
Abstract
Alzheimer’s disease (AD), a progressive neurodegenerative disorder characterized by memory loss and cognitive functions decline, is a leading cause for dementia and currently ranked as the sixth foremost cause of death. As of present, treatment of AD is symptomatic without convincing therapeutic benefits and new, effective, therapeutic agents are pursued. Due to massive loss of cholinergic neurons and decreased acetylcholine levels, cholinesterase inhibitors like galantamine, remain the backbone of pharmacological treatment of the disease. In the present study, using behavioral and biochemical methods, four newly synthesized galantamine derivatives, Gal 34, Gal 43, Gal 44, and Gal 46, were evaluated for a beneficial effect in a scopolamine model of dementia in mice. They were designed to have all the advantages of galantamine and additionally to inhibit β-secretase and exert favorable effects on plasma lipids. Behavioral tests included step-through inhibitory avoidance, T-maze, and the hole-board test, whereas biochemical evaluations involved assessment of acetylcholinesterase activity, brain monoamines levels, lipid peroxidation, catalase, glutathione peroxidase, and superoxide dismutase activities along with measurement of total glutathione. Results show that Gal 43, Gal 44, and, in particular, Gal 46 are especially effective in improving both short- and long-term memory and in the case of Gal 46 having a significant effect on exploratory activity as well. Although Gal 34 did not show behavioral effects as convincing as those of the other three galantamine derivatives, it demonstrated persuasive antioxidant and restorative capacities, making all four galantamine derivatives promising AD treatment agents and prompting further research, especially that in many of our studies they performed better than galantamine.
INTRODUCTION
Dementia, a heterogenous syndrome involving progressive impairments to memory, thinking, and behavior, has affected about 50 million people worldwide, with the number expected to increase to more than 131 million by 2050 [1, 2]. Alzheimer’s disease (AD), a progressive neurodegenerative disorder, characterized by memory loss and cognitive functions decline, is a leading cause for dementia and classified as the sixth foremost cause of death in US [3, 4], making it a focus of extensive research. Except for deposition of intraneuronal fibrillary tangles, composed of a highly phosphorylated form of the tau protein, and extracellular amyloid-β (Aβ) aggregates [5–7], massive loss of cholinergic neurons and decreased acetylcholine (ACh) levels [8], are among the most important features of the disease [9]. Thus, although in recent years different pharmacological strategies have been tried, acetylcholinesterase (AChE) inhibitors, blocking AChE and ensuring high ACh levels in brain [10], remain the backbone of pharmacological treatment of AD. At present four AChE inhibitors are approved and in use for the treatment of AD: tacrine, rivastigmine, galantamine, and donepezil with tacrine, the prototypical cholinesterase inhibitor for the treatment of the disease [11], discontinued over concerns of liver toxicity [12]. The use of none of the above inhibitors, however, translates into convincing therapeutic benefits [13], either delaying or stopping the progression of the disease, further pushing the design, synthesis, and biological evaluation of new and modified anticholinesterase drugs [14, 15].
Galantamine is a tertiary alkaloid isolated for the first time in 1952 from the Caucasian snowdrop Galanthus woronowii [16] and found in species of the Amaryllidaceae family. The Bulgarian chemist D. Paskov was the first to introduce galantamine for industrial use (under the name Nivalin) for the treatment of poliomyelitis paralysis, myasthenia gravis, myopathies, and others [17], prior to its current use as a licensed AD drug. Over the other AChE inhibitors, galantamine holds unique advantage influencing the cholinergic system in two favorable ways, i.e., inhibition of AChE along with allosteric modulation of nicotinic acetylcholine receptors (nAChR) [18, 19], resulting overall in increased availability of ACh. Additionally, galantamine acts further as a neuroprotector, first by stimulating the nAChR and inhibiting the Aβ- and glutamate-induced cytotoxicity [20–22] and second, by functioning as an anti-aggregatory agent due to its early direct interaction with soluble Aβ and inhibition of the formation of Aβ plaques [23]. In fact, galantamine turns out to be one of the most effective cholinesterase inhibitors as, although it is less potent than other marketed (for example donepezil), it shows significant improvement of cognitive performance in AD patients, comparable to or even exceeding that of the other inhibitors [24–26]. Thus, it is no surprise that galantamine appears an appealing template for new AD drugs. To further increase its potency, biological activity, and eventually therapeutic efficacy, different research groups have introduced changes in the structure of galantamine and synthesized new derivatives [27–31]. Especially attractive are derivatives with peptide fragments, which lower galantamine toxicity and confer entirely new properties on the galantamine molecule like inhibition of the activity of γ-secretase and β-secretase [32, 33], the enzymes responsible for the sequential cleavage of the amyloid-β protein precursor (AβPP), the build-up of Aβ, and further formation of amyloid plaques [34].
The aim of the present study was, using behavioral and biochemical methods, to evaluate four newly synthesized galantamine derivatives, Gal 34, Gal 43, Gal 44, and Gal 46 [35], in a scopolamine model of dementia in mice.
The newly synthesized derivatives of galantamine [35], comprising modified peptide moiety at position 11 of Norgalantamine (Fig. 1), were designed to have all the beneficial effects of galantamine and additionally to inhibit β-secretase, due to a shortened analogue of the β-secretase inhibitor OM99-2 (BocAsn-Leu-Ala-Val-OH) [36]. Moreover nicotinic/isonicotinic residues included in the derivatives were meant to exert favorable effects on plasma lipids as increased serum cholesterol and a low high-density lipoproteins (HDL) level in serum and cerebrospinal fluid are a risk factor for the development of AD [37] and a cholesterol-lowering drug BM15.766 has been shown to reduce Aβ pathology in a transgenic mouse model of AD [38]. Indeed, nicotinic acid (niacin) is one of the oldest antihyperlipidemic agents, known to lower cholesterol and triglyceride and increase HDL levels [39], properties retained by its derivatives as well [40–44]. In addition, the nicotinic/isonicotinic residues included in the derivatives were intended to facilitate blood-brain barrier penetration as data in the literature show that molecules containing residues of nicotinic acid readily cross the blood-brain barrier [45]. The new derivatives showed decreased toxicity compared to galantamine [35] and very good antioxidant properties with no side effects of their own [46].
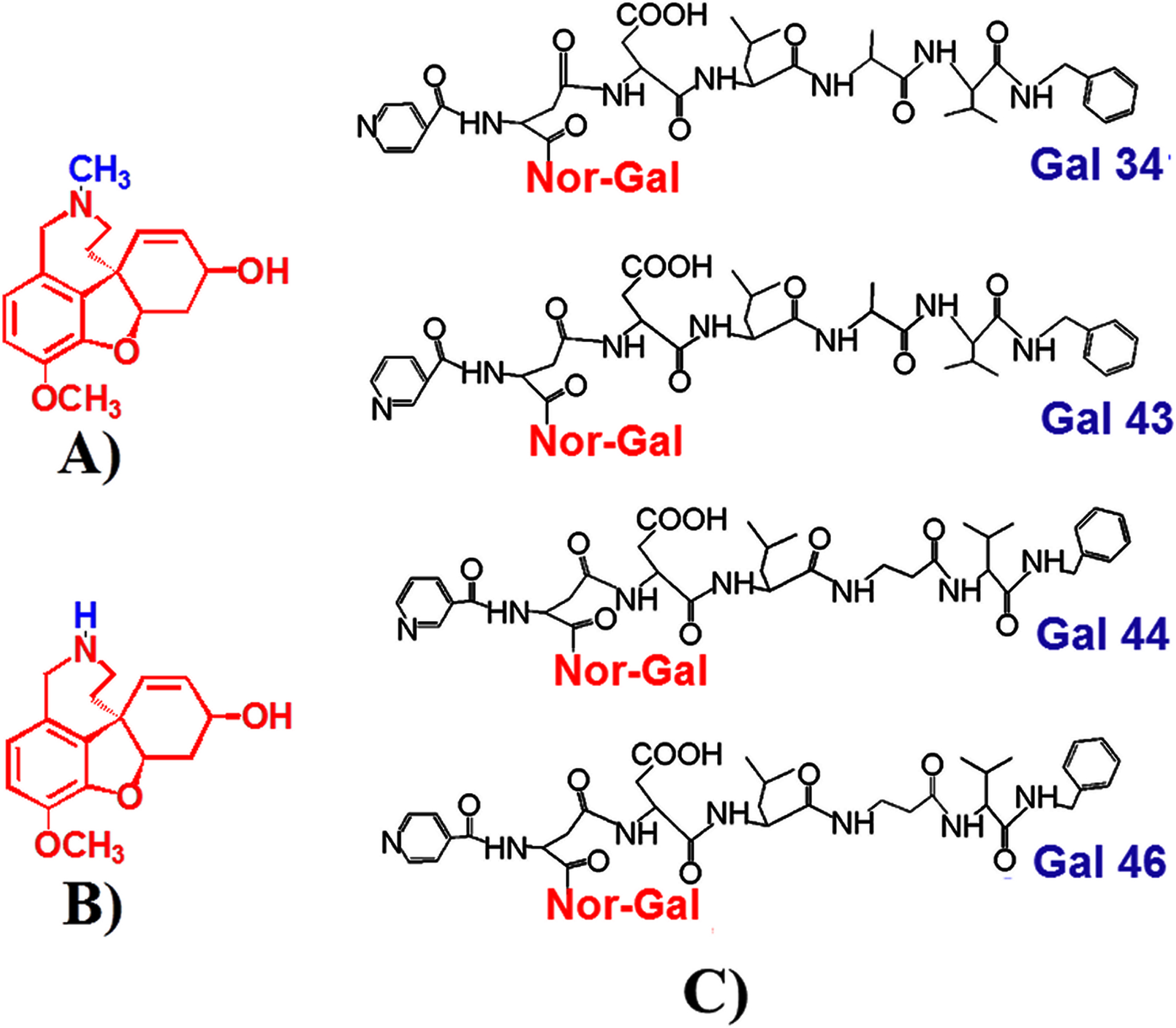
Chemical structures of galantamine (A), norgalantamine (N-demethylgalantamine) (B), and Gal 34, Gal 43, Gal 44, and Gal 46 with a peptide fragment and a pyridyl (as either nicotinoyl in the case of Gal 43 and Gal 44 or isonicotinoyl in the case of Gal 34 and Gal 46) moiety, attached to the N atom of norgalantamine (C).
Scopolamine, a non-selective competitive mACh Rs antagonist [47], which causes memory and cognitive damages both in animals and humans [48–55], has long been used to present a dementia model. Even more, it continues to prove an efficient tool to elicit AD pathology, based not only on induced cholinergic function deficiencies, but also on increased Aβ and phosphorylated tau production [56, 57], important hallmarks of AD. Besides, scopolamine induces a number of cellular alterations including changes in the monoaminergic systems, an impaired antioxidative defense system, increased oxidative stress, mitochondrial dysfunction, apoptosis, and neuroinflammation (e.g., [58]; for review, see [59]), pathological changes found in AD patients and in other AD models [60–67].
Studies of the effect of some of these peptide derivatives of galantamine on learning and memory in mice with scopolamine-induced amnesia have been published [68]. Independently of this, presented here experiments cover more detailed investigations of the effect of the compounds on the behavior and biochemistry of mice with scopolamine induced amnesia.
MATERIALS AND METHODS
Animals, drug treatment, and experimental design
Male albino ICR mice (18–20 g) were used. They were kept in plastic cages (n = 6) at a natural day/night light cycle and with free access to food and water (unless otherwise required by certain experiments). Mice were divided into seven experimental groups (n = 6), which were injected i.p. for nine consecutive days either with: saline (0.1 ml/10 g), the Control group; scopolamine (1 mg/kg), the Sco group as a model of dementia; scopolamine (1 mg/kg) and galantamine (1 mg/kg), the Sco + Gal group for comparison or scopolamine (1 mg/kg) and one of four galantamine derivatives (50 mg/kg) making the last four groups: Sco + Gal 34, Sco + Gal 43, Sco + Gal 44, and Sco + Gal 46. Behavioral tests such as the step-through inhibitory avoidance (ST), T-maze, and the hole-board test were carried out at different times (1st and 24th hour and at the 10th day for ST and at the 10th day for T-maze and the hole-board test) to assess learning and memory performance as well as locomotion and exploratory activity in each group. In the case of the ST test, scopolamine was injected 30 min before training and 30 min after galantamine or its derivatives in the groups, where those substances were applied together. Levels of oxidative stress were also estimated by measuring, in brain homogenates from the mice sacrificed on the 10th day, biochemical parameters such as lipid peroxidation, level of total glutathione, catalase, glutathione peroxidase, and superoxide dismutase activities. AChE activity and serotonin, dopamine, and noradrenaline were also assessed.
All the experiments with mice were carried out (conducted) in conformity with the institutional guidelines (Bioethics Committee at the Institute of Neurobiology, Bulgarian Academy of Sciences, Bulgaria) in compliance with the national and international laws and policies (the new Directive 2010/63/EU (22 September 2010) of the European Parliament and the Council of the European Union replacing the older Council Directive (86/609/EEC) on the protection of animals used for scientific purposes; the National Institute of Health (NIH) Guide for the Care and Use of Laboratory animals, NIH Publication No. 85-23, 1985, Bulgarian laws).
Step-through passive avoidance test
The test is routinely used to assess learning and memory deficits in rodents [69]. The apparatus consists of two separate chambers connected through a guillotine door. One chamber of the apparatus is illuminated and the other-dark. The floor of the dark chamber is a steel grid for delivering electric shocks by an isolated stimulator. At the time of the training phase, after a habituation period in the illuminated chamber (around 30 s and whenever the mouse is standing at the guillotine door with its back), the guillotine door is opened and, after the mouse enters the dark compartment, the door is closed and an unescapable electric foot shock (0.5 mA, 1 s, once) delivered. The time it takes the mouse to enter the dark chamber is recorded as initial latency. At the time of the test phase (recall) (1st and 24th hour and at the 10th day), mice are again placed one by one in the illuminated chamber and the time to enter the dark chamber (step-through latency (STL) with a cut-off time of 180 s) is recorded and assessed.
T-maze test
The test is commonly used to assess spatial working memory [70]. The T-maze apparatus consists of a start arm and two arms arranged in a T-shape. In the “spontaneous alternation” variant of the test, mice that are placed at the base of the T-maze tend to switch arms, visited during consecutive trials. In the “reinforced alternation” variant, used by us, the tendency of switching arms is reinforced by baiting sequentially the two different arms and using mildly food deprived mice. In the training phase, before the start of any pharmacological treatment, mice were trained to reach a certain level of correct choices. A modified protocol by Hussein et al. (2018) [71] was used. In short, after two days of handling the animals (when they are getting accustomed to the touch of the experimenter by being lifted by the body and set on his arm several times a day) on day 3 and 4 animals were habituated to the T-maze. During habituation the mice were allowed to freely explore the maze for 15 min. At day 3, scattered chocolate pieces over a short distance were found all along the maze to teach the mouse that food is available in the maze and to motivate the animal to move forward. At day 4, the pieces were found only at the ends of the two T-shaped arms to motivate the animal to choose and enter one of those arms. In the actual training part on day 5, the mouse was placed at the base of the starting arm in a compartment restrained by a guillotine door that was lifted after 10 s. In this first, forced trial, a chocolate piece was to be found only at the end of one randomly selected arm with free access, whereas the other arm was closed with a guillotine door. In the following nine trials, both arms were open, but baited was only the opposite arm of the arm baited in the previous trial. Recorded were the arm entries when the whole animal, including the tail tip, moved in one arm with successful entries being the ones, when the animal entered the baited arm and unsuccessful—when the other arm was entered. Performance in percentage was calculated as the number of correct choices divided by the number of total trials minus one and the ratio multiplied by 100. After each of the 10 trials the animal was lifted gently and returned for a next trial at the base arm after 5 min in the home cage. If the animal made an incorrect choice and entered an unbaited arm, it was allowed to continue exploring the maze until finding the reward piece in the other arm and eating it. The above training was repeated on day 6 and 7. From day one the mice were food deprived to 85%of their ad libitum body weight throughout the whole training phase and kept without any food for the last 12 h before the experiment on day 10.
Hole-board test
The hole-board test was conducted in a transparent box (50×50×40 cm), in which a plastic board with 16 equidistant holes (2.5 cm in diameter) and divided with a water-resistant marker into nine equivalent squares, was positioned 15 cm above the bottom of the box. On the 10th day after the pharmacological treatment, the mouse was placed in the center of the board and the following parameters were recorded for 5 min: the number of crossings of the lines defining the squares, the number of the head dips (pokings in holes up to the level of the ears) and the number of rearings (when the mouse is standing on its hind legs, raising its forelimbs off the ground, often momentarily [72].
Analytical methods
After the last behavioral test, mice were sacrificed by decapitation under mild CO2 inhalation and their brain tissues quickly removed on ice, cleaned with ice-cold saline, and stored at –20°C for biochemical assays. Prior to the assays, brain tissues were thawed at room temperature and 10%homogenates in 0.1 M sodium phosphate buffer (pH = 8) prepared. The homogenates were next centrifuged at 3000 rpm for 10 min and part of the supernatant used to determine the AChE activity, total protein, and total glutathione content. Another part of the supernatant was centrifuged at 12,000 g for 20 min and the obtained supernatant in turn used to determine catalase, glutathione peroxidase, and superoxide dismutase activities.
Acetylcholinesterase (AChE) activity was determined based on the Ellman method [73]. According to the method, the enzyme activity is measured by following the increase in the yellow color produced from thiocholine, when it reacts with the dithiobisnitrobenzoate ion. Thiocholine is produced under the catalytic action of AChE, when the substrate acetylthiocholine is added. In our experiments, 100μl of the supernatant were mixed with 100μl 0.01M 5,5′-dithio-bis(2-nitrobenzoic acid) (DTNB)(Ellman’s reagent), 2.9 ml 0.1 M phosphate buffer and 20μl 0.075M freshly prepared acetylthiocholine iodide. 500μl of the reaction mixture were injected into a Semi-auto Chemistry Analyzer and kinetics of the reaction monitored for 3 min at 405–412 nm. After determination of the total protein content by the Biuret method (12.5μl of the supernatant were mixed with 1 ml ready-made reagent containing 0.6 M NaOH, 32 mM KNaC4H4O6, 20 mM KI, 12 mM CuSO4, incubated for 10 min at 37°C and the protein content determined at 546 nm), the enzyme activity in IU/l, was presented as IU/ml/mg protein.
Lipid peroxidation (LPO) was determined by the reaction of malondialdehyde (MDA), an end production of lipid peroxidation, with thiobarbituric acid (TBA) to form a colorimetric (532 nm)/fluorometric (λex = 532/λem = 553 nm) product, proportional to the MDA present. A Lipid Peroxidation (MDA) Assay Kit (Cat No MAK085, Sigma-Aldrich) was used and, after proten determination, LPO expressed as nmoles MDA/mg protein.
Total glutathione (GSH) content, expressed in ng/mg protein, was measured by the method of Rahman et al. (2006) [74], based on oxidation of GSH by DTNB to form the yellow derivative 5′-thio-2-nitrobenzoic acid (TNB), measurable at 412 nm.
Cu, Zn-superoxide dismutase (SOD) activity was determined based on the assay developed by Peskin and Winterbourn (2017) [75] and using hypoxantine, xanthine oxidase, and the water soluble tetrazolium WST-1. The superoxide anion, formed upon the oxidation of hypoxanthine by xanthine oxidase, reduces WST-1 to a yellow-colored formazan dye, whose absorbance at 440 nm is proportional to the amount of the superoxide anion. SOD catalyzes the dismutation of the superoxide anion into hydrogen peroxide and molecular oxygen. Thus, SOD activity as an inhibition activity for the formation of the formazan dye can be quantified by measuring the decrease in the color development at 440 nm. SOD activity was assayed using the 19160 SOD Determination Kit, Sigma –Aldrich and presented in U/mg protein (one unit of SOD activity being the amount of the enzyme producing 50%inhibition of the tetrazolium reduction).
Catalase (CAT) activity determination is based on the measurement of the hydrogen peroxide substrate remaining after the action of catalase. The colorimetric method uses a substituted phenol (3,5-dichloro-2-hydroxybenzenesulfonic acid), which couples oxidatively to 4-aminoantipyrine in the presence of hydrogen peroxide and horseradish peroxidase (HRP) to give a red quinoneimine dye (N-(4-antipyryl)-3-chloro-5-sulfonatep-benzoquinone-monoimine) that absorbs at 520 nm. Catalase Activity Assay Kit (Cat. No CAT 100) is used and catalase activity presented in U/mg protein.
Glutathione peroxidase (GPx) activity was measured based on the oxidation of glutathione to oxi-dized glutathione (GSSG) catalyzed by GPx, which is then coupled to the recycling of GSSG back to glutathione utilizing glutathione reductase (GR) and NADPH (β-Nicotinamide Adenine Dinucleotide Phosphate, Reduced). The decrease in NADPH absorbance measured at 340 nm during the oxidation of NADPH to NADP+ is indicative of GPx activity, since GPx is the rate limiting factor of the coupled reactions. GPx activity was assayed using the Glutathione Peroxidase Cellular Activity Assay Kit (Catalog Number CGP1, Sigma-Aldrich) and presented in U/mg protein (one unit of GPx activity being the amount of the enzyme causing the formation of 1.0μmol of NADP+ from NADPH per minute).
Determination of dopamine (DA), serotonin (5-HT), and noradrenaline (NA)
DA, 5-HT, and NA were assayed using the method for rapid determination of norepinephrine, dopamine, and serotonin in the same region developed by Jacobowitz and Richardson (1978) [76]. Briefly, the mice brains were weighed and homogenized in 5 ml butanol and an appropriate amount of 0.01 N HCl, so that the tissue water plus 0.01 N HCl would equal 0.75 ml (assuming tissue weight is 70%water) in 5 ml of butanol. Next the homogenates were centrifuged at 1500 rpm for 10 min and NA, DA and 5-HT determined. For quantitative determination of DA and NA phosphate buffer was used and 2 ml of the butanol supernatant were added to 1.5 ml 0.1 M phosphate buffer in ice water. For quantitative determination of 5-HT HCl was used and 2 ml of the butanol supernatant were added to 5 ml heptane and 0.5 ml of 0.1N HCl in ice water. After vortexing, both mixtures were centrifuged at 3000 rpm for 10 min. After centrifugation, NA and DA were turned into fluorophores by adding 1 ml of the aqueous layer of the centrifugate to 0.25 ml Versene (Versene prepared as 4 g disodium ethylenediaminetetra-acetic acid in 100 ml distilled water, pH = 6-6.5). Next, at precisely 2 min intervals, the following substances were added: 0.20 ml iodine (iodine prepared as 4.8 g potassium iodide and 0.25 g iodine in 100 ml distilled water); 0.25 ml alkaline sodium sulfite (alkaline sodium sulfite prepared as 2.5 g sodium sulfite in 100 ml 4 N NaOH) and 0.3 ml 5N CH3COOH. After keeping the test tubes in boiling water for 5 min, then in ice water for 1 min, NA fluorescence was immediately determined (excitation 385 nm/emission 485 nm) and DA read 20 min later at 320 nm/385 nm. 5-HT was turned into a fluorophore by adding 0.3 ml of the aqueous layer of the respective centrifugate to 0.2 ml ortho-phthaldialdehyde (OPT) followed immediately by 1.5 ml of 10N HCl. After keeping the test tube in boiling water for 10 min and cooling it at room temperature, 5-HT was determined at 360 nm/470 nm. Fluorescence readings for DA, NA, and 5-HT were converted [76] and the three neurotransmitters presented as μg per gtissue.
Statistical analysis
Results were expressed as mean values±standard error of the mean. Data were analyzed with the use of one-way analysis of variance (ANOVA) and the Tukey post hoc comparison test. Differences were considered significant at p < 0.05.
RESULTS
Effects of galantamine derivatives on AChE activity in brain in a scopolamine model of dementia in mice
In Fig. 2 it is shown that scopolamine treatment (1 mg/kg, i.p.) for 9 consecutive days statistically significantly increases AChE activity with 45%versus control (p < 0.001). Galantamine fails to restore control values and there is a statistically significant difference of 32%between the Sco + Gal and the control group (p < 0.05). Although values for scopolamine-induced AChE activity in the presence of the four galantamine derivatives remain higher than control values neither of the Sco + Gal 34, Sco + Gal 43, Sco + Gal 44, and Sco + Gal 46 groups shows a statistically significant difference with the control group (p > 0.05).
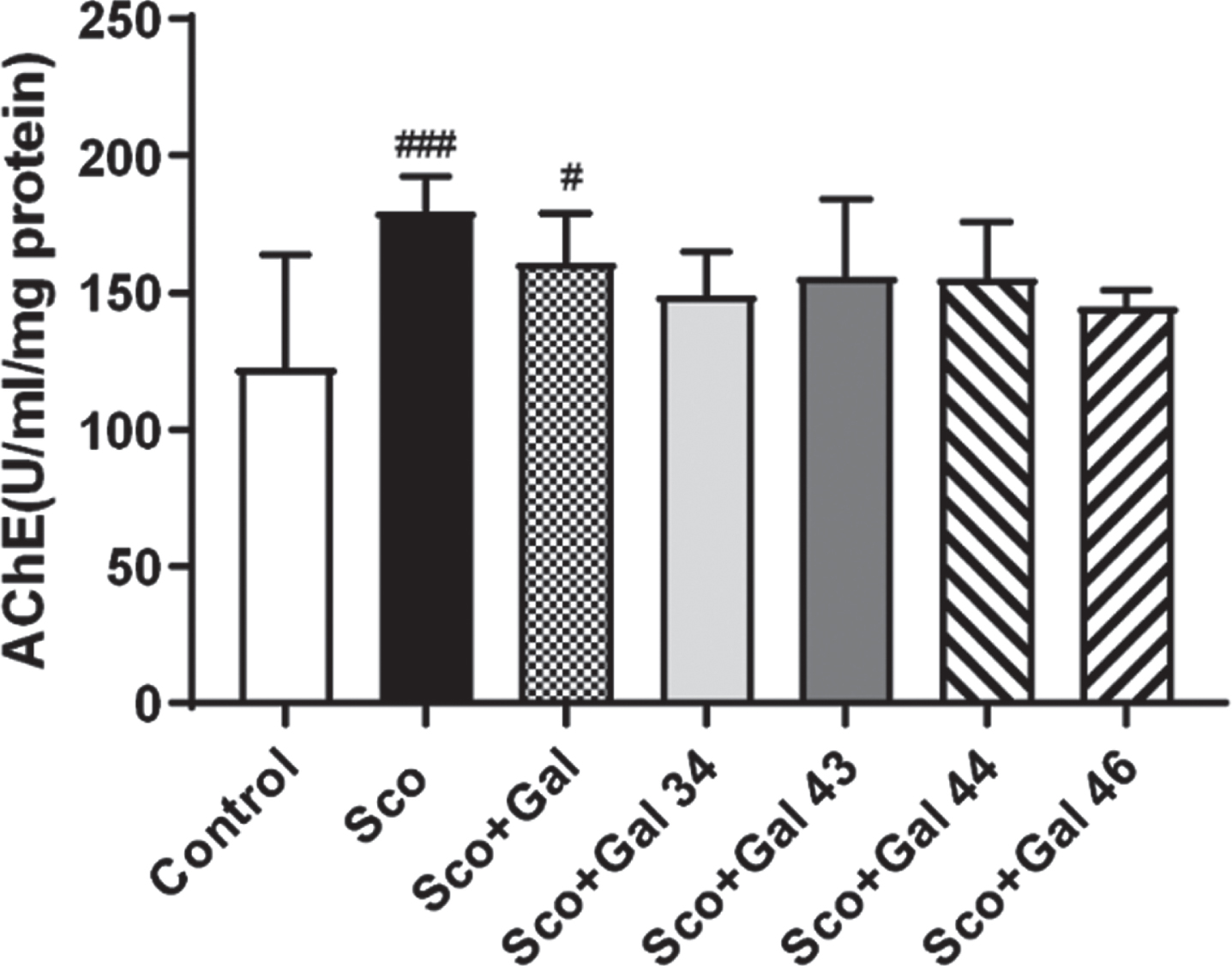
Effects of galantamine and four galantamine derivatives Gal 34, Gal 43, Gal 44, and Gal 46 on brain acetylcholinesterase (AChE) activity in a scopolamine model of dementia in mice. AChE activity was assessed after 9 days of treatment with saline (control group); with scopolamine, 1 mg/kg (Sco group); with scopolamine (1 mg/kg) and galantamine (1 mg/kg) (Sco + Gal group) and with scopolamine (1 mg/kg) and one of the four galantamine derivatives (50 mg/kg) (Sco + Gal 34; Sco + Gal 43; Sco + Gal 44; and Sco + Gal 46). Data are presented as mean values±SEM (n = 6) and significant differences among experimental groups are denoted with “#” whenever groups were compared with the saline (control) group and denoted with “*” whenever groups were compared with the dementia (Sco) group. The number of “#” or “*” above bars was assigned as follows: # or * at p < 0.05; ## or ** at p < 0.01; ### or *** at p < 0.001 and #### or **** at p < 0.0001.
Effects of galantamine derivatives on learning and memory in a scopolamine model of dementia in mice
In Fig. 3A-C, it is shown that scopolamine treatment decreases the step-through latency (STL) at 1 h, 24 h, and 10 days with a declining effect over time, the decrease for Control versus Sco being 90%at 1 h (p < 0.0001), 46%at 24 h (p < 0.05), and 23%at 10 days (p > 0.05). At 1 h, although increasing the STL value, galantamine failed to restore the control value with a difference in Control versus Sco + Gal of 53%(p < 0.01). While Gal 34 also failed to restore the control value (with a difference in Control versus Sco + Gal 34 of 67%, p < 0.001), Gal 43, Gal 44, and Gal 46 showed favorable effects and no statistically significant difference with the control was registered. At 24 h again Gal 34 failed to restore control values, while galantamine, Gal 43, Gal 44, and Gal 46 showed favorable effects and no statistically significant difference with the control. At 10 days, although scopolamine decreased the control STL values by 23%, no statistically significant difference with the control was observed, an effect seen for the rest of the groups, when compared with controls, as well. In Fig. 3D, it is shown that, although scopolamine decreases T-maze performance by 28%on the 10th day, the effect did not reach statistical significance (p > 0.05). No statistically significant differences were recorded also between the control and any of the Sco + Gal, Sco + Gal 34, Sco + Gal 43, Sco + Gal 44, or Sco + Gal 46 groups.
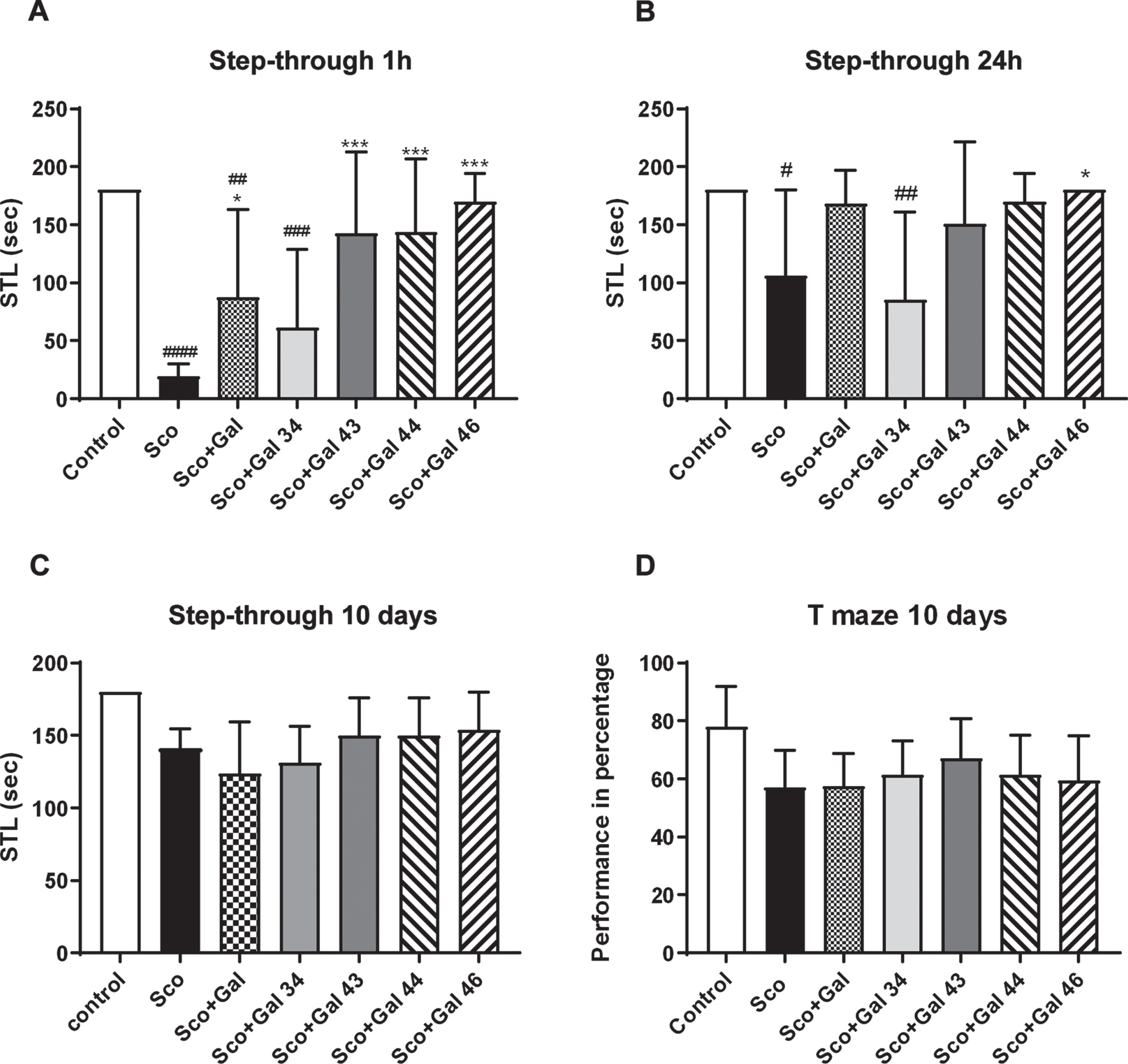
Effects of galantamine and four galantamine derivatives Gal 34, Gal 43, Gal 44, and Gal 46 on learning and memory in a scopolamine model of dementia in mice. Step-through latency is assessed at 1 h, 24 h, and on the 10th day (A, B, and C, respectively) and T-maze performance is evaluated on the 10th day (D) for the control, Sco, Sco + Gal, Sco + Gal 34; Sco + Gal 43; Sco + Gal 44 and Sco + Gal 46 groups. Data are presented as mean values±SEM (n = 6) and significant differences among experimental groups are denoted with “#” whenever groups were compared with the saline (control) group and denoted with “*” whenever groups were compared with the dementia (Sco) group. The number of “#” or “*” above bars was assigned as follows: # or * at p < 0.05; ## or ** at p < 0.01; ### or *** at p < 0.001 and #### or **** at p < 0.0001.
Effects of galantamine derivatives on locomotion and exploratory activity in a scopolamine model of dementia in mice
In Fig. 4A, it is shown that, although in the Sco, Sco + Gal, Sco + Gal 34, and Sco + Gal 46 groups the number of line crossings, as a measure of locomotion, is increased and this number decreased in the Sco + Gal 44 and Sco + Gal 43 groups, compared to control, neither of these effects reached statistical significance.

Effects of galantamine and four galantamine derivatives Gal 34, Gal 43, Gal 44, and Gal 46 on locomotion and exploratory activity in a scopolamine model of dementia in mice. The number of line crossings (A), head dips (B), and rearings (C) is assessed on the 10th day for the control, Sco, Sco + Gal, Sco + Gal 34; Sco + Gal 43; Sco + Gal 44 and Sco + Gal 46 groups. Data are presented as mean values±SEM (n = 6) and significant differences among experimental groups are denoted with “#” whenever groups were compared with the saline (control) group and denoted with “*” whenever groups were compared with the dementia (Sco) group. The number of “#” or “*” above bars was assigned as follows: # or * at p < 0.05; ## or ** at p < 0.01; ### or *** at p < 0.001 and #### or **** at p < 0.0001.
In Fig. 4B, it is shown that Sco decreases and Gal 46, in the presence of scopolamine, quite substantially increases the number of head dips. However, those effects do not reach statistical significance and for the groups Sco and Sco + Gal 46, as well as for the rest of the groups, i.e., Sco + Gal, Sco + Gal 34, Sco + Gal 43, and Sco + Gal 44, no significant differences were detected when compared to the control group (p > 0.5).
In Fig. 4C, it is shown that Sco and Gal 44, in the presence of scopolamine, decrease the number of rearings. Galantamine, Gal 34, Gal 43, and especially Gal 46, in the presence of scopolamine, increase the number of rearings. However, neither of those effects reached statistical significance and for the groups Sco and Sco + Gal 44, as well as for the rest of the groups, i.e., Sco + Gal, Sco + Gal 34, Sco + Gal 43, and Sco + Gal 46, no significant differences were detected in comparison with the control group (p > 0.5).
Effects of galantamine derivatives on oxidative stress and antioxidant capacity in a scopolamine model of dementia in mice
Results for the effects of the four galantamine derivatives on oxidative stress are presented in Fig. 5. In Fig. 5, it is shown that scopolamine increases MDA levels in brain by 34%(Sco versus Control, p < 0.01). Galantamine and all galantamine derivatives, especially Gal 34, which decreases MDA levels to the most, restore control values with neither of the groups Sco + Gal, Sco + Gal 34, Sco + Gal 43, Sco + Gal 44, and Sco + Gal 46 showing a significant difference with the control group (p > 0.5).
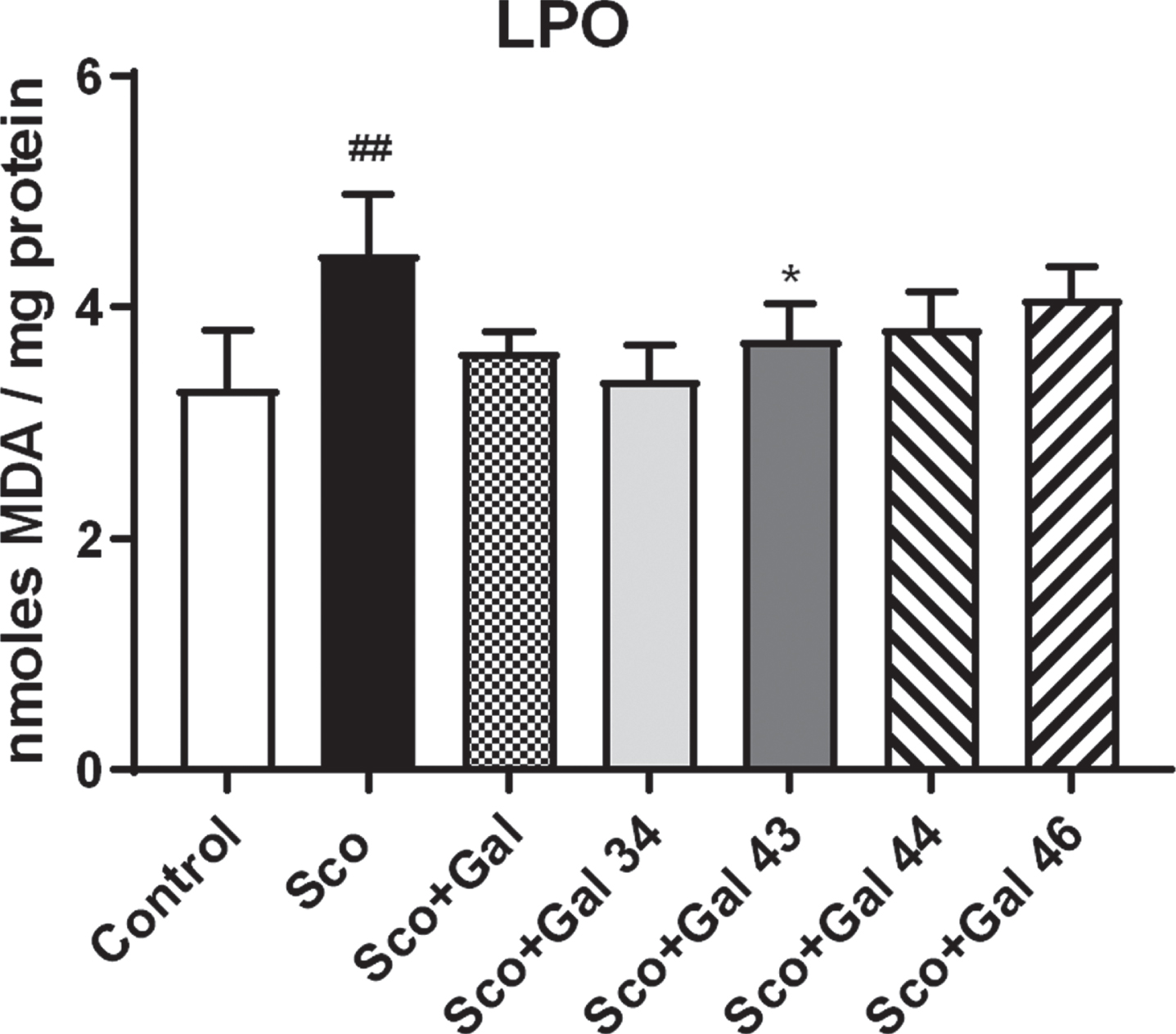
Effects of galantamine and four galantamine derivatives Gal 34, Gal 43, Gal 44, and Gal 46 on oxidative stress in a scopolamine model of dementia in mice. Oxidative stress is evaluated after 9 days of treatment by assessing lipid peroxidation through measuring MDA levels in the groups injected with saline (control group); with scopolamine, 1 mg/kg (Sco group); with scopolamine (1 mg/kg) and galantamine (1 mg/kg) (Sco + Gal group) and with scopolamine (1 mg/kg) and one of the four galantamine derivatives (50 mg/kg) (Sco + Gal 34; Sco + Gal 43; Sco + Gal 44 and Sco + Gal 46). Data are presented as mean values±SEM (n = 6) and significant differences among experimental groups are denoted with “#” whenever groups were compared with the saline (control) group and denoted with “*” whenever groups were compared with the dementia (Sco) group. The number of “#” or “*” above bars was assigned as follows: # or * at p < 0.05; ## or ** at p < 0.01; ### or *** at p < 0.001 and #### or **** at p < 0.0001.
Results for the effects of the four galanta-mine derivatives on brain antioxidant capacity are presented in Fig. 6. In Fig. 6A, it is shown that although Sco and Gal, in the presence of scopolamine, slightly decrease GSH brain levels and Gal 43, Gal 44, and Gal 46 slightly increase brain GSH levels, none of the above shows any significant difference with the control group (p > 0.5). On their background, however, Gal 34 reveals notable antioxidant capacity increasing significantly, by 25%, GSH levels (p < 0.5) compared with the control group and by 59%compared with the scopolamine group (p < 0.001). In Fig. 6B-D, the effects of galantamine and the four galantamine derivatives on the activities of the main antioxidant enzymes are shown. In Fig. 6B, it is shown that scopolamine decreases SOD activity by 31%(p < 0.5). While both galantamine and all the galantamine derivatives show no significant difference with the control, it is Gal 43 and again Gal 34 that practically fully restore the control values for SOD activity. In line with the latter, the two galantamine derivatives, in the presence of scopolamine, show a statistically significant difference with the scopolamine group (p < 0.5). Regarding CAT and GPx activities, scopolamine decreases both of them, with 23%for CAT and 14%for GPx (Fig. 6C, D), although the effects are not statistically significant. In the case of CAT, galantamine and Gal 43 almost completely restore the control values (Fig. 6C), whereas, in the case of GPx, Gal, Gal 43, and especially Gal 34, even exceed them (Fig. 6D).
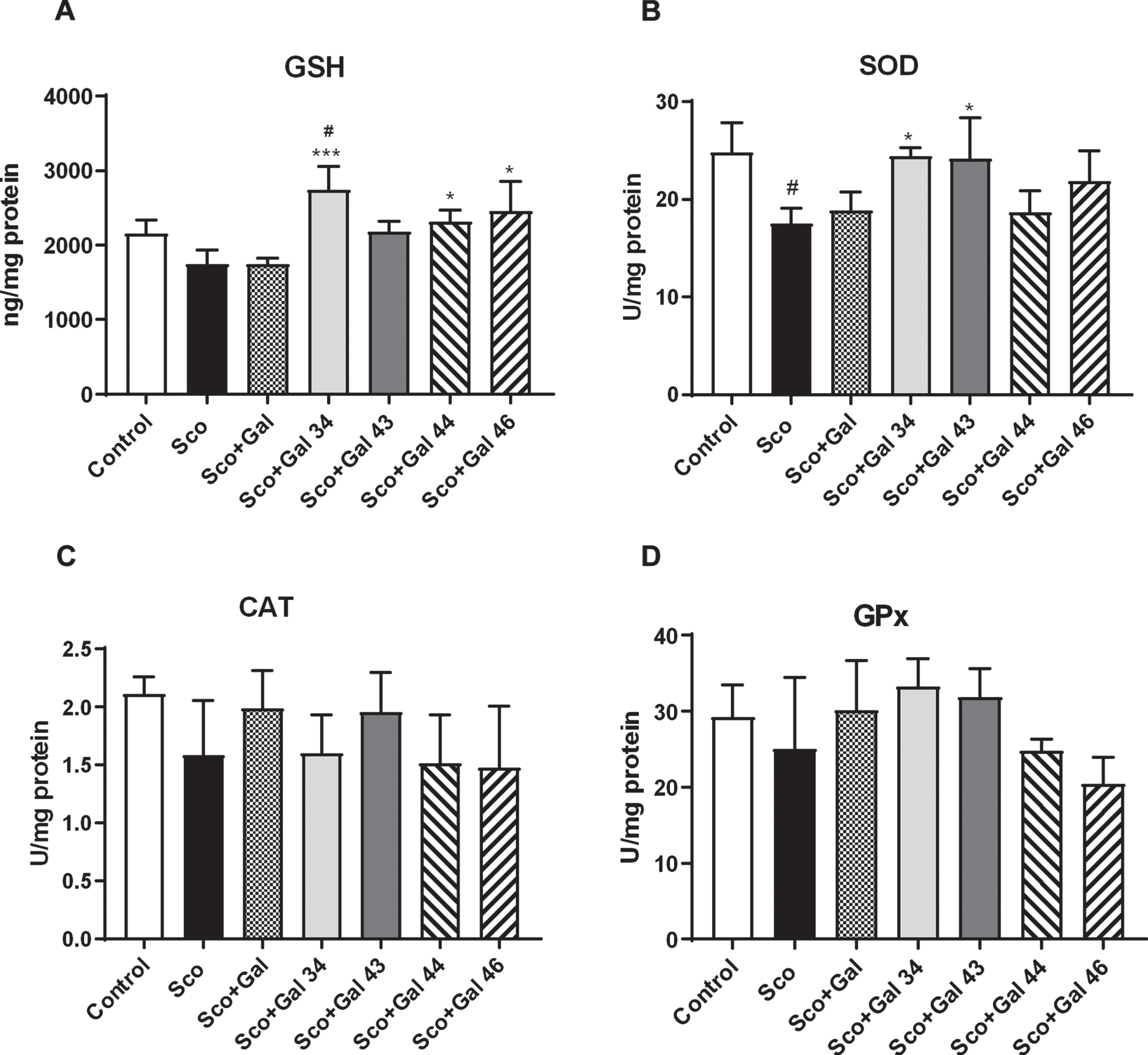
Effects of galantamine and four galantamine derivatives Gal 34, Gal 43, Gal 44, and Gal 46 on brain antioxidant capacity in a scopolamine model of dementia in mice. Antioxidant brain capacity is evaluated after 9 days of treatment by assessing GSH levels (A), SOD activity (B), CAT activity (C), and GPx activity (D) in the groups injected with saline (control group); with scopolamine, 1 mg/kg (Sco group); with scopolamine (1 mg/kg) and galantamine (1 mg/kg) (Sco + Gal group) and with scopolamine (1 mg/kg) and one of the four galantamine derivatives (50 mg/kg) (Sco + Gal 34; Sco + Gal 43; Sco + Gal 44 and Sco + Gal 46). Data are presented as mean values±SEM (n = 6) and significant differences among experimental groups are denoted with “#” whenever groups were compared with the saline (control) group and denoted with “*” whenever groups were compared with the dementia (Sco) group. The number of “#” or “*” above bars was assigned as follows: # or * at p < 0.05; ## or ** at p < 0.01; ### or *** at p < 0.001 and #### or **** at p < 0.0001.
Effects of galantamine derivatives on brain biogenic amines (DA, 5-HT, and NA) in a scopolamine model of dementia in mice
Results for the effects of the four galantamine derivatives on brain biogenic amines are presented in Fig. 7. In Fig. 7A, it is shown that Sco, galantamine, and all galantamine derivatives increase serotonin levels and this effect for Gal 34, Gal 44, and Gal 46 is statistically significant; for Gal 34, the increase is 300%(Control versus Sco + Gal 34 p < 0.0001), for Gal 44 180%(Control versus Sco + Gal 44, p < 0.01), and for Gal 46 160%(Control versus Sco + Gal 46, p < 0.05). In Fig. 7B, it is shown that Sco statistically significantly decreases NA levels (Control versus Sco p < 0.01). While galantamine, Gal 34, and Gal 44 fail to restore control values and there is a statistically significant difference Control versus Sco + Gal (p < 0.01), Control versus Sco + Gal 34 (p < 0.01), and Control versus Sco + Gal 44 (p < 0.05), Sco43 and Sco 46 restore those and the groups Sco + Gal 43 and Sco + Gal 46 show no statistically significant difference with the control (Control versus Sco + Gal 43, p > 0.05; Control versus Sco + Gal 46, p > 0.05). In Fig. 7C, it is shown that scopolamine reduces immensely, with more than 95%, DA levels (Control versus Sco, p < 0.0001), an effect that cannot be compensated neither by galantamine nor by any of the studied galantamine derivatives (Control versus Sco + Gal, p < 0.0001; Control versus Sco + Gal 34, p < 0.0001; Control versus Sco + Gal 43, p < 0.0001; Control versus Sco + Gal 44, p < 0.0001; Control versus Sco + Gal 46, p < 0.0001).

Effects of galantamine and four galantamine derivatives Gal 34, Gal 43, Gal 44, and Gal 46 on brain biogenic amines in a scopolamine model of dementia in mice. Biogenic amine levels were evaluated after 9 days of treatment by assessing 5-HT levels (A), NA levels (B), and DA levels (C) in the groups injected with saline (control group); with scopolamine, 1 mg/kg (Sco group); with scopolamine (1 mg/kg) and galantamine (1 mg/kg) (Sco + Gal group) and with scopolamine (1 mg/kg) and one of the four galantamine derivatives (50 mg/kg) (Sco + Gal 34; Sco + Gal 43; Sco + Gal 44 and Sco + Gal 46). Data are presented as mean values±SEM (n = 6) and significant differences among experimental groups are denoted with “#” whenever groups were compared with the saline (control) group and denoted with “*” whenever groups were compared with the dementia (Sco) group. The number of “#” or “*” above bars was assigned as follows: # or * at p < 0.05; ## or ** at p < 0.01; ### or *** at p < 0.001 and #### or **** at p < 0.0001.
DISCUSSION
In the present study, using behavioral and biochemical methods, we examined the neuroprotective effects of four galantamine derivatives, Gal 34, Gal 43, Gal 44, and Gal 46, in scopolamine-induced Alzheimer-type dementia in mice. Scopolamine was applied i.p for 9 consecutive days at a dosage of 1 mg/kg in male mice. Behavioral tests included step-through inhibitory avoidance (ST), T-maze, and the hole-board test to assess learning and memory performance as well as locomotion and exploratory activity. Biochemical evaluations involved assessment of AChE activity, of brain monoamines levels, of lipid peroxidation, of CAT, GPx, and SOD activities along with measurement of the level of total GSH. As discussed in detail below, all four galantamine derivatives were effective in improving various scopolamine-induced deficiencies, in many cases performing better than galantamine (Table 1), staging those substances as potential AD treatment agents and for sure urging their future studies.
Comparison of the effects of galantamine with the effects of Gal 34, Gal 43, Gal 44, and Gal 46
“-” denotes an effect worse than that of galantamine, “+” –an effect better than that of galantamine, and “x” denotes an effect same as that of galantamine in the respective study. AChE, acetylcholinesterase, LPO- lipid peroxidation, GSH-total glutathione, SOD-superoxide dismutase, CAT-catalase, GPx- peroxidase, 5-HT-serotonin, NA-noradrenaline, DA-dopamine.
Scopolamine is well known to induce cholinergic dysfunction. Except for being a muscarinic receptor antagonist, it decreases acetylcholintransferase activity relevant to the synthesis of ACh [77] and in numerous studies has been shown to increase AChE (e.g., [77–82]) and butyrylcholinesterase activities [80, 83], relevant to the degradation of ACh. Our study confirmed the AChE-activating effect of scopolamine.
Unlike galantamine, which partially decreased scopolamine-induced increases in AChE activity, but failed to return the activity to its control values, all the four galantamine derivatives, and especially Gal 34 and Gal 46, antagonized the scopolamine-induced AChE increase, neither of them showing statistically significant difference with the control group. The slightly better effects of Gal 34 and Gal 46 compared to the other galantamine derivatives could be due to the isonicotinic residues in their molecules (Gal 43 and Gal 44 have instead nicotinic residues), possibly ensuring more favorable steric interaction with AChE and its better block.
Of importance is to note that AChE activity has a detrimental effect in AD not only by decreasing ACh levels, but also due to inducing neurotoxicity. First, AChE interacts with Aβ and promotes amyloid fibril formation [84] and second, AChE-Aβ complexes are more toxic than amyloid fibrils alone [85]. Anti-cholinesterase drugs like propidium and decamethonium were able to block amyloid formation [86]. Thus, efficiently blocking AChE activity, like the one we have shown, is of many values and can turn an effective, complex strategy in the treatment of AD.
It is well established that, among the many neuro-transmitter systems, the cholinergic plays a fundamental role in memory. The forebrain cholinergic system is involved in arousal and attention, induced by novelty and sensory stimulation, in sustained attention, needed in complex operant tasks, in information acquisition and memory formation by making possible the encoding of new information of various types including spatial information [87, 88]. In line with the above, scopolamine, as an anticholinergic drug, produces a well described impairment in maintaining attention along with deficits in processing of information and the acquisition and holding of new knowledge and thus is especially successful in damaging acquisition/learning, short term and working memory, including spatial memory [54]. Consequently, scopolamine has been used as a standard tool in various behavioral tests for assessment of learning and working memory, including spatial memory. Some of the classical behavioral methods for evaluating the above are the passive avoidance test and different types of maze, among which extensively used are the Morris water maze, the T-maze, and the radial arm maze [77,89–92, 77,89–92].
In our study, step-through passive avoidance and the T-maze tests were used. Passive avoidance, also referred to as inhibitory avoidance, as the animal learns to avoid a punishment, is an emotionally arousing paradigm that involves: an explicit, associative component to the context; an operant-like conditioning component to the shock, since the animal may avoid the aversive stimulus, and a spatial memory component, since the animal remembers the location where the noxious stimulus was given during acquisition [93]. Passive avoidance allows to study both short-term and long-term memory as recall can be performed at different times after acquisition. In our experiments, scopolamine was applied 30 min before training and recall was considered at 1 h and 24 h, followed by further application of scopolamine for 8 more days and assessment of long-term memory on the 10th day. In agreement with earlier reports [81, 94–100], training under the effect of scopolamine produced a significant passive avoidance performance deficit expressed in the significant difference of the step through latency times between the Sco and control groups at 1 and 24 h after training. Over time, the effect of scopolamine decreased, and at 24 h it was less than at 1 h and at 10 days less than at 24 h, compatible with the current view that, whereas some mechanisms are involved in both [101], different molecular instruments may be needed to form short-term and long-term memory [102]. The latter statement has been convincingly demonstrated in inhibitory avoidance tasks with the use of pharmacological treatments that differentially block short- and long-term memory[101–105].
All four galantamine derivatives and in particular Gal 43, Gal 44, and Gal 46, showed favorable effects, with Gal 43, Gal 44, and Gal 46 restoring both short-term and long-term memory and consistently (at 1 h and 10 days) revealing better results than galantamine.
The T-maze alongside with administration of scopolamine is another common behavioral method to study the effects of compromised cholinergic functioning on learning and memory [106, 107]. It offers a simple test of spatial working memory as mice learn to alternate between the arms of the maze over successive trials in order to receive a food reward [107]. The T-maze task was chosen by us as, apart from offering a simple and straightforward test of spatial working memory (one of the few behavioral methods to assess this particular type of memory [70]), it is well recognized that mice perform worse than rats in water mazes [108].
Previous studies show that application of scopolamine, including chronic application at dosages of 1 mg/kg in mice as in our study, results in spatial memory impairments as judged by changes in T-maze parameters such as increase in the latency time for the mice to enter the preferred arm; decrease in the number of entries and time spent in the preferred arm in a T-maze with rewarded alternation and reduction in the percentage of alterations in a T-maze with spontaneous alterations [109, 110]. Correspondingly in our experiments, on the 10th day after application of scopolamine, we observed reduction in the percentage of T-maze performance, although not reaching statistical significance. Similar to the passive avoidance task, all four galantamine derivatives, with Gal 43 standing out, showed favorable effects increasing T-maze performance and revealing better results than galantamine.
Thus, it could be concluded that all the studied galantamine derivatives and especially Gal 43, Gal 44, and Gal 46 with Gal 43 standing out, demonstrate improvements in scopolamine-induced deficits in associative memory (improving recall in both short- and long-term passive avoidance tasks) and spatial working memory. As AD is characterized by impaired recall and recognition memory, impaired working memory, disorientation in time and place (Table 1) [111], the studied galantamine derivatives present themselves as promising candidates for treating AD memory damages.
The advantageous effects of the above galantamine derivatives are observed at the background of no impaired locomotor activity. Data in the literature show that scopolamine has been generally found to increase locomotor activity (e.g., [112, 113]), although a decrease has also been reported (e.g., [92, 114]). In our study, although scopolamine increased locomotion, the effect did not reach statistical significance. Among all tested galantamine derivatives, only Gal 44 decreased locomotion, but the number of line crossings as a measure of locomotor activity, was not any statistically significant different from the control value, when mice were injected with physiological solution.
The hole-board method is being currently widely used in many fields of behavioral pharmacology to evaluate the attraction of rodents toward novelty and their exploratory activity [115]. Unlike the open field test, which according to some authors evokes a strong fear response, “masking” unbiased rodent exploration examination [116], the hole-board task gives a possibility to distinguish between locomotion and directed exploration.
The number of head dips as well as rearings is indicative of interest in novelty and exploratory activity [72, 118]. According to Lever et al. (2006) [72], rearing can be used to examine encoding, storage, consolidation and retrieval processes, and long-term plasticity models, associated with spatial/contextual learning functions of the hippocampus, without requiring that the animal performs navigation tasks, which rest upon this function. In our study scopolamine decreased the number of both head dips and rearings. While the decrease was not statistically significant, it should be noted that among all tested substances Gal 46 performed best and not only restored, but even exceeded controlvalues.
Oxidative stress is an affecting factor and another characteristic feature of AD [60–63, 119–121]. Scopolamine-induced memory impairments, in turn, have been shown to be related to increased oxidative stress in the brain.
Oxidative stress could be judged by the level of MDA, a product of lipid peroxidation and by the activity of the cellular antioxidant enzymes such as CAT, GPx, and SOD. As reduced glutathione acts as an antioxidant either directly or as a substrate for GPx, determining the level of GSH is also a useful marker. Abundant reports in mice show that scopolamine treatment, including with 1 mg/kg as in our study, markedly increases MDA levels, decreases CAT, GPx, and SOD activities, and reduces GSH amounts [77, 122–124]. As no exception, our study also revealed all the features of oxidative stress induced by scopolamine such as enhanced MDA levels, reduced CAT, GPx and SOD activities, and diminished GSH amounts. While all galantamine derivatives staged themselves very well and, in the cases with GSH levels and SOD activity, better than galantamine, it was Gal 34 that was especially noticeable and consistently presented very good effects by influencing simultaneously all oxidative stress parameters. Gal 34 remarkably increased GSH levels, improved SOD and GPx activities with best efficacy, and most efficiently decreased MDA levels. Although it did not raise the GSH levels as much as Gal 34 did, Gal 43 was the best in restoring the activities of all antioxidant enzymes and performed almost as well as Gal 34 in reducing the levels of MDA augmented by scopolamine.
Apart from the cholinergic system, other neurotransmitter systems, such as the serotonergic, noradrenergic. and dopaminergic systems, are also involved in learning and memory processes [125]. Subsequently changes in those systems have been observed in AD as well.
Reports demonstrate reductions of 5-HT and its metabolites in postmortem brains of AD patients [126, 127]. In fact, both 5-HT and 5HIAA cortical levels correlate negatively with the number of neurofibrillary tangles suggesting that the impairment of the 5-HT-ergic system parallels disease progression [128]. Other changes accompanying AD are diminished serotonergic innervation of cortical and subcortical regions, decreased activity of TPH, the principal enzyme in the biosynthesis of 5-HT, decreased 5-HT transporter activity and changes in the density of 5-HT receptors (for review, see [129]).
Regarding its participation in learning and memory, although the serotonergic system is demonstrated to be relevant in mediating acquisition, consolidation, and retention of learning and thus in memory formation [125, 130], its precise function is not fully elucidated and findings are controversial. On one hand, treatment with 5-HT receptor agonists produces impairment of learning and memory in various behavioral tasks ([131]; for review, see 130]); neurotoxin- and chemically induced depletion of 5-HT improves complex maze- and active avoidance learning [132, 133] and extensive data in the literature show that 5-HT antagonists have memory-enhancing effects [130, 134–136]. On the other hand, reports demonstrate that 5-HT agonists enhance memory [137] and 5-HT depletion hampers active avoidance learning [138]. Even the widely popular notion that the serotonergic and cholinergic systems are mutually inhibitory [139, 140] is opposed by results of some studies indicating that those systems act in concert with each other [141–143]. Furthermore, blocking of serotonin uptake, which increases synaptic concentrations of 5-HT, betters mnemonic task performance in animals and humans (Table 6 from 144, 145, 146). In addition, acute tryptophan depletion, a precursor of 5-HT, causes impairment in memory consolidation [147], also in patients with AD [148]. So, whether an increase in 5-HT would facilitate memory processes, remains obscure. We believe that the augmented 5-HT levels observed in the presence of the galantamine derivatives (and especially Gal 34, Gal 44, and Gal 46) in our study possibly exert favorable effects on the scopolamine-induced deficiencies in memory and especially for the spatial and passive avoidance tasks. In support of this, experiments, that have examined the effects of different pharmacological manipulations of the serotonergic and cholinergic systems on measures of learning and memory, have shown, that combined treatment with the 5-HT uptake inhibitor alaproclate and oxotremorine, a muscarinic agonist, enhances retrieval of passive avoidance behavior in mice at doses which, when given individually, were too low for either drug to produce an effect [149]. Additionally, citalopram, a selective serotonin reuptake inhibitor that has been shown to raise extracellular concentration of 5-HT in hippocampus [150], increases ACh release in the rat frontal cortex and dorsal hippocampus [151, 152] and reverses scopolamine-induced spatial memory deficits [153].
The most prominent effect of AD on the noradrenergic system is the loss of up to 70%of locus coeruleus (LC) noradrenergic neurons [66]. Reductions in the number of LC noradrenergic neurons and NA deficiency correlate with increased cortical amyloid plaque and neurofibrillary tangles loads as well as with cognitive deterioration [154–156]. In fact, it has been suggested that reductions in LC neurons parallel even better the onset and duration of AD than cholinergic nucleus basalis degeneration [157, 158].
Except for visceral functions (respiration, cardiovascular function, secretion of hormones), sleep-wake cycle, emotion, mood, motor and pain control, the noradrenergic system regulates a wide range of cognitive functions, including working memory, learning, and attention as well as memory consolidation and retrieval [159–163]. Thus, the beneficial effects on memory of the examined galantamine derivatives (and especially Gal 43 and Gal 46, which perform best and restore NA levels to control values), aside from lowering AChE activity, could be due to increasing NA amounts. Indeed, enhancement of brain NA neurotransmission reduces both neuroinflammation and cognitive decline [164] and selective loss of noradrenaline exacerbates early cognitive dysfunction and synaptic deficits in APP/PS1 mice [165].
Reductions in DA, DA metabolites, and DA receptors have been revealed (observed) in AD [129]. Mouse models of AD also present reduced DA release in the hippocampus and insular cortex [166, 167].
Similar to the noradrenergic system, the dopaminergic system is involved in the regulation of a diversity of physiological functions like the reward system, mood and emotions, control of movement and pain control, visceral functions and last but not least cognitive functions involving motivation, attention, working memory, motor planning, thinking and abstract reasoning, temporal analysis, speech and language, learning, and social behavior [66]. Therefore, not unexpectedly, DA depletion in rats has been reported to lead to impaired performance in passive avoidance and maze tasks [168–170]. All galantamine derivatives, including galantamine itself, did not restore the decreased by scopolamine levels of DA, indicating no participation of the neurotransmitter in their favorable effects on memory.
In conclusion, our results show that Gal 43, Gal 44, and in particular Gal 46 are especially effective in improving both short-term and long- term memory and in the case with Gal 46 having a significant effect on exploratory activity as well. The slightly better behavioral effects of Gal 46 compared to the other galantamine derivatives could at least partially be explained with the favorable increase in NA and 5-HT induced by Gal 46, its very good antioxidant capacity (especially through increasing scopolamine-reduced GSH levels and SOD activity), and last but not least with effectively blocking the AChE activity, the latter at least in part due to the flexibility of the Gal 46 carbon chain imposed by the isonicotinic residue and β-Ala and ensuring more favorable steric interaction with AChE. It should be noted, that although Gal 34 did not show behavioral effects as convincing as those of the other three galantamine derivatives, it demonstrated persuasive antioxidant and restorative capacities (LPO, GSH, GPx, SOD), making all four galantamine derivatives (for sure not excluding Gal 43, which showed best results in the spatial memory test) promising AD treatment agents. As derivatives of galantamine and similar to the other cholinesterase inhibitors, if used as drugs, the four studied compounds might possibly cause gastrointestinal problems, nausea, and vomiting and their low solubility in water might present issues with absorption and bioavailability. However, the favorable effects shown in the present study, in many cases superseding those of galantamine and the very low toxicity compared to galantamine shown in vitro in cell cultures and in vivo on mice [35] (insertion of the peptide fragments in the galantamine molecule has been reported to lower toxicity about 100 times [35]), renders them attractive AD drug candidates to be possibly moved to clinical trial stages.
Footnotes
ACKNOWLEDGMENTS
This work was supported by the Bulgarian National Science Fund, project DN-03-8/2016 “Galantamine and 4-aminopyridine derivatives, containing a peptide motif, with expected effect on Alzheimer’sdisease and multiple sclerosis”. The authors also ack-nowledge the Bulgarian Ministry of Education and Science for their financial aid through the National Programme “European Scientific Networks”, project D01-278/05.10.2020 “Drug molecule”.