
Abstract
Background:
Sphingosine 1-phosphate (S1P) and ceramides have been implicated in the development of Alzheimer’s disease. Apolipoprotein E (ApoE) isoforms are also involved in the development of Alzheimer’s disease.
Objective:
We aimed at elucidating the potential association of the ApoE isoforms with sphingolipid metabolism in the central nervous system.
Methods:
We investigated the modulations of apolipoprotein M (apoM), a carrier of S1P, S1P, and ceramides in Apoeshl mice, which spontaneously lack apoE, and U251 cells and SH-SY5Y cells infected with adenovirus vectors encoding for apoE2, apoE3, and apoE4.
Results:
In the brains of Apoeshl mice, the levels of apoM were lower, while those of ceramides were higher. In U251 cells, cellular apoM and S1P levels were the highest in the cells overexpressing apoE2 among the apoE isoforms. The cellular and medium contents of ceramides decreased in the order of the cells overexpressing apoE3 > apoE2 and increased in the cells overexpressing apoE4. In SH-SY5Y cells, apoM mRNA and medium S1P levels were also the highest in the cells overexpressing apoE2. The cellular contents of ceramides decreased in the order of the cells overexpressing apoE3 > apoE2 = apoE4 and those in medium decreased in the order of the cells overexpressing apoE3 > apoE2, while increased in the cells overexpressing apoE4.
Conclusion:
The modulation of apoM and S1P might partly explain the protective effects of apoE2 against Alzheimer’s disease, and the modulation of ceramides might be one of the mechanisms explaining the association of apoE4 with the development of Alzheimer’s disease.
INTRODUCTION
It is well established that sphingolipids, represented by ceramides, sphingosine, and sphingosine 1-phosphate (S1P), represent important bioactive lipids involved in the development of various diseases in humans. Among the diseases, both basic and clinical studies have demonstrated the involvement of sphingolipids in the development of diseases of the central nervous system (CNS), including Alzheimer’s disease [1], Parkinson’s disease [2], amyotrophic lateral sclerosis [3], and multiple sclerosis [4]. In general, among the sphingolipids, ceramides and sphingosine exhibit cytotoxicity and induce apoptosis [5], while S1P possess neuroprotective properties, promoting cell survival and protecting against mitochondrial dysfunction, inflammation, and oxidative stress [6].
Sphingolipids are largely regulated by the circulating lipoproteins. Ceramides and sphingosine are contained in abundance in lipoproteins [7, 8] and their levels are modulated by treatment with statins [9, 10]. S1P is also carried in abundance on high-density lipoprotein (HDL) via apolipoprotein M (apoM) [11] and the plasma S1P concentrations are modulated by the levels of proteins related to lipoprotein metabolism, such as the low-density lipoprotein (LDL) receptor protein [12, 13] and cholesteryl ester transfer protein [14]. Up to the present, the involvement of apoM in Alzheimer’s disease is unclear. Only limited papers reported that the polymorphisms within the apoM gene were associated with Alzheimer’s disease [15] and that apoM decreased in the cerebrospinal fluids of Alzheimer’s disease [16]. Considering that apoM augments the properties of S1P in promoting cell survival and protecting against mitochondrial and endothelial dysfunction [17–22], the impaired expression of apoM might be involved in the pathogenesis of Alzheimer’s disease.
In the CNS, in contrast to the case in the circulation, apolipoprotein E (apoE)-rich HDL is the major lipoprotein [23]. A number of studies have reported the close involvement of apoE, especially some isoforms of apoE, in the pathogenesis of neurological diseases [24], such as Alzheimer’s disease; apoE4 has been reported to be associated with an increase in the risk, and apoE2 has been reported to be associated with a decrease in the risk of Alzheimer’s disease [25, 26]. At present, many mechanisms have been proposed to explain these clinical associations, such as apoE4 accelerates the accumulation of amyloid-β [27], whereas apoE2 decreases the accumulation of amyloid-β [28]. ApoE isoforms are also reported to influence tau pathology and synaptic integrity and plasticity [29]. Among the proposed mechanisms, one of the important is that apoE isoforms regulate the lipid homeostasis in the CNS. For example, it has been demonstrated that apoE4 impairs cholesterol efflux of apoE-rich HDL in the cerebrospinal fluid [30].
In the circulation, apoE can affect not only the blood levels of cholesterol, but also the blood levels of other lipids, including sphingolipids. Sharman et al. demonstrated, using apoE2-, apoE3-, and apoE4-knockin mice, that the plasma levels of ceramides were higher in apoE2-knockin mice than in apoE3- and apoE4-knockin mice [31]. In addition, in a recent clinical study, the levels of several species of ceramides were found to be lower in subjects with the ApoE2 allele, and higher in those with the apoE4 allele [32]. In the CNS, on the other hand, the modulation by apoE and/or apoE isoforms of the levels of ceramides remains controversial. Sharman et al. reported the absence of any significant differences in the brain levels of ceramides among apoE2-, apoE3-, and apoE4-knockin mice [31], whereas den Hoedt et al. reported that the ceramide levels in the brain were lower in apoE-knockout and apoE4-knockin mice [33]. In regard to S1P, we found that in apoE-deficient mice, the plasma S1P levels were higher and that S1P bound to HDL/apoM was cleared from the circulation through the LDL receptor, possibly with apoE serving as the ligand [12]. Until date, there are no reports of studies conducted to investigate the associations of apoE/apoE isoforms with the levels of S1P in the CNS.
Considering these backgrounds, we hypothesized that apoE isoforms might be involved in modulating apoM/S1P and ceramide metabolism in the CNS, which might represent a novel mechanism to explain the involvement of apoE isoforms in the development of CNS disorders, especially Alzheimer’s disease. Therefore, in the present study, we attempted to elucidate the influences of apoE and apoE isoforms on the sphingolipid metabolism and apoM levels in the brain, using Apoeshl mice, which spontaneously lack apoE, U251 cells, a glioblastoma cell line, SH-SY5Y cells, a neuroblastoma cell line, and adenovirus vectors encoding for apoE2, apoE3, and apoE4.
MATERIALS AND METHODS
Animal experiments
C57BL/6 (WT) mice and B6.KOR/StmSlc-Apoeshl (Apoeshl) mice, which spontaneously lack apoE, were obtained from Japan SLC (Shizuoka, Japan). Ten-week-old WT or Apoeshl mice were denied access to food for 6 h, and then plasma samples and brain samples were collected. All the animal experiments were conducted in accordance with the guidelines for animal care and with the approval of the Animal Care Committee of the University of Tokyo (P17-074).
Cell experiments
U-251 cells were purchased from JCRB Cell Bank and HepG2 cells and SH-SY5Y cells were from ATCC (Manassas, VA). U-251 cells and HepG2 cells were cultured in DMEM (D5796, Sigma-Aldrich Co.) supplemented with 10% fetal bovine serum (FBS; 554-02655; BioSera Inc., Orange, CA) and 1% penicillin/streptomycin (168-23191; WAKO Pure Chemical Industries). SH-SY5Y cells were cultured in DMEM/Ham’s F12 (D8437, Sigma-Aldrich Co.) supplemented with 10% FBS and 1% penicillin/streptomycin.
To examine the modulation of sphingolipid levels by overexpression or knockdown of ApoE, U-251 cells, SH-SY5Y cell, and HepG2 cells were infected with adenovirus vectors at an MOI of 25 or were administered with siRNA against apoE (sc-29708; Santa Cruz Biotechnology) or with control siRNA (sc-37007; Santa Cruz Biotechnology) using lipofectamine RNAi MAX (13778075; Invitrogen Co., Carlsbad, CA). After 48 h, the medium was replaced with serum-free medium, and a further 24 h later, the culture media and cellular components were collected and subjected to analyses. as described below. The adenovirus vectors encoding for apoE2, apoE3, and apoE4 (Ad-apoE2, Ad-apoE3, and Ad-apoE4) and the control β-galactosidase-expressing adenovirus vector (Ad-LacZ) have been described previously [34].
Western blotting
The proteins of the cerebral cortex of the brains were extracted using RIPA lysis buffer (25 mM Tris HCl [pH 7.6], 1% NP-40, 0.1% SDS, 150 mM NaCl, 0.02% sodium deoxycholate, 1 mM orthovanadate, 1 mM PMSF, and protease inhibitor cocktail [Roche, Mannheim, Germany]). The nuclear proteins were prepared as described previously [35, 36]. Western blotting was performed using 10μg of the cellular and brain proteins, 2.5μg of the nuclear proteins, or 0.02μL of plasma, according to a previously described standard method. To analyze the proteins in the media, the amount of medium subjected to western blotting was adjusted according to the cellular protein level. The following antibodies were used: anti-apoE antibody (AB740 and AB947; Chemicon International Inc., Temecula, CA), anti-mouse apoM antibody (A00954, GenScript Co, Piskataway, NJ), anti-β-actin antibody (PM053, MBL, Nagoya, Japan), anti-mouse albumin antibody, anti-RXRα antibody, anti-LXRα antibody, anti-PPARβ antibody, anti-PPARγ antibody, anti-HNF-4α antibody, anti-Lamin A/C antibody (sc-11795, sc-515929, sc-271064, sc-74517, sc-7196, sc-374229, sc-20681; Santa Cruz Biotechnology), PPARδ antibody (ab137724; Abcam plc, Cambridge, UK), anti-human SIRT1 antibody and anti-mouse SIRT1 antibody (2493 and 9475, Cell Signaling Technology, Beverly, MA). The anti-human apoM antibody was developed and validated in a previously reported paper [37]. The intensities of the bands were measured using the ImageJ software (from the NIH). The bands from different gels were compared after normalization using the same sample run on both gels as the internal standard. The intensities of the apoM and apoE bands were adjusted to those of albumin or β-actin and those of nuclear proteins were adjusted to those of Lamin A/C.
Reverse-transcriptase PCR
The total RNA extracted from the cerebral cortex of the brains or cells using the GenElute Mammalian Total RNA Miniprep kit was subjected to reverse transcription with the ReverTra Ace qPCR RT Master Mix. Quantitative PCR was performed using an ABI 7300 Real-Time PCR System (Applied Biosystems), with hybridization probes and primers purchased from Applied Biosystems: murine apoM (Mm00444525_m1), murine apoE (Mm01307193_g1), murine GAPDH (Mm99999915_g1), human apoM (Hs01597780_g1), human apoE (Hs00171168_m1), and human GAPDH (Hs02786624_g1). The reaction conditions were as follows: incubation for 10 min at 95°C, followed by 40 cycles at 95°C for 15 s and 60°C for 1 min. The expression levels of the genes of interest were normalized to those of the endogenous control GAPDH mRNA.
Measurement of lysophospholipids, diacyl phospholipids, and eicosanoids/related compounds
We measured the levels of ceramide and sphingosine using an LC8060 system consisting of a quantum ultra triple quadrupole mass spectrometer (Shimadzu, Japan), including S1P, dihydrosphingosine 1-phosphate (dhS1P), six ceramide species (Cer d18 : 1/16 : 0 [C16], Cer d18 : 1/18 : 0 [C18 : 0], Cer d18 : 1/18 : 1 [C18 : 1], Cer d18 : 1/20 : 0 [C20], Cer d18 : 1/22 : 0 [C22], Cer d18 : 1/24 : 0 [C24]), sphingosine (Sph), and dihydrosphingosine (dhSph), as described and validated previously [38–40]. The brain contents were normalized to the brain protein levels, and the medium and cellular contents were adjusted to the cellular protein levels.
Statistical analysis
The results are expressed as dot plots. The differences between two groups were assessed using the unpaired Student’s t test with Welch’s correction, while differences among four groups were assessed using one-way ANOVA, followed by Scheffe’s test for post-hoc comparisons. p values of less than 0.05 were deemed as being statistically significant.
RESULTS
The plasma levels of S1P, dhS1P, and ceramides were higher in the Apoe shl mice
First, we compared the plasma levels of apoM and sphingolipids between the WT mice and Apoeshl mice; in the Apoeshl mice, apoE was not detected, and no change of the apoM levels was observed, consistent with our previous report [12] (Fig. 1A, B). In regard to the sphingolipids, the plasma levels of S1P, dhS1P, and ceramides were higher in the Apoeshl mice, while the plasma levels of Sph and dhSph were not significantly different from those in the WT mice (Fig. 1C–G).
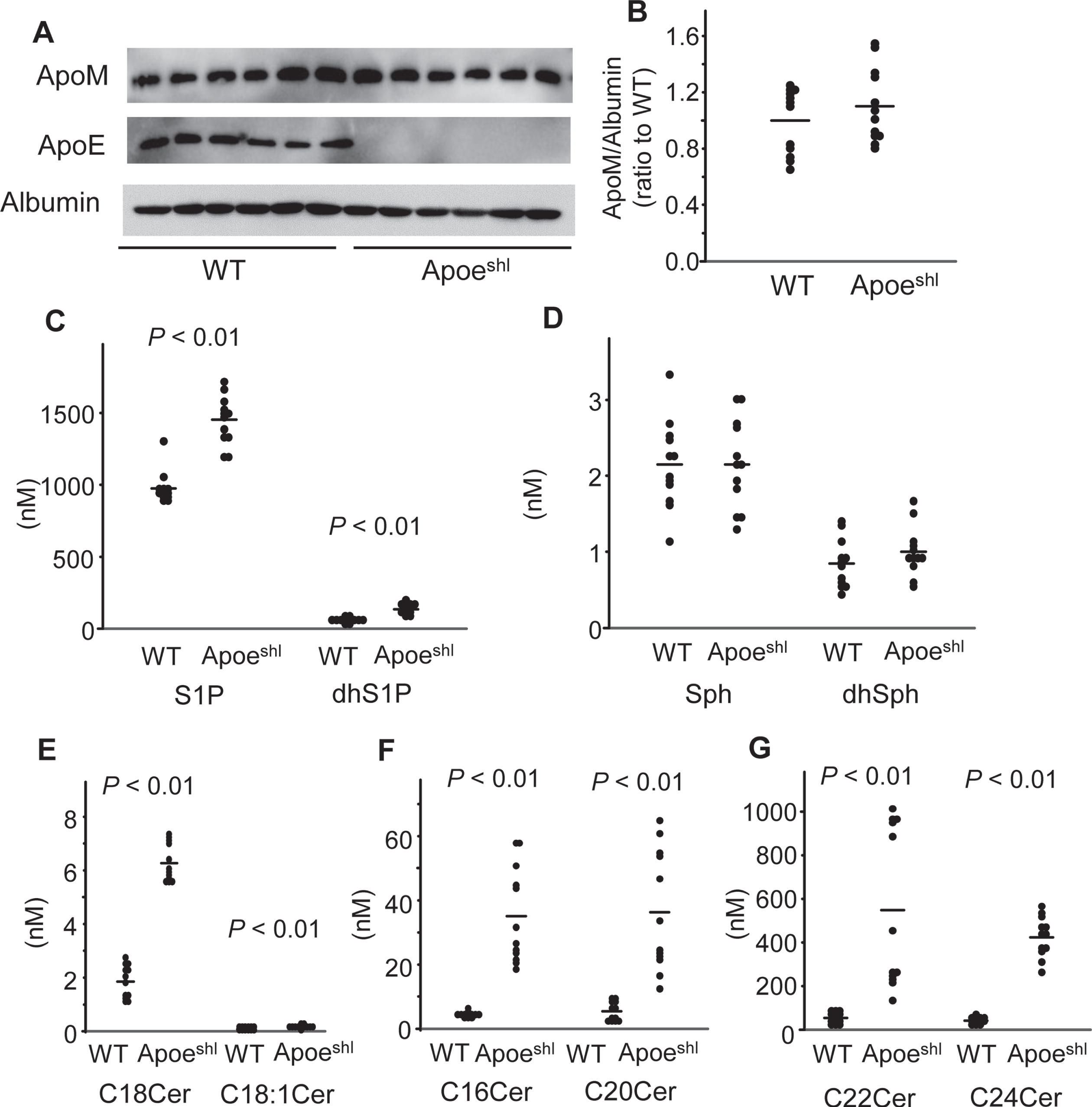
Modulation of the plasma levels of apoM and sphingolipids in the Apoeshl mice. Ten-week-old C57BL/6 (WT) mice and B6.KOR/StmSlc-Apoeshl (Apoeshl) mice were denied access to food for 6 h, and plasma samples were collected for measurement of the levels of apolipoproteins and sphingolipids (n = 12). A, B) Western blots for plasma apolipoproteins (A) and the bands for apoM were quantified with the ImageJ software (B). C–G) Plasma sphingolipid levels. The horizontal bars represent the means of independently analyzed samples.
The contents of apoM were lower and those of ceramides were higher in the brains of the Apoe shl mice
Next, we compared the brain contents of apoM and sphingolipids between the WT mice and Apoeshl mice. In the Apoeshl mice, there exist several amounts of apoE mRNA in brain, while the protein levels of ApoE were too low to be detected by the western blot analyses (Fig. 2A–C). The levels of apoM protein, a carrier and modulator of S1P [11, 37], were lower, while those of apoM mRNA were not significantly different from those in the WT mice (Fig. 2A–C). In regard to the brain contents of sphingolipids in the Apoeshl mice, the contents of C16, C18 : 0, C20, C22, and C24 ceramides, Sph, and dhSph were higher, whereas those of C18 : 1 ceramide, S1P, and dhS1P were not significantly different from those in the WT mice (Fig. 2D–H).

Modulation of the levels of apoM and sphingolipids in the brains of Apoeshl mice. The brains were collected from the mice analyzed in Fig. 1 for measurement of the levels of apolipoproteins and sphingolipids (n = 12). A, B) Western blots for plasma apolipoproteins (A) and the bands for apoM were quantified with the ImageJ software, using albumin as the internal standard (B). C) Real-time PCR for apoM and apoE mRNA using GAPDH as the internal standard. D–H) The brain contents of the sphingolipids adjusted to the protein levels. The horizontal bars represent the means of independently analyzed samples.
Overexpression of apoE in the U251 cells modulated the cellular contents of apoM and sphingolipids, depending on the apoE isoform overexpressed
We infected U251 cells with Ad-LacZ (LacZ-U251 cells), Ad-apoE2 (E2-U251 cells), Ad-apoE3 (E3-U251 cells), and Ad-apoE4 (E4-U251 cells). The apoE protein and mRNA levels were not significantly different among the E2-, E3-, and E4-U251 cells (Fig. 3A, C, E). In regard to the expression of apoM, overexpression of apoE decreased the expression levels of apoM at both the protein and mRNA levels (Fig. 3A, B, D). The apoM protein-lowering potency of apoE appeared to be in the order of apoE3 = apoE4 > apoE2 (Fig. 3B). Concordantly, the levels of S1P were lower in the E3- and E4-U251 cells than the LacZ- and E2-U251 cells (Fig. 3F). The levels of dhS1P remained unchanged (Fig. 3F). The levels of Sph, were lower in the E3-U251 cells, followed by the E2-U251 cells, and those of dhSph were higher in the E4-U251 cells (Fig. 3G). Regarding the modulation of ceramides, the levels of C20, C22, and C24 ceramides were lower in the E3-U251 cells and those of C20 ceramide were lower in the E2-U251 cells than the LacZ-U251 cells, while the levels of C16, C18 : 1, C24 ceramides were higher in the E4-U251 cells (Fig. 3H-J). Considering these results together, the cellular ceramide-lowering effects of apoE were in the order of apoE3 > apoE2 and apoE4 possesses the cellular ceramide-increasing effects.
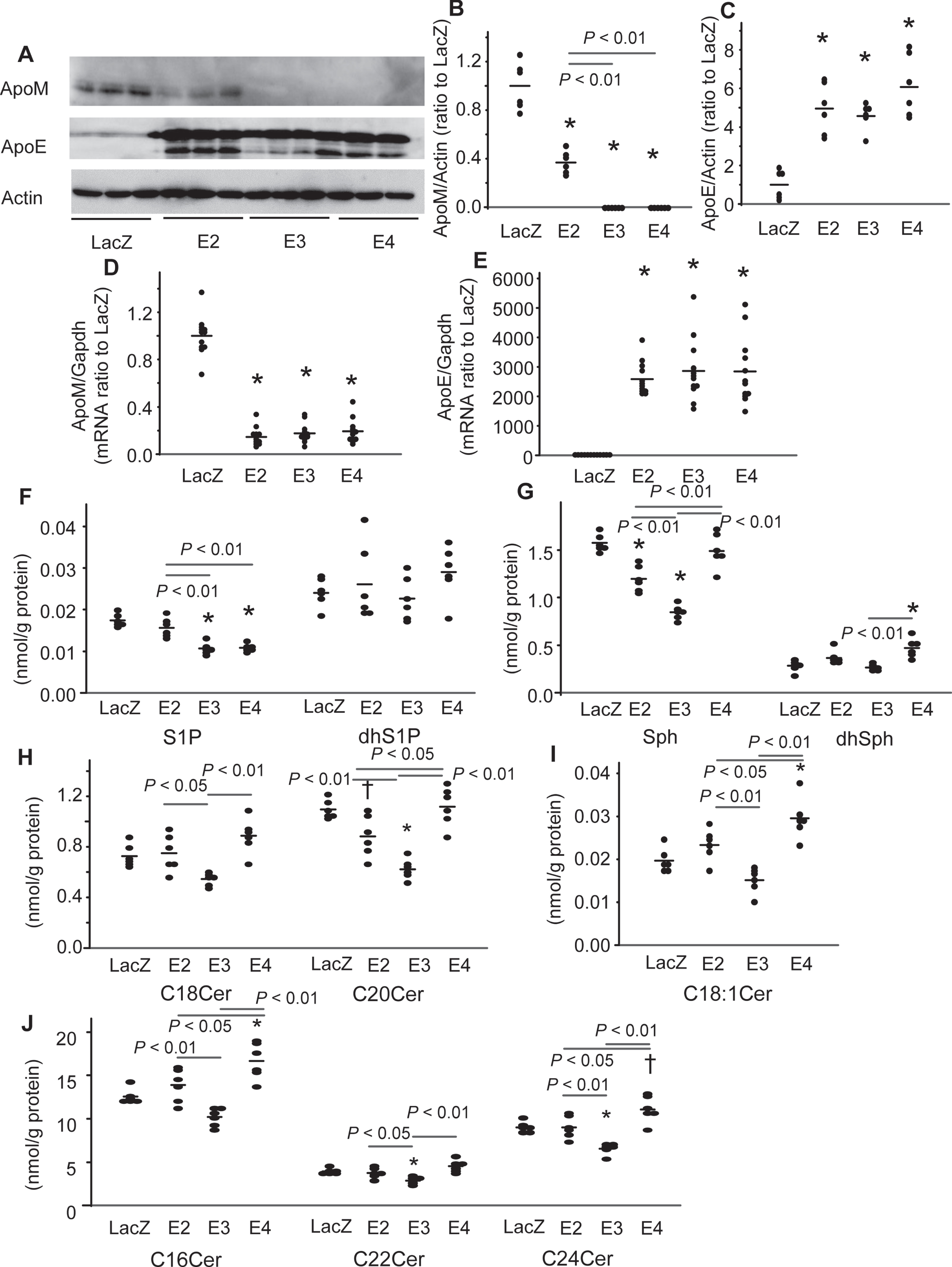
Modulation of the cellular levels of apoM and sphingolipids by overexpression of apoE isoforms in the U251 cells. U251 cells were infected with Ad-LacZ (LacZ), Ad-apoE2 (E2), Ad-apoE3 (E3), and Ad-apoE4 (E4). After 48 h, the medium was replaced with serum-free medium, and a further 24 h later, the medium and cellular components were collected. A–C) Western blots for cellular apolipoproteins (A) and the bands for apoM and apoE were quantified with the ImageJ software, using β-actin as the internal standard (B, C) (n = 6). D, E) Real-time PCR for apoM and apoE mRNA (n = 12) using GAPDH used as the internal standard. F–J) The cellular contents of the sphingolipids were adjusted to the protein levels (n = 6). *p < 0.01 versus LacZ, †p < 0.05 versus LacZ. The p values for the differences among E2, E3, and E4 are shown in the figure panels. The horizontal bars represent the means of independently analyzed samples.
Overexpression of apoE3 in the U251 cells decreased, while overexpression of apoE4 increased the levels of ceramides in the medium
We next investigated modulation of the levels of apoM and sphingolipids in the culture media of the apoE-overexpressing U251 cells. There were no significant differences in the medium levels of apoE among the E2-, E3-, and E4-U251 cells (Fig. 4A, C). Overexpression of apoE decreased the medium levels of apoM (Fig. 4A, B). No effects of apoE overexpression were observed on the medium levels of S1P and dhS1P (Fig. 4D). The levels of Sph were higher in the E4-U251 cells than the LacZ-U251 cells and lower in the E3-U251 cells than the E2- and E4-U251 cells, while those of dhSph were lower in the E2-U251 cells than the E4-U251 cells (Fig. 4E). The effects on the medium levels of ceramides were opposite between the E3- or E2-U251 cells and the E4-U251 cells. The levels of C16, C18 : 0, C20, C22, and C24 ceramides were lower in the medium of the E3-U251 cells, but higher in that of the E4-U251 cells (Fig. 4F, H). In the E4-U251 cells, the levels of C18 : 1 ceramide were also higher (Fig. 4G). Although no significant changes in the levels of ceramides were observed in the medium of the E2-U251 cells, the overexpression of ApoE2 tended to decrease the levels of C16, C18, C22, and C24 ceramides. The medium ceramide-lowering effects of apoE were also in the order of apoE3 > apoE2 and apoE4 possesses the medium ceramide-increasing effects.
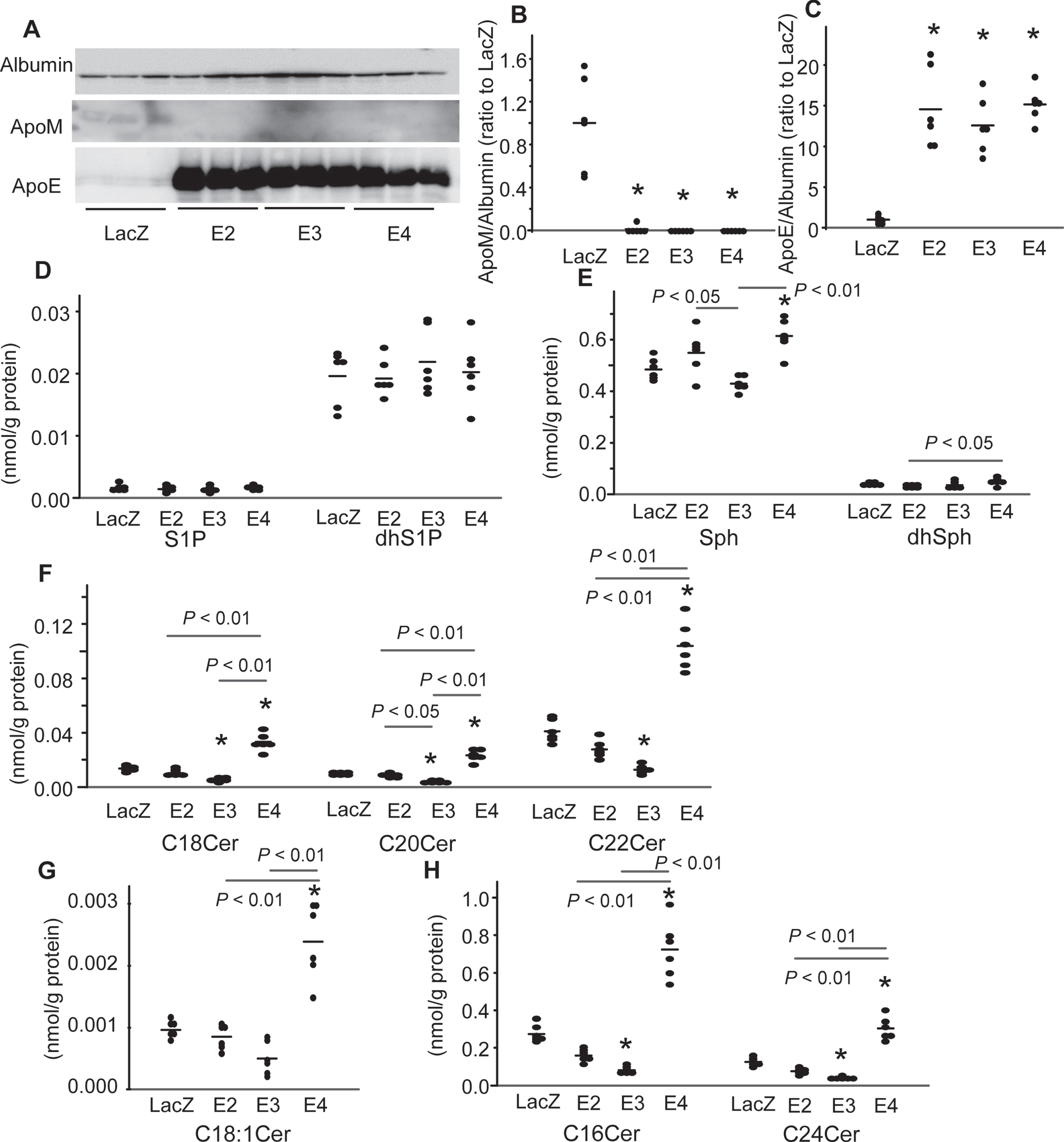
Modulation of the culture medium apoM and sphingolipid levels by overexpression of apoE isoforms in the U251 cells. The medium of the U251 cells described in Fig. 3 was analyzed for apolipoproteins and sphingolipids. A–C) Western blots for medium apolipoproteins (A) and the bands for apoM and apoE were quantified with the ImageJ software, using albumin as the internal standard (B, C) (n = 6). D–H) The medium levels of the sphingolipids were adjusted to the protein levels (n = 6). *p < 0.01 versus LacZ, †p < 0.05 versus LacZ. The p values for the differences among E2, E3, and E4 are shown in the figure panels. The horizontal bars represent the means of independently analyzed samples.
Overexpression of apoE in the SH-SY5Y cells modulated the cellular contents of apoM and sphingolipids, depending on the apoE isoform overexpressed
Next, we infected SH-SY5Y cells with Ad-LacZ (LacZ- SH-SY5Y cells), Ad-apoE2 (E2- SH-SY5Y cells), Ad-apoE3 (E3- SH-SY5Y cells), and Ad-apoE4 (E4- SH-SY5Y cells). The apoE protein and mRNA levels were not significantly different among the E2-, E3-, and E4-SH-SY5Y cells (Fig. 5A, C, E). The overexpression of apoE decreased the expression levels of apoM at both the cellular protein and mRNA levels (Fig. 3A, B, D). The mRNA levels of apoM in the SH-SY5Y cells were the highest in the cells overexpressing apoE2 among the apoE isoforms (Fig. 5D). The apoM and albumin were not detected by the western blot analyses in the medium of the SH-SY5Y cells.
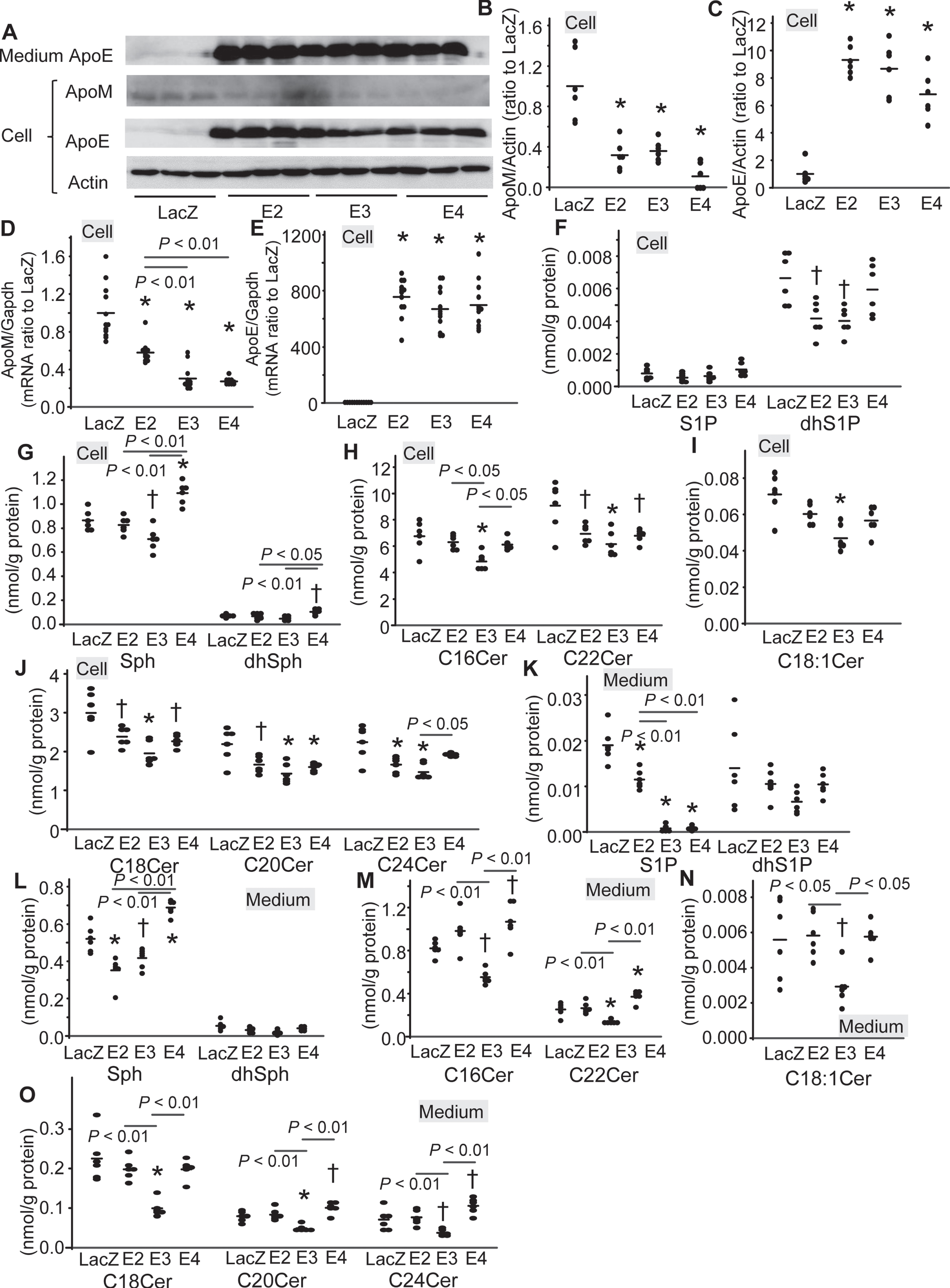
Modulation of the levels of apoM and sphingolipids by overexpression of apoE isoforms in the SH-SY5Y cells. SH-SY5Y cells were infected with Ad-LacZ (LacZ), Ad-apoE2 (E2), Ad-apoE3 (E3), and Ad-apoE4 (E4). After 48 h, the medium was replaced with serum-free medium, and a further 24 h later, the medium and cellular components were collected. A–C) Western blots for cellular and medium apolipoproteins (A) and the bands for apoM and apoE were quantified with the ImageJ software, using β-actin as the internal standard (B, C) (n = 6). D, E) Real-time PCR for apoM and apoE mRNA (n = 12) using GAPDH used as the internal standard. F–J) The cellular contents of the sphingolipids were adjusted to the protein levels (n = 6). K-O) The medium levels of the sphingolipids were adjusted to the protein levels (n = 6). *p < 0.01 versus LacZ, †p < 0.05 versus LacZ. The p values for the differences among E2, E3, and E4 are shown in the figure panels. The horizontal bars represent the means of independently analyzed samples.
Regarding the modulation of cellular S1P, dhS1P, Sph, and dhSph levels, the levels of dhS1P were lower in the ApoE2-SH-SY5Y cells, those of dhS1P and Sph were lower in the ApoE3-SH-SY5Y cells, and those of Sph and dhSph were higher in the ApoE4-SH-SY5Y cells (Fig. 5F, G). The levels of cellular ceramides were decreased by the overexpression of apoE, while the effects seemed highest in apoE3 (Fig. 5H-J). In the culture medium, the overexpression of apoE decreased the medium S1P levels, while the S1P levels were the highest in the E2-SH-SY5Y cells (Fig. 5K). The medium levels of Sph were lower in the apoE2-SH-SY5Y cells and those of Sph and C16, C18, C18 : 1, C20, C22, C24 ceramides were lower in the apoE3-SH-SY5Y cells, while those of Sph, C16, C20, C22, and C24 ceramides were higher in the apoE4-SH-SY-5Y cells (Fig. 5L-O).
Knockdown of apoE in the U251 cells increased the cellular and medium levels of apoM and sphingolipids
Although the modulation of Sph and ceramides by the overexpression of apoE3 was opposite to that in the brain of the Apoeshl mice, the modulation of apoM was not opposite to that in the brain of the Apoeshl mice; both the knockout and overexpression of apoE decreased the apoM levels. Therefore, we next investigated the effects of knockdown of apoE with siRNA on the levels of apoM and sphingolipids in the U251 cells. As shown in Fig. 6A-E, the knockdown of apoE increased the apoM medium protein, cellular protein, and mRNA levels. Concordantly, the cellular S1P levels were higher in the apoE-knockdown U251 cells (Fig. 6F). Regarding the other sphingolipids, the knockdown of apoE increased the cellular levels of Sph and C16, C18 : 0, C20, C22, and C24 ceramides (Fig. 6G-J). Although the knockdown of apoE did not significantly modulate the medium levels of S1P and Sph (Fig. 6K, L), it increased the medium levels of C16, C18 : 0, C20, C22, and C24 ceramides (Fig. 6M-O). These results suggested that apoE3, wild type apoE, possesses the lowering effects on apoM and sphingolipids in the U251 cells.
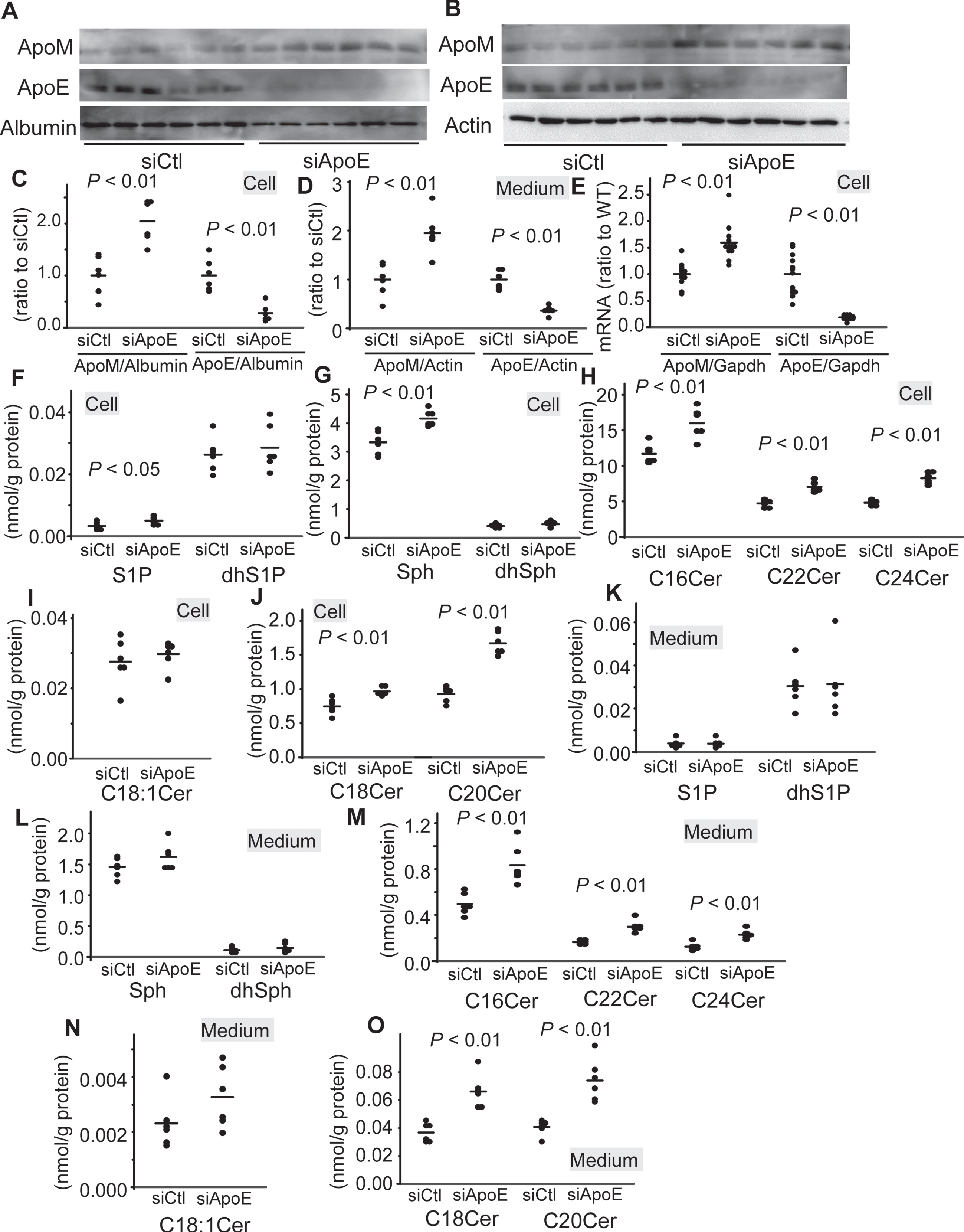
Modulation of the levels of apoM and sphingolipids by knockdown of apoE in the U251 cells. U251 cells were administered with siRNA against apoE (siApoE) or with control siRNA (siCtl). After 48 h, the medium was replaced with serum-free medium, and a further 24 h later, the medium and cellular components were collected. A–D) Western blots for medium and cellular apolipoproteins (A, B) and the bands for apoM and apoE were quantified with the ImageJ software, using β-actin as the internal standard (C, D) (n = 6). E) Real-time PCR for apoM and apoE mRNA (n = 12), using GAPDH used as the internal standard. F–J) The cellular contents of the sphingolipids were adjusted to the protein levels (n = 6). K-O) The medium levels of the sphingolipids were adjusted to the protein levels (n = 6). The horizontal bars represent the means of independently analyzed samples.
ApoE might differently modulate the nuclear proteins, which reportedly regulate apoM, between the U251 cells and the murine brains
So far, we have demonstrated wild type apoE (apoE3) negatively regulated the sphingosine and ceramide levels; however, the mechanisms for the discrepancies in the effects on apoM between the U251 cells or the SH-SY5Y cells and the murine brains remained unclear. To investigate this issue, we investigated the modulation of the nuclear proteins, which reportedly regulate apoM. RXRα, LXRα, PPARβ, and PPARδ negatively regulate apoM levels [41–43] and HNF-4α and SIRT1 positively regulate apoM [13, 44]. PPARγ also regulates the expression of apoM in a bell-shaped manner, with the greatest apoM levels observed when PPARγ was mildly activated in hepatocytes [45].
In the apoE-overexpressing U251 cells, the nuclear levels of RXRα and PPARδ were lower and those of HNF-4α were higher in the apoE2- and apoE4-U251 cells (Fig. 7A-C). Considering the modulation of apoM by nuclear proteins reported previously [43, 44], the modulation of these nuclear proteins was not responsible for the decreased expression of apoM. The knockdown of apoE decreased the nuclear levels of LXRα, PPARγ, and HNF-4α, while increased those of PPARδ (Fig. 7D-F). Among these nuclear proteins, the modulation of LXRα was concordant with the increased levels of apoM in the apoE-knockdown U251 cells. However, in the brains of the Apoeshl mice, the modulation of LXRα was not observed. Instead, the decreased levels of PPARβ, PPARγ, and SIRT1 were observed (Fig. 7G-I). Among them, the modulation of PPARγ and SIRT1 was concordant with the decreased levels of apoM in the brains of the Apoeshl mice, although the mRNA levels of apoM did not change.
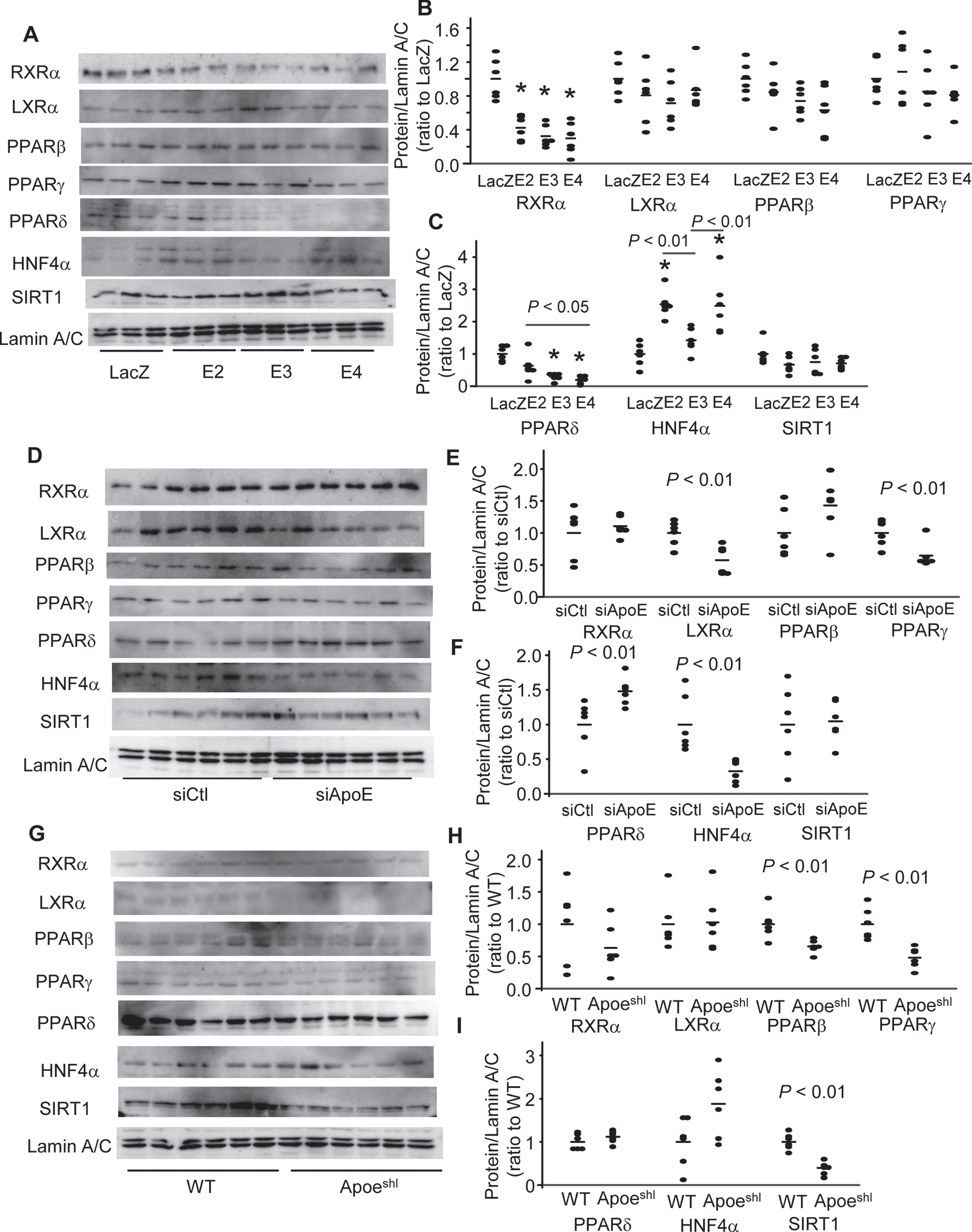
Modulation of the nuclear proteins in the U251 cells overexpressing apoE isoforms, the apoE-knockdown U251 cells, or the brains of Apoeshl mice. The nuclear proteins were prepared from the U251 cells overexpressing apoE isoforms (A-C), the apoE-knockdown U251 cells (D-F), or the brains of the Apoeshl mice as described in Fig. 3, Fig. 6, and Fig. 1 (n = 6). A, D, G) Western blots for nuclear proteins. B, C, E, F, H, I) the bands for nuclear proteins were quantified with the ImageJ software, using Lamin A/C as the internal standard (n = 6).
Overexpression of apoE2 and apoE4 decreased the cellular S1P and apoM levels in the HpG2 cells
Lastly, to compare with the results of the experiments using U251 cells and SH-SY5Y cells with those using other cell lines, we also infected HepG2 cells with Ad-LacZ (LacZ-HepG2 cells), Ad-apoE2 (E2-HepG2 cells), Ad-apoE3 (E3-HepG2 cells), and Ad-apoE4 (E4-HepG2 cells). The cellular apoE protein levels were higher in the E2- than the E3-HepG2 cells (Supplementary Figure 1A, C), while the cellular apoE mRNA levels were similar between the E2- and E3-HepG2 cells (Supplementary Figure 1E). In regard to apoM, the cellular apoM protein levels were lower in the E2- and E4-, but not E3-HepG2 cells (Supplementary Figure 1A, B), whereas the cellular apoM mRNA levels were lower in the E3- and E4-, but not E2-HepG2 cells (Supplementary Figure 1D). The cellular levels of S1P were lower in the E2- and E4-HepG2 cells, and those of Sph were lower in the E4-HepG2 cells (Fig. 5F, G). No significant changes were observed in the cellular expression levels of dhS1P, dhSph, or ceramides in any of the E2-, E3-, or E4-HepG2 cells (Supplementary Figure 1F–H).
Overexpression of apoE2 and apoE3 decreased the levels of S1P in the culture medium while that of apoE2, apoE3, or apoE4 decreased the levels of apoM in the culture medium of the HpG2 cells
The apoE levels were not different among the E2-, E3-, and E4-HpG2 cells (Supplementary Figure 2A, C). The apoM levels were lower in the HepG2 cells overexpressing any isoform types of apoE (Supplementary Figure 2A, B). On the other hand, the S1P levels were lower in the medium of the E2- and E3-, but not in that of the E4-HpG2 cells (Fig. 6D). No significant changes were observed in the levels of dhS1P, Sph, dhSph, or ceramides in the culture media of any of the E2-, E3-, or E4-HpG2 cells (Fig. 6D–F).
DISCUSSION
In the present study, we hypothesized that the modulations of apoM and sphingolipids might be among the mechanisms underlying the involvement of apoE isoforms in the development of Alzheimer’s disease and investigated the modulation of apoM and sphingolipids by apoE and its isoforms. We found that apoE negatively regulates the expression of apoM in the U251 cells and SH-SY5Y cells; however, the brain levels of apoM were lower in the Apoeshl mice. apoE3, wild type apoE, suppressed the cellular and medium levels of ceramides and the levels of ceramides were higher in the brains of the Apoeshl mice. Regarding the difference among apoE isoforms, the apoM and S1P levels were the highest in the cells overexpressing apoE2 and the lowering effects on the ceramide and sphingosine levels were the highest in the cells overexpressing apoE3, while the ceramide and sphingosine levels increased, especially in the medium, in the cells overexpressing apoE4. These modulation of apoM and sphingolipids might be unique to the CNS, since the similar modulation was not observed in the HepG2 cells.
In regard to the influences of the apoE on the metabolism of apoM and S1P in the brains of Apoeshl mice, which spontaneously lack apoE, the protein levels of apoM were lower and the levels of Sph, dhSph, and ceramides were higher in the brains of the Apoeshl mice (Fig. 2). Considering that apoE4 has relatively low influence on lipid metabolism as compared to apoE2 or apoE3 [23], the apoM levels in subjects with the apoE4 allele might most resemble those in the Apoeshl mice. On the contrary, both the cellular and culture medium levels of apoM protein were lower in the apoE-overexpressing U251 cells and SH-SY5Y cells (Fig. 3A, 3B, 4A, 4B, 5A, and 5B) and were higher in the apoE-knockdown U251 cells (Fig. 6A-D), suggesting that apoE might negatively regulate apoM in vitro. The regulation of apoM by apoE does not seem too simplistic though, since the brain apoM mRNA levels remained unchanged in the Apoeshl mice, whereas the apoE-overexpressing U251 cells and SH-SY5Y cells showed reduced levels of apoM mRNA (Fig. 2C, 3D, and 5D) and the apoE-knockdown U251 cells increased them (Fig. 6E). Until date, there have been no reports about the modulation of apoM by apoE. In regard to the regulation at the mRNA level, considering that apoE overexpression could modulate the lipid homeostasis [46] and that apoM has been reported to be regulated by several nuclear factors, such as PPARβ/δ [43], PPARγ [45], RXR, LXR [42, 47], HNF-4α [44], and SIRT1 [13], the activities of which can be modulated by lipid agonists, the modulation of the lipid contents might be at least partly involved in the regulation. In the present study, we observed that the overexpression of apoE decreased the nuclear levels of RXRα and PPARδ in the apoE-overexpressing U251 cells and increased those of HNF-4α in the apoE2- and apoE4-U251 cells and the knockdown of apoE decreased the nuclear levels of LXRα, PPARγ, and HNF-4α and increased those of PPARδ (Fig. 7A-F). Considering the previous reports, the modulation of LXRα was concordant with the increased levels of apoM in the apoE-knockdown U251 cells; however, the modulation of any nuclear proteins could not explain the decreased expression of apoM in the apoE-overexpressing U251 cells. Although the mRNA levels of apoM did not change, the decreased nuclear levels of PPARγ and SIRT1 were concordant with the decreased levels of apoM in the brains of the Apoeshl mice (Fig. 7G-I). In conclusion, these results could not clearly explain the modulation of apoM in the CNS; however, the modulation of the investigated nuclear proteins might be quite different between U251 cells and the murine brains, suggesting the possible existence of some unknown factors which differently regulate apoM levels between in vitro and in vivo experiments. Further studies are necessary to investigate the regulation of apoM in the CNS, as well as the involvement of apoM in CNS disorders.
In any case, the cellular protein or mRNA levels of apoM seemed to be the highest for apoE2 among apoE2, apoE3, and apoE4 in U251 cells or SH-SY5Y cells. The influences on apoM might represent one of the protective effects of apoE2, considering that apoM possesses beneficial properties and that apoE2 protects against Alzheimer’s disease. The modulation of S1P observed in the brains of the Apoeshl mice in this study (Fig. 2D) was not concordant with that of apoM, which was not consistent with the concept that apoM regulates the metabolism of S1P [37], while the modulation of S1P in the cellular levels of S1P in U251 cells was concordant with this concept (Fig. 3F and 6F). These results might reflect the fact that the levels of S1P were quite different between in vivo and in vitro. However, since apoM has been reported to determine not only the quantity of S1P, but also the properties of S1P [21], the effects of the apoE isoforms might explain, at least in part, the mechanisms underlying the involvement of apoE2 in the development of Alzheimer’s disease.
In regard to the modulation of sphingosine and ceramide levels, the levels of Sph, dhSph, C16, C18, C20, C22, and C24 ceramides were higher in the brains of the Apoeshl mice (Fig. 2E-H). Up to the present, only the levels of C16 : 0 ceramide have been reported to be lower in the apoE- knockout mice [33], a report that is not concordant with the observations in the present study. The reasons for these discrepancies might be that the experimental conditions were different: the present study investigated the modulation of ceramides in 10-week-old male Apoeshl mice, which still possess a small amount of apoE mRNA in the brain (Fig. 2C), while the other study conducted the investigation in 12-month-old female genetically apoE-deficient mice [33]. Since the deficiency of apoE may be expected to affect many pathophysiological processes in the brain [29], we believe that the lower levels of apoE per se would be associated with accumulation of ceramides in the brain. Actually, the plasma ceramide levels were higher in Apoeshl mice (Fig. 1E-G), which is consistent with the observations reported in genetically apoE-deficient mice [48]. Overexpression of apoE3, wild type apoE, in the U251 cells and SH-SY5Y cells also decreased the cellular sphingosine and ceramide levels, which might be reasonable since the results were opposite to those observed in the brain of the Apoeshl mice and the apoE-knockdown U251 cells. In regard to the ceramide levels in the culture media, overexpression of apoE3 decreased the medium ceramide levels, whereas that of apoE4 increased them (Fig. 4F-H, 5M-O). Considering that apoE4 is an established risk factor for Alzheimer’s disease and that ceramides have cytotoxic properties, the results might be concordant with the clinical phenotypes of apoE4. Although the mechanisms underlying the different effects of apoE isoforms on ceramides still remain unknown, the isoforms of apoE reportedly affect cholesterol metabolism in the CNS, and a recent study also demonstrated that apoE4 disturbs the homeostasis of non-cholesterol lipids in the CNS [49]. Overall, the modulation of ceramide metabolism by apoE is consistent with the clinical phenotypes, more for the case of apoE4 than for that of apoE3, since the medium and cellular ceramide levels were higher in the E4-U251 cells and the E4-SH-SY5Y cells, whereas the results were inconsistent when the effects of apoE2 and apoE3 were compared, since the intracellular ceramide-lowering effects were weaker and the medium ceramide levels were lower in the E3-U251 cells and the E3-SH-SY5Y cells, compared with E2-U251 cells and E2-SH-SY5Y cells.
Interestingly, the modulation of apoM and sphingolipids in the apoE-overexpressing HepG2 cells was quite different from that in the apoE-overexpressing U251 cells. First, the cellular apoM and S1P levels were lower in the E2- and E4-, but not E3-HpG2 cells, and the medium apoM and S1P levels were lower in the E2- and E3-HpG2 cells and the medium apoM levels were lower in the E4-HpG2 cells (Supplementary Figure 1A–F, Supplementary Figure 2A–D). The results described in the previous article were replicated in the E3-HpG2 cells, while the suppression of apoM mRNA levels was a novel finding, which might be explained in a manner similar to the aforementioned discussion on the U251 cells. The modulation of apoM by apoE overexpression might have been different between the E2- and E4-HpG2 cells, even though the cellular apoE levels were apparently the same; the apoM protein and mRNA levels were decreased in the E4-HpG2 cells, while only the apoM protein levels were decreased in the E2-HpG2 cells. The differential effects of the apoE isoforms observed in hepatic lipid homeostasis are also suspected to be applicable to the case of the brain, and further investigations are required, considering the important roles of apoM and apoE in circulation [21, 50]. In any case, the modulations of apoM and ceramides observed in the murine brains, U251 cells, and SH-SY5Y cells might be specific to the CNS.
One of the major limitations of the present study is that we did not elucidate in detail the mechanisms underlying the associations of the apoE isoforms with sphingolipid homeostasis; however, the present study is the first to propose that apoE isoforms might be closely involved in the disturbance of sphingolipid metabolism in both the circulation and the CNS in a different manner. This novel association might be partly involved in the pathogenesis of Alzheimer’s disease.
In summary, the levels of apoM were lower in the brains of Apoeshl mice, while overexpression of apoE decreased both the cellular and medium apoM levels in the order of apoE3 = apoE4 > apoE2, which might partly explain the protective effects of apoE2 against Alzheimer’s disease. The contents of ceramides were higher in the brains of the Apoeshl mice and the apoE-knockdown U251 cells, while overexpression of apoE2 and apoE3 decreased the cellular and medium levels of ceramides in the U251 cells and SH-SY5Y cells, in the order of apoE3 > apoE2, and overexpression of apoE4 increased them. Such modulations in the levels of ceramides by the apoE isoforms might represent one of the mechanisms underlying the linkage between apoE4 and Alzheimer’s disease.