and color (small green triangular prism). On both days 3 and 4, mice were given 15 min to explore the arena containing the objects. Clear visuospatial orientation to the object within 2 cm proximity and physical interaction with the object was coded as exploratory behavior. A researcher blinded to the treatment and genotype of the mice conducted all behavioral analyses. The time spent interacting with each object was analyzed, and the percent time spent exploring the familiar and novel object was used to determine novel object recognition.
Water maze
The Morris water maze tests a rodent’s spatial learning and memory using the location of a hidden escape platform with the aid of visual cues located around a pool. The pool is 140 cm in diameter and is filled with water made opaque with white chalk. The water is allowed to sit overnight to reach room temperature, 22±2°C.
In this 5-day paradigm, mice underwent 1 day of visible platform training followed by 4 days of hidden platform training. The training consisted of 2 sessions per day (AM and PM) with 2 trials per session, totaling 4 trials per day. Trials were separated by 10 min, and sessions were separated by 3 h. On the first day, mice underwent “visible” platform trials, in which a colored flag clearly marked the submerged escape platform. The following 4 days consisted of “hidden” platform trials, for which the flag was removed from the submerged platform. During sessions 3–6, the platform remained in the same location, and visual cues on the curtains surrounding the pool served as geographic markers for navigation to the platform location. The platform was moved to a new target location following the first probe trial and remained in the same location during sessions 7–10.
Probe trials, 1 min trials with no platform, were used to assess the memory of the trained mice. The first probe trial occurred post-training in the afternoon of day 4, and the final probe trial occurred post-training on day 5.
Contextual and cued fear learning and memory
Classical fear conditioning is an associative learning task that assesses fear response to a neutral conditioned stimulus (a tone) paired with an aversive unconditioned stimulus (a shock). Freezing in rodents, defined as cessation of body movement, is a defensive behavior expressed in response to the impending threat. During the training phase of the fear conditioning test, mice were placed inside a dark fear-conditioning chamber, chamber lights were turned on at time zero (100 lux), and mice were given 2 min to habituate. This baseline was followed by a 30 s tone (80 db at 2800 Hz), co-terminating with a 2 s 0.35 mA foot shock, repeated once more during the training trial. Between each tone-shock pairing, there was a 90 s inter-stimulus interval. Chambers were cleaned with 0.5% acetic acid between trials. Fear acquisition was analyzed by quantifying the percent time spent freezing during the baseline, tone, and shock periods using Video Freeze automated scoring software (Med Associates Inc., St. Albans, VT).
The following day, the association of the environment (hippocampus-dependent contextual memory test) with the unconditioned stimulus (shock) was determined by placing animals back in the same chamber at the same time of day for 5 min and measuring freezing behavior. Three hours later, mice were exposed to a modified environment to assess the learned association between the unconditioned stimulus and conditioned stimulus. The environment of the chamber was altered by placing white plastic floor covers over the bottom of the chamber, covered by black triangular plastic inserts. The scent of the chamber was altered using a vanilla-soaked cotton nestlet attached to the top of the plastic inserts, and 10% isopropanol was used for cleaning the enclosure between trials. During the hippocampus-independent cued memory test, there was a 90 s baseline followed by a 180 s tone (80 db @ 2800 Hz), and this series was repeated a total of 5 times. Fear memory was measured as the percent time spent freezing in response to the environment (contextual) or tone (cued). The cued fear memory test is used to assess the degree to which the conditioned stimulus, the tone, is associated with the expectation of the unconditioned stimulus, the shock.
DNA methylation analysis
Reduced representation bisulfite sequencing library generation
Genomic DNA was extracted from the mouse hippocampi using the Qiagen All prep kit, and reduced representation bisulfite sequencing (RRBS) libraries were generated from 200 ng of hippocampal genomic DNA. Overnight digestion was performed with the restriction enzyme MspI (New England Biolabs, Ipswich, MA), which cuts the sequence CCGG and generates sticky ends, enabling every read to start with a CpG. Reactions were cleaned up with AMPure XP magnetic beads (Beckman Coulter), and libraries were prepared using the NEXTflex Bisulfite-Seq Kit (Bioo Scientific Corporation, Austin, TX). The digested DNA was end-repaired, A-tailed, and then ligated with the NEBNext Methylated Adaptor (New England Biolabs). The ligated DNA was size-selected to produce a final library size of 350 bp. Bisulfite conversion was performed with the EZ DNA Methylation-Gold Kit (Zymo Research, Irvine, CA, USA), and libraries were barcoded using the NEBNext Multiplex Oligos (New England Biolabs). The resulting libraries were quantified with the Qubit High Sensitivity dsDNA Assay (Life Technologies, Carlsbad, CA) and the Bioanalyzer High Sensitivity Analysis (Agilent, Santa Clara, CA). Libraries were then pooled and sequenced on the Illumina NextSeq 500 with the high-output, 75-bp cycle protocol (Supplementary Table 1).
Differential methylation analysis
RRBS reads were analyzed for quality with FastQC (v0.11.8) [33], followed by trimming with TrimGalore (v0.6.1) [34] with the “—rrbs” parameter specified. Trimmed reads were aligned to the mouse reference genome from Ensembl, version m38 (mm10), with Bismark (v0.20.0) [35] using default parameters. The coverage files outputted from Bismark were used to obtain CpG methylation rates.
We first used principal component analysis (PCA) to visualize the CpG methylation data and determine how samples compared in terms of CpG methylation profiles. The R package methylKit (v1.12.0) [36] was used for differential methylation analyses. CpG sites with less than 10X coverage and greater than the 99.9th percentile of coverage were removed (Supplementary Table 2). For each pairwise comparison performed, CpGs found in 4 samples per group for each of the comparisons were retained for differential analysis resulting in around 400,000 CpGs included in the differential analysis (Supplementary Table 3). For differentially methylated regional (DMR) analysis, the genome was tiled into 1,000 bp non-overlapping regions, and the CpG methylation rates were averaged over the region.
Statistical analysis
We tested the null hypothesis that mouse genotype (APOE3 versus APOE4) has no effect on cognitive function after exposure to anesthesia and surgery. For behavioral data, repeated measures ANOVAs were used to analyze the effects of treatment for the distance moved and the duration of time spent in the center of the open field as outcome measures using SPSS software (Chicago, IL). Because APOE3 and APOE4 mice were not littermates, and were bred as homozygous mice, we assessed these effects in APOE3 and APOE4 mice separately. Paired t-tests compared the time spent exploring the familiar and novel objects in each experimental group in the object recognition test. A one-way ANOVA assessed the effects of treatment on average baseline motion during acquisition day of fear conditioning. A repeated-measures ANOVA compared average motion during the shocks, freezing during the tones, and freezing during inter-stimulus intervals during the fear conditioning training trial. A one-way ANOVA analyzed freezing behavior in the contextual memory test, and a repeated-measures ANOVA assessed differences in freezing behavior between the baseline and during the tone in the cued fear memory test. Because we expected genotype differences a priori, we assessed treatment effects within the genotypes individually.
For differential methylation analyses, a logistic regression model using a Chi-square test determined if the model with the treatment vector better predicts the outcome variable (methylation proportion) than the null model. Our model included treatment with genotype and sex as covariates. p-values were corrected to q-values via the SLIM method [37]. An over-dispersion correction was applied. Significant DMRs were defined by selecting DMRs with a q-value < 0.1 and percent methylation difference of at least 10%. Ensembl annotations (GRCm38.99) were used to annotate significant DMRs to overlapping gene regions and nearby genes using custom scripts and ChIPseeker (v 1.22.0) (Supplementary Tables 4 and 5) [38]. Since DMRs were annotated at the transcript level and multiple alternative transcripts might be present for a given gene, a single DMR can overlap multiple gene features within the same gene. While we report all instances in the supplementary tables, we prioritized DMRs overlapping promoters for our analyses. Gene IDs were used to perform pathway analyses using EnrichR [39]. All significant pathways (p < 0.01) are reported in Supplementary Table 6.
RESULTS
Behavioral performance in the open field and novel object recognition
First, we assessed the effects of surgery on exploratory behavior in the open field, which involves hippocampal function. During habituation to the open field, total distance moved (cm) was used as a measure of exploratory behavior. There was a significant effect of surgery on activity levels in the E3 but not E4 mice. On days 1 and 2, E3 surgery mice moved significantly less in the open field than E3 sham mice (F(1,21) = 4.755, p = 0.041). In contrast, no effect of surgery was seen in the E4 mice (F(1,29) 0.001, p = 0.981) (Fig. 3A, B).

Results from the open field (A-D) and novel object recognition (E, F). A) During exploration of the open field, E3 mice moved less on day 2 than day 1, F (1, 21) = 87.526, p < 0.01. On both days of the open field, E3 surgery mice moved significantly less than E3 sham mice, F (1, 21) = 4.755, p = 0.041. B) E4 mice also moved significantly less on day 2 than day 1, F (1, 29) = 330.847, p < 0.01. However, there were no significant treatment effects. C) When measuring duration in the center of the open field, E3 mice spent significantly less time in the center of the arena on day 2 than day 1, F (1, 21) = 43.133, p < 0.001. E3 surgery mice spent more time in the center of the open field across both days than E3 sham mice, F (1, 21) = 5.874, p = 0.024. D) While E4 mice spent significantly less time in the center of the open field on day 2 than day 1 (F (1, 29) = 12.891, p = 0.001), there was no significant effect of treatment. E) During the novel object recognition task, the E3 sham group spent significantly more time with the novel object than the familiar object (t (9) = –4.958, p = 0.001) suggesting successful novel object recognition, a trend that was not observed in the E3 surgery group. F) In E4 mice, there were not significant differences in either treatment group in object exploration.
Next, we assessed the effects of surgery on time spent in the more anxiety-provoking center of the open field. E3 surgery mice spent more time in the center on both days than E3 sham mice (F(1, 21) = 5.874, p = 0.024). In contrast, no effect of surgery was seen in the E4 mice (F(1, 29) = 0.010, p = 0.923). Thus, E3 surgery mice have decreased motor activity and display less anxiety-like behavior than E3 sham mice (Fig. 3C, D). We then analyzed whether activity levels in the open field were related to time spent in the center of the open field. In both genotypes, there was a positive correlation between activity levels and time spent in the center of the open field (E3: r = 0.3793, p = 0.0093, Spearman, 46 X-Y pairs; E4: r = 0.4844, p < 0.0001, Spearman, 62 X-Y pairs, Supplementary Figure 1). These data suggest that while activity levels and time spent in the center are related, they cannot explain the genotype-dependent effects of surgery only seen in E3 mice.
E3 sham mice spent significantly more time with the novel than the familiar object (t(9) = 4.958, p = 0.001), while this preference for exploring the novel object was not seen in E3 surgery mice (t(5) = 0.692, p = 0.520). Within the E4 genotype, neither sham nor surgery mice showed a preference for exploring the novel object (sham: t(8) = 1.604, p = 0.147, surgery: t(2) = 0.814, p = 0.501). Thus, E3 surgery mice show impaired novel object recognition, while E4 mice show impaired novel object recognition in both surgery and sham groups (Fig. 3E, F).
Spatial learning and memory in the water maze
There were no significant differences between sham and surgery groups in cumulative distance to the visible platform (cued learning) in either genotype (Fig. 4A, B). After mice were trained to locate the first hidden platform location, there was no effect of surgery on E3. However, E4 surgery mice had a lower cumulative distance to the platform in trial 7 than E4 sham mice, F(1, 29) = 9.736, p = 0.004 (Fig. 4 B). There were no significant differences between treatment groups in either genotype for cumulative distance to the target platform location during the first and second probe trials (no platform). There was no effect of surgery in either E3 or E4 mice’s ability to locate the second hidden platform location (Fig. 4C, D).

Results from water maze (A - D) and fear conditioning (E - H). A) During water maze, in the E3 mice, there were no significant differences between treatment groups in cumulative distance to the platform. B) In the E4 mice, sham mice had a significantly higher cumulative distance to the platform in trial 7 than the surgery mice, F (1, 29) = 9.736, p = 0.004. C, D) There were no significant differences between treatment groups in either genotype for cumulative distance to the target platform in both probe locations during Probe 1 and Probe 2. E) During fear conditioning acquisition, E3 mice froze more during tone 2 than tone 1, demonstrating successful association of the tone with the shock F (1, 21) = 27.483, p < 0.001. Additionally, within the E3 mice, surgery mice froze significantly more overall than sham mice F (1, 21) = 4.876, p = 0.038. F) While E4 mice froze significantly more during tone 2 than tone 1 (F (1, 29) = 41.661, p < 0.001), there were no significant effects of treatment within this genotype. G) During contextual fear memory recall, there was a significant effect of treatment in E3 mice, such that E3 surgery mice froze significantly more than E3 sham mice, F (1, 21) = 5.212, p = 0.033. H) There was no significant effect of treatment in E4 mice.
Contextual and cued fear learning and memory
During fear conditioning, activity levels prior to a training period were assessed. There was no effect of surgery on the baseline activity or response to shock in either genotype.
During the training session, percent freezing was used as a measure of fear learning in mice. There was an effect of surgery on freezing during the conditioned stimulus, the tone, in the E3 mice only. E3 surgery mice froze significantly more than E3 sham mice, F(1, 21) = 4.876, p = 0.038 (Fig. 4E); however, there was no significant effect of surgery in the E4 mice (Fig. 4F).
Hippocampus-dependent contextual fear memory was assessed one day after the training day. There was an effect of surgery in the E3 genotype only; E3 surgery mice froze more than E3 sham mice, F(1, 21) = 5.212, p = 0.033 (Fig. 4G, H).
Finally, hippocampus-independent cued fear memory was assessed by placing the mice in a new environment. After a 90 s baseline, the tone was played that was also present during the training. There was no effect of surgery on freezing in either genotype (Supplementary Table 7).
Exposure to anesthesia and surgery results in changes in DNA methylation
We applied RRBS on genomic DNA extracted from the hippocampi of aging E3 and E4 mice to identify differential DNA methylation between the mice that underwent surgery versus sham and determine if such differences depended on genotype. Using PCA to compare global CpG methylation profiles, there was no separation between the surgery and sham groups in either E3 or E4 mice (Fig. 5A), suggesting the absence of global differences between the different groups. When performing our differential methylation analysis between surgery and sham, we obtained 313 differentially methylated regions (DMRs) for the E3 mice comparison, generally split between hypo- and hyper-methylated (164, 52% and 149, 48%, respectively). There were more DMRs (416) for the E4 mice comparison, similarly split between hypo- (214, 51%) and hyper-methylated (202, 49%). We then annotated the DMRs for overlap with respect to genes (i.e., promoters, introns, exons, 5’ UTR, 3’ UTR, and inter-genic). We observed some variation in the distribution between hyper- and hypomethylated DMRs. For instance, promoters most commonly overlapped with hypermethylated DMRs in both E3 and E4 (41% and 36%, respectively; Fig. 5B and Supplementary Table 8). In turn, hypomethylated DMRs were skewed towards introns and intergenic regions.

A) PCA does not show evident separation between the groups when surgery (surg) and sham are compared within each genotype and when surgery groups are compared between genotypes, suggesting the lack of global changes. B) The bar graph shows the distribution of hyper- and hypomethylated DMRs with respect to genomic features for each comparison.
Overall, we found little overlap between the DMRs identified in the two comparisons. We identified only two overlapping DMRs; one overlaps with the promoter of the Scratch Family Transcriptional Repressor 2 (scrt2) and is hypermethylated in the surgery group in both comparisons. The other region overlaps with a predicted gene (Gm38247).
Pathway analysis of the DMRs identified several biologically relevant pathways in both genotypes, a selection of which are reported in Fig. 6 (see Supplementary Table 6 for complete list). Axonogenesis (GO: 00007409) was the single pathway identified in both E3 and E4 mice which was enriched after surgery in hypomethylated DMRs in both genotypes. Of note, the ephrin receptor signaling pathway was the most significantly (p = 0.0002) enriched in hypermethylated DMRs after surgery in E3 mice. For two of the genes belonging to this pathway (ephrin B2, Efnb2 and Eph receptor A7, Epha7), the DMRs identified correspond to regions that have been annotated as accessible (i.e., active) chromatin through Assay for Transposase-Accessible Chromatin with high-throughput sequencing (ATAC seq) and active promoter chromatin state in the mouse forebrain by the ENCODE project (Fig. 7) [40]. This supports a functional role for these DMRs in the mouse brain.

A selection of the most significant GO biological processes identified using EnrichR Pathway Analysis are shown for hyper- and hypomethylated DRMs in each comparison (full list available in Supplementary Table 6). Pathways are ranked based on the -Log10 of p-values.

Screenshots from the UCSC Browser show three genes belonging to ephrin receptor signaling pathway. DMRs are indicated by the black blocks and highlighted by red boxes. For the two top genes, an overlap is evident between the DMR, CpG island, and the active promoter chromatin state in the mouse forebrain (in green). ATAC-seq peaks delineate accessible chromatin within the same regions.
DISCUSSION
Postoperative NCDs in geriatric patients represent an important clinical problem that is not well understood at the molecular level. Therefore, we developed a murine model that combines four relevant factors related to postoperative NCD: surgery, general anesthesia, aging, and genotype. We used the model to test the hypothesis that exposure to surgery and anesthesia alters DNA methylation in a brain region involved in cognition. Our findings suggest that the neurocognitive and behavioral effects of surgery and anesthesia differentially affect aging mice depending on the APOE isoform. Furthermore, exposure to surgery and general anesthesia in aging mice results in differential methylation of specific genes within the hippocampus involved in learning and memory.
In aged mice, we found that exposure to surgery and anesthesia reduced motor activity and exploratory and anxiety-like behavior, impaired learning and memory, and increased fear conditioning in aged E3 mice only. The lack of effect of surgery on E4 mice might be related to a “floor effect” in the E4 mice due to a different cognitive baseline compared to E3 mice. This is consistent with our previous publications in young E4 mice [41, 42] and is likely further exacerbated in aged mice in the current study. In clinical studies, evidence on whether E4 is a risk factor for postoperative NCD is conflicting [19 , 43]. Although evidence suggests that APOE4 is associated with long-term cognitive decline after surgery, it remains unclear whether APOE4 is associated with postoperative delirium or NCD within the first year after surgery [44]. A meta-analysis concluded that the APOE4 allele was associated with a significantly increased postoperative cognitive dysfunction, but the association depended on one large study, and no association was found 1–3 months and 1 year after surgery [21]. Our previous studies demonstrated that older surgical patients with an APOE4 allele had a more rapid rate of cognitive decline in the years following surgery and general anesthesia than those without an APOE4 allele [19, 23]. It is possible that clinical studies may in part be confounded by a different cognitive baseline in the APOE4 genotype in the aging human population. In the Oxford Project to Investigate Memory and Ageing (OPTIMA) cohort, surgery accelerated cognitive decline in older patients with pre-existing cognitive impairment, but not other geriatric patients, and there was no independent influence of the APOE4 allele [45]. Although pre-existing cognitive impairment is known to be one of the major risk factors for postoperative NCD [46], there have been few studies exploring the effect of surgery and anesthesia on older patients with mild cognitive impairment (MCI) or AD. One preliminary retrospective study found that patients with MCI that underwent surgery had a greater decline in tests of attention and concentration than individuals with MCI that did not have surgery [47]. A prospective, randomized parallel-group study found that inhaled sevoflurane accelerates the progression of MCI to progressive MCI in a selected Chinese population [48].
It has been hypothesized that epigenetic mechanisms may influence an individual’s reaction to surgery and anesthesia [49]. In fact, changes in hippocampal DNA methylation have been reported to be involved in sevoflurane-induced cognitive impairment in neonatal rats [50]. Another study demonstrated that rats subjected to maternal separation during the neonatal period exhibit behavioral changes and neuroinflammation after sevoflurane anesthesia linked to DNA methylation of the glucocorticoid receptor [51]. While those studies shed light on the effects of anesthesia and stress on the developing brain, the role of DNA methylation in mediating the effects of perioperative stress in the aged brain is less clear.
To investigate potential molecular mechanisms underlying the observed behavioral changes, we compared genome-wide DNA methylation between surgery versus sham mice. There were no differences in global DNA methylation between surgery and sham groups in either E3 or E4 mice, suggesting that epigenetic changes might affect specific genes/pathways. To determine if behavioral changes correlate with DNA methylation changes of specific genes, we identified DMRs between surgery versus sham mice. We identified 313 DMRs for the E3 comparison and 416 for the E4 comparison. These DMRs were equally split between hypo- and hyper-methylated regions in both genotypes. Interestingly, after annotating the DMRs for overlap with genes, we found that promoters mostly overlapped with hypermethylated DMRs in both E3 and E4, while hypomethylated DMRs were skewed towards introns and intergenic regions. This observation suggests that the stress of surgery/anesthesia might have a repressive effect on gene expression, as both promoter hypermethylation and gene body hypomethylation are inversely correlated with gene expression [52].
Because E3 mice are the only group that showed a clear change in behavioral and cognitive performance following surgery and anesthesia, differentially methylated genes in this group are of particular interest. Using pathway analysis, DMRs were significantly enriched in the Ephrin receptor signaling pathway in E3 mice after surgery. Ephrin receptors are the largest known subfamily of receptor tyrosine kinases. Ephrins and their receptors are membrane-bound proteins that require direct cell-cell interactions for Eph receptor activation. The Eph/ephrin system regulates many developmental processes, such as axonal guidance and adult tissue functions, including long-term potentiation [53]. Our analysis revealed that the DMRs overlapping Ephrin B2 and Eph A7 receptor genes also correspond to accessible chromatin, suggesting a possible functional role for these DMRs in mediating the effects of surgery/anesthesia on cognition and behavior.
Eph receptors and corresponding ephrin ligands contribute to vital neuronal functions, including memory formation, by regulating events such as presynaptic transmitter release, postsynaptic glutamate receptor conductance and trafficking, synaptic glutamate reuptake, and dendritic spine morphogenesis [54]. These proteins participate in forming various types of memories that are subserved by different neurons and brain regions. Importantly, Eph receptors and ephrins play essential roles in cognitive impairment and behavioral changes characteristic of several brain disorders, including AD [55]. For example, Eph B2 depletion contributes to amyloid-beta-induced neuronal deficits and cognitive dysfunction by impairing NMDA-type glutamate receptor functions [56]. Depleted Eph B2 levels have been detected in mouse models of AD and in humans with AD. Increasing Eph B2 levels in a mouse model of AD improves memory [56]. Eph A receptors and ligands are expressed primarily in the hippocampus and are implicated in hippocampal synaptic plasticity and learning. Relevant to the current study, anesthesia leads to retrograde amnesia in mice, and post-anesthesia activation of Eph A receptors significantly ameliorates this amnesia [57]. Eph receptors and Ephrin ligands may therefore induce cellular alterations mandatory for memory formation and serve as a target for pharmacological intervention for the treatment of memory-related brain diseases, including postoperative NCDs.
In addition to genes specifically modified after surgery in E3 mice, some differences in methylation were observed in both E3 and E4, but these epigenetic modifications did not correlate with a change in behavioral performance after surgery in E4 mice. Therefore, these genes are either implicated in functions other than cognition, their impact may not have been apparent within the experimental conditions or timeframe of our study, or they could simply be biomarkers for anesthesia/surgery exposure. For example, we found that scrt2 promoter is hypermethylated after surgery in both E3 and E4 groups. Scrt2 expression is highly enriched in the brain [58] and has been implicated in neurogenesis and neuronal migration, and survival during embryonic brain development [59 –61]. Since scrt2 is a transcriptional repressor, its hypermethylation potentially indicates activation of downstream genes. These genes’ identity and function are unknown, but they could be related to the effect of anesthesia on neurogenesis. For example, unlike its effect in the developing brain, where isoflurane decreases neurogenesis, scrt2 has been shown to enhance neurogenesis in adult animals [62]. Therefore, it is possible that scrt2 hypomethylation is indicative of an effect on neurogenesis, of which the functional consequence on neurocognitive and behavioral outcomes may be delayed beyond the 4-week testing period in our study, or masked by adverse effects of aging and E4 on neurogenesis. Interestingly, in a recent study of DNA methylation changes in the blood of geriatric patients after major surgeries, scrt2 was identified as being significantly hypomethylated [63]. The latter observation is the opposite of our finding that scrt2 is hypermethylated in the hippocampus, suggesting that scrt2 is activated in blood, which leads to downstream gene suppression in blood. This again emphasizes tissue specificity of epigenetic DNA modifications and suggests that while methylation changes in blood could be used as biomarkers for anesthesia exposure, they may not necessarily reflect similar directional changes in the brain.
Our study has the following limitations. The first limitation is that we did not monitor blood pressure and oxygen saturation, as recommended by guidelines for rodent postoperative NCD research published following the completion of our experiments [64]. Although we monitored temperature, respiration rate, and mucous membrane color, we cannot completely rule out hypotension and hypoxia as confounders responsible for group differences. Next, we recognize that residual pain might have contributed to the reduced activity levels and enhanced contextual fear memory of E3 surgery mice. Another important limitation to consider is that our study was not designed to distinguish between the effects of anesthesia versus surgery. Like most rodent studies of perioperative neurocognitive disorders [64 –70], we attempted to make our model more representative of patient populations with postoperative neurocognitive disorders by using aged animals that are subjected to both surgery and general anesthesia rather than anesthesia alone. Prior studies have found that anesthetics have few behavioral or neuroinflammatory effects on their own, but they can modulate the effects of surgery or genetics [69, 70]. Also, for reasons of animal welfare, IACUC-guidelines, and translational relevance, buprenorphine was included as part of the postoperative care plan. Because we did not treat control mice with buprenorphine, we cannot exclude potential effects of buprenorphine on behavioral performance and effects on brain DNA methylation observed in the surgery groups. Review of the literature and a closer look at our own data suggest that it is unlikely that buprenorphine mediated the effects of surgery on behavior or cognition. In addition to its analgesic effect, buprenorphine typically causes moderate excitement and agitation, which is associated with increased motor activity [71], rather than decreased motor activity seen in E3 mice in our study. Similarly, a reduction in pain is associated with a reduction in anxiety, rather than the increased anxiety-like behavior seen in E3 mice in our study. Furthermore, although E4 surgery mice were also treated with buprenorphine, there were no differences in motor activity or anxiety-like behavior in these mice compared to genotype-matched control animals, nor were there differences in distance travelled in the water maze between buprenorphine-treated and untreated control mice. Similar inconsistencies and lack of association between surgery + buprenorphine treatment and results can also be seen in cognitive testing.
Finally, although mice were treated with buprenorphine for 3 days after surgery, buprenorphine treatment was stopped 24–48 h before behavioral testing. Specifically, open-field testing was performed on postoperative days 4 or 5, while the remaining tests were conducted more than a week after buprenorphine treatment had stopped, and hippocampal tissues were dissected for DNA extraction 4 weeks after the treatment had stopped. This is important because buprenorphine’s half-life in rodents is only 3 hours [72, 73], and its analgesic effect lasts no more than 12 hours, which is why it is administered twice daily [74, 75]. Therefore, by 24 h, buprenorphine is mostly cleared from the circulation, and the remaining levels are below the effective concentrations. By the second day of testing (48 h), buprenorphine should be completely cleared from the circulation.
As discussed above, studies have shown some differential DNA methylation following exposure to inhalational anesthetics on the developing brain, but few studies have explored whether inhalational anesthetics alone, without surgery, lead to epigenetic changes in the aging brain. An in vitro recent investigation found that isoflurane exposure to primary hippocampal neurons did not alter methylation of genes in an apoptosis or cytokine array, but did have methylation changes for several inflammatory genes. On the other hand, when cells from a permanent hippocampal cell line were exposed to isoflurane, there were no changes in methylation in genes in the apoptosis, inflammatory, nor cytokine arrays [76]. Although there is literature regarding epigenetic brain changes related to chronic opioid addiction, evidence is scare regarding epigenetic changes after short term use of buprenorphine or other opioids. One group measured global 5-hydroxymethylcystosine (5 hmC) levels in 10 different brain regions, including the hippocampus, following acute and chronic morphine exposure in rats. 5 hmC is an intermediate in active DNA methylation [77]. When compared to control rats, significant changes in 5 hmC levels were identified after chronic morphine exposure, but there were no significant changes following acute morphine administration. They also found no alterations in DNA methylation of mitochondrially-encoded genes after morphine exposure. After acute morphine exposure, the group did identify differential methylation of genes previously identified as implicated in morphine tolerance, including Il6, Nr3c1, Bdnf, and Il1b [77]. More studies are needed to determine the effects of acute opioid administration on DNA methylation and other epigenetic changes in the brain. Although our study is not designed to determine the effects of anesthesia or analgesia on DNA methylation, we believe that we have a more clinically relevant representation of the perioperative period.
In summary, we developed a murine model that combines several relevant factors related to postoperative NCD: surgery, general anesthesia, aging, and genotype. We found that baseline neurocognitive and behavioral performance is affected by E4 and that E3 mice sustain neurocognitive and behavioral alterations after surgery/anesthesia, which correlate with gene-specific changes in DNA methylation. Pathways identified in this study may serve as potential therapeutic targets to prevent and treat postoperative NCD and could be used as markers and predictors of long-term functional outcome in geriatric patients undergoing surgery under general anesthesia.
Footnotes
ACKNOWLEDGMENTS
The authors would like to thank Marie Kane for her assistance in editing and formatting. We thank Davelle Cocking for performing the surgical procedures. We thank Tessa Marzulla and Lance Johnson for their assistance in initiating this study. We thank Sydney Boutros and Davelle Cocking for their assistance in planning this study out in detail and for their assistance with the mice’s postoperative care. We thank Eileen Ruth Samson Torres for her help in analyzing the behavioral data.
Dr. Schenning acknowledges this work was supported by K12 HD 043488, R03 AG056462-02, the Foundation for Anesthesia Education and Research (FAER), and an Alzheimer’s Association Clinician Scientist Fellowship. This work was partially funded by RF1 AG059088, RF1 AG058273, R01 NS077767, and R21 ES027943.