
Abstract
Background:
The associations between sleep characteristics and cognition are complicated. Alzheimer’s disease (AD) pathologies have been proven to be associated with sleep characteristics.
Objective:
We aimed to investigate the associations between sleep characteristics and cognitive function and examine the roles of AD pathologies in modulating the association of sleep duration with cognition.
Methods:
A total of 974 participants who had measurements of cerebrospinal fluid (CSF) amyloid-β (Aβ), phosphorylated tau (P-tau), total tau proteins (T-tau), cognitive function, and sleep characteristics were included from the Chinese Alzheimer’s Biomarker and Lifestyle (CABLE) study. Linear regression analyses were utilized to explore the associations of sleep characteristics with cognition. Non-linear regression analyses were utilized to explore the associations of sleep habits with cognition. Causal mediation analyses were conducted to explore the mediation effects of AD pathologies on cognition.
Results:
The Pittsburgh Sleep Quality Index (PSQI) total score was significantly negatively correlated with Montreal Cognitive Assessment (MoCA) score (p = 0.0176). Long latency (p = 0.0054) and low efficiency (p = 0.0273) were associated with cognitive impairment. Habitual nap behavior was associated with lower MoCA scores (p = 0.0045). U-shaped associations were observed between sleep habits (bedtime and nocturnal sleep duration) and cognition. A causal mediation analysis indicated that P-tau/Aβ42 mediated the association of sleep duration with cognition.
Conclusion:
These findings showed sleep characteristics were associated with cognitive functions. Sleep habits (duration, bedtime) had U-shaped associations with cognition. AD core pathologies might partially mediate the influence of sleep duration on cognitive impairments.
INTRODUCTION
Some of the changes in sleep patterns that can occur in older adults include insufficient sleep time, decreased sleep efficiency, decreased quantity of slow wave and rapid eye movement (REM) sleep, increased sleep fragmentation, and increased awakenings after sleep onset [1–5]. Two cohort studies on self-reported sleep characteristics increased the risk of dementia [6, 7]. Sleep is considered as a modifiable risk factor for dementia and cognitive impairment [8].
Several studies have documented a significant association between sleep and cognitive function in the elderly [9–14]. Results were inconsistent on whether baseline sleep characteristics were associated with cognitive decline and incident dementia. Some studies reported that sleep characteristics (quality, duration, and disturbances) were statistically significantly associated with cognitive impairment and incident dementia, while others observed no association [6, 14]. However, there is still a need for long term prospective studies to confirm the associations between sleep habits and cognitive decline. Preserving and protecting sleep health may be critical to helping maintain cognitive health.
In recent years, a number of studies have reported the associations between sleep characteristics and Alzheimer’s disease (AD) pathologies. Several cross-sectional studies suggested that insufficient sleep duration [15, 16] (including sleep deprivation [17, 18]), obstructive sleep apnea (OSA) [19–21], poor sleep quality [15, 23], excessive daytime sleepiness (EDS) [16, 23], long latency [24, 25], low efficiency [24, 26], and fragmentation [24] were associated with greater amyloid deposition and higher risks of all-cause dementia or cognitive impairments [27]. We and other teams suggested that sleep duration had U-shaped relationships with biomarkers of AD core pathologies [27–30].
In the present study, we aimed to examine 1) the self-reported sleep characteristics associated with cognitive function and 2) the roles of AD pathologies in modulating the association of sleep duration with cognition.
METHODS
Study population
A total of 974 adults without dementia were included from the Chinese Alzheimer’s Biomarker and Lifestyle (CABLE) study. The CABLE study is an ongoing large-scale cohort study initiated in 2017, which mainly focuses on AD risk factors and biomarkers in the northern Chinese Han population. All participants were Han Chinese in origin and were aged between 40 to 90 years. Exclusion criteria comprised 1) major psychological disorders (e.g., major depressive disorder or generalized anxiety disorder, etc.); 2) major neurological disorders (e.g., trauma, infection, epilepsy, or multiple sclerosis, etc.); 3) malignant tumors; and 4) genetic disorders (e.g., early-onset familial AD, genetic ataxias, and hereditary spastic paraplegia, etc.). All participants underwent clinical and neuropsychological assessments via a structured questionnaire and an electronic health record system. In addition, biological samples (blood and CSF samples) were collected by doctors with standardized training.
The study was approved by the Institutional Ethics Committee of Qingdao Municipal Hospital and written informed consent was obtained from all participants or their caregivers according to the Declaration of Helsinki.
Sleep characteristics
Sleep time and sleep quality were measured using the Pittsburgh Sleep Quality Index (PSQI) questionnaire. PSQI is a self-reported questionnaire evaluating sleep characteristics during the last 4 weeks, including sleep quality, sleep latency (minutes spent before falling asleep), nocturnal sleep duration (hours), sleep efficiency (the ratio of sleep duration to the total time spent in bed [TIB]), sleep disturbances, sleep drug use, and daytime dysfunction. The sum of the scores of the above-mentioned seven components was an overall score ranging from 0 to 21 (the global PSQI score), with higher scores indicating worse sleep quality. In addition, several specific questions (e.g., “when do you go to bed?”, “Do you snore when you sleep?”, “Do you have habitual daytime napping?”, “How long does each napping last?”) were also included in the questionnaire. All participants completed the PSQI questionnaire the day before the lumbar puncture.
Cognitive assessments
General cognitive function was evaluated by Montreal Cognitive Assessment (MoCA). The maximum score of MoCA is 30, with higher scores indicating better cognitive function. MoCA tests multiple cognitive subdomains including executive function, fluency, orientation, calculation, abstraction, delayed recall, visuospatial, naming, and attention. Specially trained neurologists administered the tests [31].
Previous studies showed the MoCA was a more sensitive and accurate tool in screening for MCI compared to the Mini-Mental State Examination (MMSE), suggesting that the MoCA was superior to the MMSE in discerning earlier stages of cognitive decline [31, 32].
CSF AD biomarkers
In CABLE, CSF was taken by lumbar puncture and processed within 2 h. CSF samples were centrifuged at 2000×g for 10 min. CSF was separated and stored in an enzyme free EP tube at –80°C until assay. Avoid repeated freeze-thaw cycles (less than 3 times). CSF Aβ42, Aβ40, total tau (t-tau), and phosphorylated tau181 (p-tau181) were detected by the enzyme-linked immunosorbent assay (ELISA) kit (Innotest; Fujirebio, Ghent, Belgium) on the microplate reader (Multiskan MK3; Thermo Fisher Scientific, Waltham, MA). All CSF samples were measured in duplicate by trained technicians who were blinded to clinical information. The within-batch precision values were < 6%(5.6%for Aβ42, 3.8%for Aβ40, 2.5%for p-tau, and 4.8%for t-tau). The inter-batch CVs were < 11%(7.7%for Aβ42, 10.6%for Aβ40, 10.1%for p-tau, and 7.4%for t-tau).
APOE ɛ4 genotyping
DNA was extracted from fasting blood samples with QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany). Fasting blood specimens were centrifuged at 2000×g for 10 min after being collected and stored at –20°C until assay. The extracted DNA was isolated and stored in an enzyme-free EP tube at –80°C for apolipoprotein E gene (APOE) ɛ4 genotyping. Avoid repeated freeze-thaw cycles (less than 3 times). Specific loci (rs7412 and rs429358) were selected for genotyping with restriction fragment length polymorphism (RFLP) technology.
Statistical analyses
Characteristics of the participants were summarized using descriptive statistical methods. As for MoCA scores, transformation was performed to achieve or approximate a normal distribution (Kol-mogorov-Smirnov test > 0.01) via “car” package of R software in case of skewed distribution [33].
Multiple linear regression models were run separately for MoCA scores and sleep measures (seven domains in PSQI), with the MoCA as the dependent variable and sleep characteristics as the independent variables. Each linear model was adjusted for age, gender, years of education, APOE ɛ4 status, medical history (hypertension, diabetes, coronary heart disease, and stroke), smoking and alcohol intake. Apart from the above factors, sleep quality and sleep duration were adjusted in the multiple linear regression models of nap habits and MoCA scores.
To investigate the nonlinear associations between sleep habits (nocturnal sleep duration, or the bedtime) and MoCA scores, non-linear regression analyses via the quadratic model (y =βx2 +αx + c) were employed [34]. When the coefficient of the quadratic term was significantly larger (U-shaped) or smaller (reverse U-shaped) than zero, significant nonlinear associations were implied. Covariates include age, gender, years of education, APOE ɛ4 status, medical history (hypertension, diabetes, coronary heart disease, and stroke), smoking and alcohol intake.
To test whether there was a mediation effect (sleep duration ⟶ AD pathologies ⟶ cognition) on the development of cognition, we used a general approach to the estimation of indirect effects in simple mediation models and made inferences about instantaneous indirect effects θ. θ means the instantaneous indirect effect of X on Y through M. It quantifies the change in Y through M as X is changing. The analysis is a formal approach to exploring the mechanism in which AD pathology may work as a mediator between sleep duration and cognition [34].
Bonferroni method was used for multiple correction (for seven sleep characteristics) except where specifically noted. R version 3.5.1 (R Project for Statistical Computing; http://www.r-project.org) and GraphPad Prism 8.00 software were used for statistical analyses and figure preparation.
RESULTS
Participant characteristics
Participant characteristics in CABLE were summarized in Table 1. A total of 974 individuals who underwent the MoCA and completed the PSQI questionnaire were included. They were cognitively unimpaired (mean CM-MMSE score = 27.5) individuals in the late midlife with a mean age of 61.6 years old (standardized deviation [SD]: 10.3 years old) and a female proportion of roughly 42.1%.
Basic characteristics of participants included
MOCA, Montreal Cognitive Assessment-Basic; CSF, cerebros-pinal fluid; AD, Alzheimer’s disease; PSQI, Pittsburgh Sleep Quality Index.
PSQI domains and cognitive function
Three sleep characteristics (latency, efficiency, sleep duration) showed significant associations with some cognitive function domains, especially orientation and abstraction function (Fig. 1). A significantly negative correlation between PSQI total score and MoCA score (Fig. 2A) (β= –6.64 p = 0.0176) was observed. Sleep latency (Fig. 2B) (β= –29.66 p = 0.0054) and sleep efficiency (Fig. 2C) (β= –19.44 p = 0.0273) were associated with lower MoCA scores.

Heatmap showing the associations of the PSQI with cognitive domains. PSQI showed several suggestive or significant associations with different cognitive domains in the whole participants. There is a difference in the function of delayed recall between the elderly with and without the habit of taking a nap.
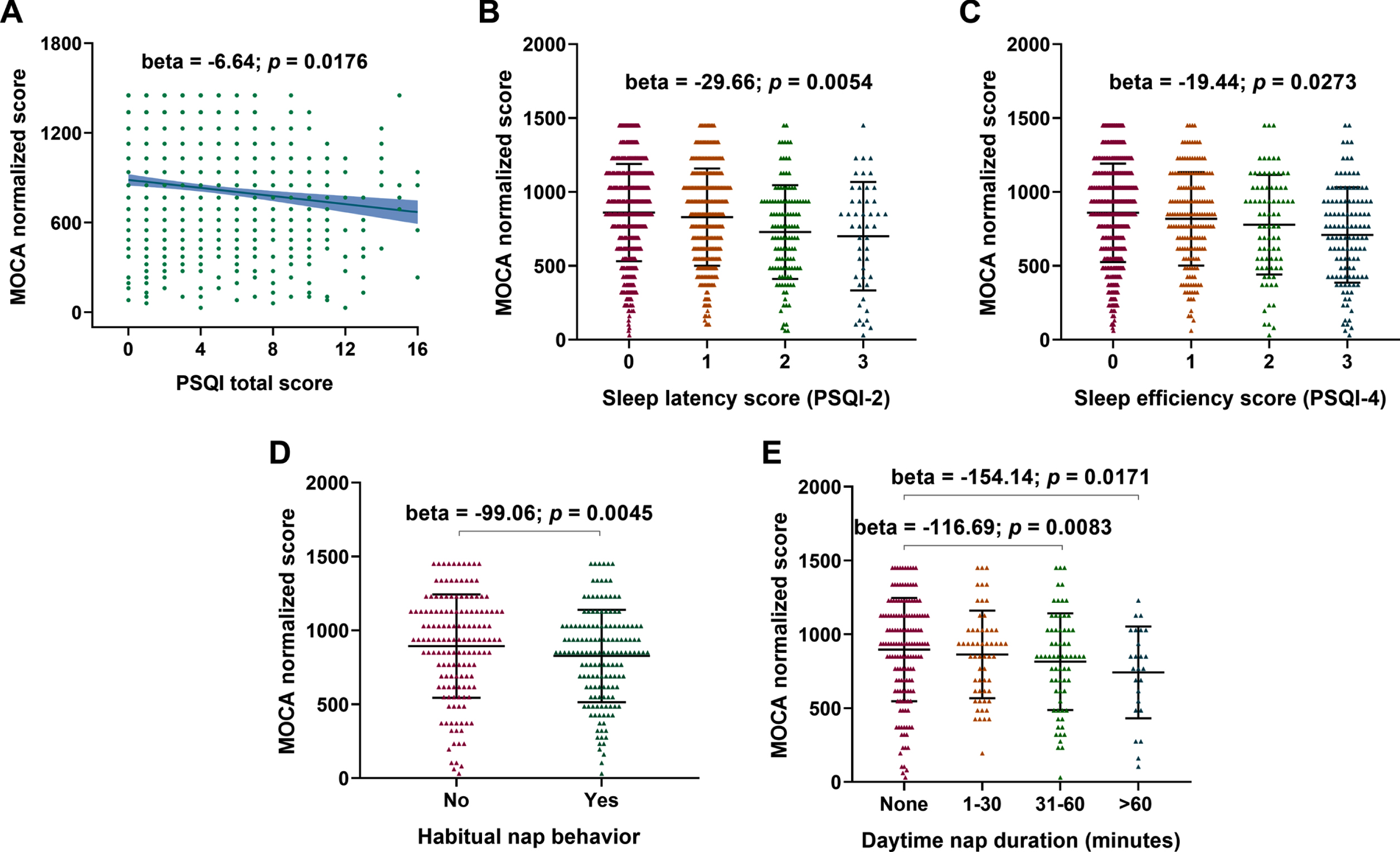
Linear relationships between MoCA scores with PSQI total scores and different sleep characteristics. A negative correlation was revealed between of MoCA score with PSQI scores (A) and nap habits, especially sleep latency (B) and sleep efficiency (C) scores.
Nap duration and cognitive function
The elderly with habitual daytime napping had low MoCA scores (Fig. 2D) (β= –99.06 p = 0.0045). And as the nap duration increased, the MoCA scores decreased gradually. People with 31–60 minutes nap duration (Fig. 2E) (β= –116.69 p = 0.0083) or excessive (>60 min) nap duration (Fig. 2E) (β= –154.14 p = 0.0171) had significantly lower MoCA scores compared to the no habitual daytime napping group.
Sleep time and cognitive function
We found a significant nonlinear relationship of MoCA scores with bedtime. Non-linear regression analyses showed that the reflection point for bedtime was around 11 : 16 pm (Table 2; Fig. 3A) (β= 247.51). Moreover, a nonlinear relationship was revealed between sleep duration and cognitive function. Both insufficient and excessive sleep durations were associated with lower cognitive function as indicated by lower MoCA scores. Non-linear regression analyses indicated that the reflection point was ≈6.74 h (Table 2; Fig. 3B) (β= 132.79).
Multivariable analyses of associations of global cognitive function and CSF biomarkers with sleep habits
Non-linear regression analyses via the quadratic model (y =βx2 +αx + c) were employed. β and α indicate estimated coefficients of global cognitive function and quadratic term in the model, respectively. Covariates include age, gender, education, APOE ɛ4 status, hypertension (yes versus no), diabetes (yes versus no), coronary heart disease (yes versus no), stroke (yes versus no), smoking (yes versus no) and drinking (yes versus no). ns, non-significant. ap < 0.1 (trend); bp < 0.05 (significant); cp < 0.01 (significant).

Nonlinear relationships of sleep habits with MoCA scores and CSF biomarkers of AD pathologies. Nonlinear relationships were revealed of CSF indicators of AD pathology and MoCA scores with nocturnal sleep habits, including sleep duration (B to D) and bedtime (A).
Sleep time and CSF AD biomarkers
Sleep time was non-linearly associated with CSF AD biomarkers. Insufficient or excessive sleep duration was associated with lower CSF Aβ42 and higher CSF P-tau/Aβ42. The reflection points for CSF Aβ42 and CSF P-tau/Aβ42 were situated around 6 : 00 hours (Table 2; Fig. 3C, 3D) (βAβ42 = 0.00271, βP - tau/Aβ42 = 0.00308).
Causal mediation analyses
The equation θ= 11.11–1.87x (Fig. 4) for the instantaneous indirect effect revealed that the indirect effect of sleep duration through MoCA scores decreased linearly as duration increased. In the elderly with relatively poor duration, increased sleep duration would slightly increase MoCA scores by decreasing CSF P-tau/Aβ. However, in the elderly with sufficient or long sleep duration, an increase in sleep duration would lead to a reduction in MoCA scores by increasing CSF P-tau/Aβ.
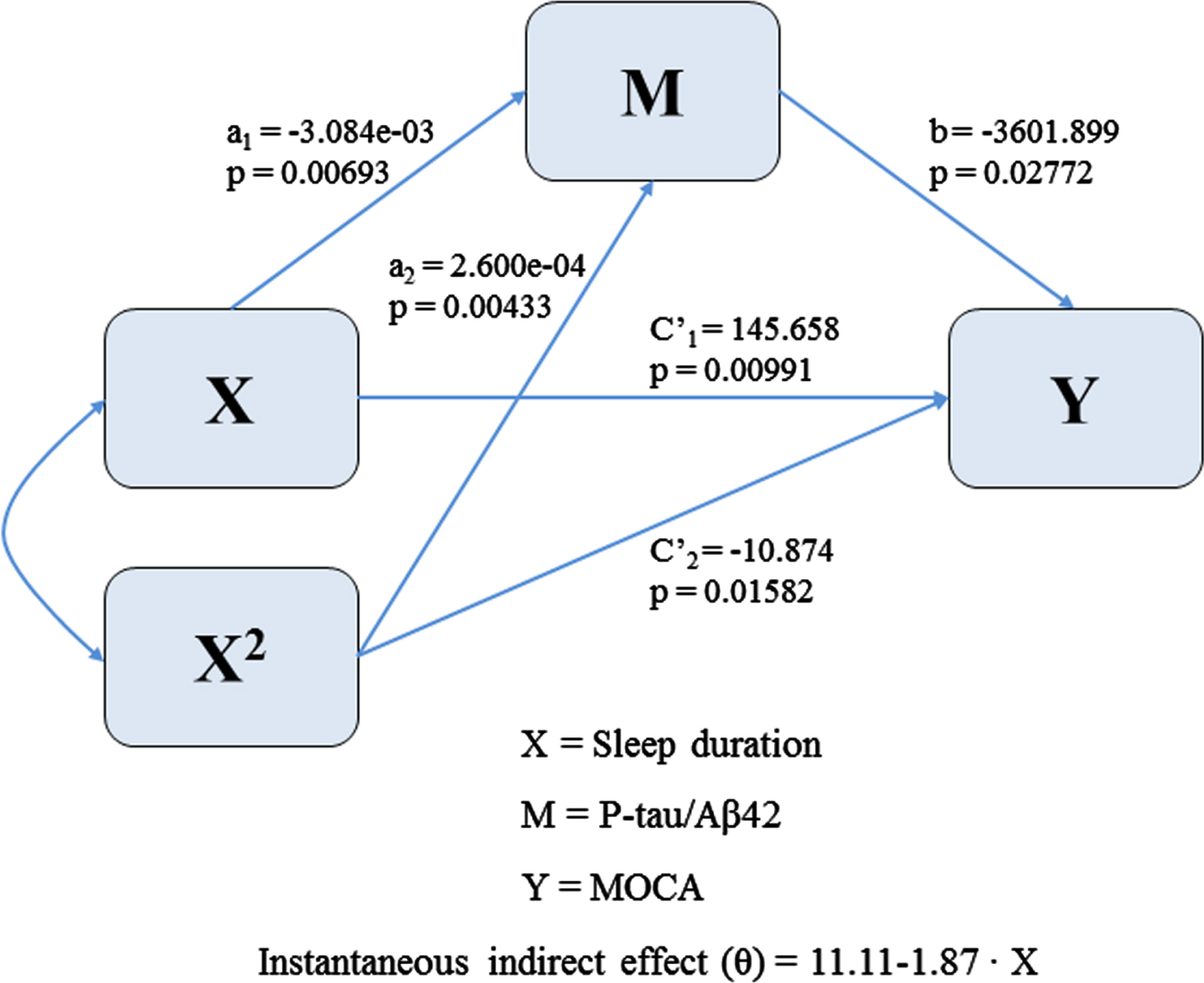
Path model for estimating the instantaneous indirect effect of sleep duration on MoCA scores through P-tau/Aβ42.
DISCUSSION
The present study suggested that several sleep characteristics, including sleep duration, sleep la-tency, sleep efficiency, and nap duration were associated with cognitive function, especially orientation and abstraction. U-shaped relationships between sleep habits and cognitive decline were observed. P-tau/Aβ42 mediated the effects of sleep duration on cognition.
The associations between cognitive decline and sleep characteristics have been investigated. Olivier et al. found scores for the PSQI items, especially sleep disturbances and sleep duration, were significantly associated with 1-year incident cognitive decline [35]. Terri et al. revealed the associations of objectively and subjectively measured sleep characteristics with cognitive impairment. The associations of reduced sleep efficiency, greater nighttime wakefulness, greater number of long wake episodes, and poor sleep quality with cognitive impairment were documented [9]. Recently, Simon et al. reported that shorter sleep latency and longer daytime sleep duration were associated with significantly higher levels of cognitive abilities, such as visuospatial ability, processing speed and verbal memory [11]. Of note, previous studies in samples with wider age ranges found that longer nap duration was associated with cognitive decline, which was consistent with the present study. Simon et al. revealed the associations of nap duration with longitudinal declines in processing speed and visuospatial ability among the elderly [11]. Moreover, the cross-sectional and longitudinal associations of sleep duration and global cognition have been investigated in numerous studies. A large number of clinical trials have shown that sleep deprivation can result in cognitive impairments, including decreased attention, impaired working memory, and perceptual retardation, affecting the normal learning process [36, 37]. Previous studies found that excessive sleep duration was cross-sectionally and longitudinally associated with cognitive decline in the elderly [10, 38]. A U-shaped relationship between sleep duration and cognitive decline was observed. Virta et al. noted that individuals with excessive or insufficient sleep duration had lower cognitive scores [39]. Recently, our team and Ma et al. have reported an inverted U-shaped association between sleep duration and global cognitive impairment [28, 29], which is consistent with some previous studies [13, 40]. However, some studies do not support this finding possibly due to short follow-ups and limited sample sizes [6, 41]. A meta-analysis including 51 eligible cohorts with 15 sleep problems showed that six (insomnia, fragmentation, daytime dysfunction, prolonged latency, sleep behavioral disorder and excessive time in bed) with moderate-to high levels of evidence and four (apnea, no habitual napping, inefficiency, and increased sleep duration) with low levels of evidence, were associated with cognitive decline in the elderly without dementia. The quantitative meta-analysis indicated that either insufficient (<4 h) or excessive (>10 h) sleep duration was a risk factor for all-cause cognitive disorders or AD dementia [27].
Although the associations between sleep characteristics and cognitive decline were revealed, the underlying mechanisms remain unknown. There are many potential mechanisms that could explain these associations. During sleep, the glymphatic system was highly active [42], which contributed to the clearance of metabolic wastes in the brain, such as lactate and Aβ [42–44]. Longer sleep latency, insufficient sleep duration, or other sleep habit changes caused Aβ deposition and tau hyperphosphorylation [18, 45–49]. Besides, sleep characteristic changes might increase Aβ and tau exocytosis [50]. In consistent with the above findings, several lines of evidence showed that sleep disorders increased pathological Aβ deposition in the brain and CSF biomarkers of AD pathology. A review suggested that in addition to the effects of sleep disturbances on Aβ and tau, sleep deprivation induced the accumulation of pathological stress granule aggregates [50]. Another review showed that inflammation mediated the associations between sleep disorders and cognitive decline. Sleep disorders induced increased systemic inflammation, which increased Aβ burden and drove AD pathogenesis [51, 52]. The inflammatory response of the nervous system was mediated by activated microglia. A study found that sleep disturbances or sleep insufficiency induced microglial activation, leading to cognitive decline [53]. Patel et al. observed linear associations of interleukin 6 and CRP with sleep duration, suggesting that excessive sleep duration could induce the activation of interleukin 6 and C-reactive protein (CRP) inflammatory pathways [54]. In this study, the causal mediation analysis suggested that AD pathologies may play non-linear intermediary roles in sleep duration and cognitive ability. In the elderly with relatively poor duration, increased sleep duration would improve cognitive function by decreasing Aβ deposition or/and tau hyperphosphorylation [46, 47]. However, in the elderly with sufficient or long sleep duration, an increase in sleep duration would impair cognitive function by increasing CSF P-tau/Aβ, which is consistent with several studies of the association between long sleep duration and cognitive function [55–57].
Of note, pathological changes might result in changes in sleep habits, which occurred before cognitive impairment. A cohort study found that sleep was significantly impaired in patients with AD, and even the onset of sleep disorders preceded functional and pathological changes [58]. The underlying mechanism might be that brain injury and neurofibrillary degeneration in the brain stem disrupted the maintenance of normal sleep by cholinergic activity [59, 60]. The loss and degeneration of wake-active neurons (orexinergic, dopaminergic, cholinergic, histaminergic, noradrenergic, and serotonergic types) induced by age-related changes in brain integrity resulted in the decline in sleep quality [61]. Also, increased Aβ deposition in the brain was associated with decreased sleep quality via impairing slow-wave sleep [62]. Several studies documented that inflammation mediated the associations between AD pathologies and sleep characteristics [63, 64]. In the elderly AD patients, neuroinflammation prolonged the sleep duration for the immune system to recover [65], resulting in longer sleep and nap durations. Individuals who were sleepier would be the most cognitively impaired. Indeed, healthy older people, who felt sleepy during the day, performed poorly on tests of cognitive abilities [66], and were twice as likely to be diagnosed with dementia three years later [67]. Moreover, demented patients with higher sleep propensity (those with mild to moderate AD) performed worse on tests of cognitive abilities [68]. These findings suggested that changes in sleep characteristics, cognitive degradation, as well as excessive sleep and nap durations might be related to AD pathologies.
This study has several limitations. 1) It was a cross-section study, so no causal relationships could be demonstrated. Reverse causality is possible. 2) Sleep characteristics was based on self-reported information rather than objective or subjective measurement, which might result in random bias. 3) PSQI was used to evaluate the short-term but not long-term sleep situation. Longitudinal studies with larger sample sizes are also warranted to explore the associations between sleep habits and cognition.
In summary, sleep habits were associated with cognitive function. AD pathologies may mediate the effects of sleep duration on cognitive function. In general, sleep measurement may be used to identify individuals at risk of cognitive decline. Adopting healthy sleep habits (adequate sleep quality and duration) may benefit cognitive function.
Footnotes
ACKNOWLEDGMENTS
The authors thank all participants of the present study as well as all members of staff of the CABLE study for their role in data collection.
This study was supported by grants from the National Natural Science Foundation of China (81771148, 91849126), the National Key R&D Program of China (2018YFC1314700), Taishan Scholars Program of Shandong Province (tsqn20161078), and ZHANGJIANG LAB, Tianqiao and Chrissy Chen Institute, and the State Key Laboratory of Neurobiology and Frontiers Center for Brain Science of Ministry of Education, Fudan University.