
Abstract
Background:
40 Hz light flicker is a well-known non-invasive treatment that is thought to be effective in treating Alzheimer’s disease. However, the effects of 40 Hz visual stimulation on neural networks, synaptic plasticity, and learning and memory in wild-type animals remain unclear.
Objective:
We aimed to explore the impact of 40 Hz visual stimulation on synaptic plasticity, place cell, and learning and memory in wild-type mice.
Methods:
c-Fos+ cell distribution and in vivo electrophysiology was used to explore the effects of 40 Hz chronic visual stimulation on neural networks and neuroplasticity in wild-type mice. The character of c-Fos+ distribution in the brain and the changes of corticosterone levels in the blood were used to investigate the state of animal. Place cell analysis and novel location test were utilized to examine the effects of 40 Hz chronic visual stimulation on learning and memory in wild-type mice.
Results:
We found that 40 Hz light flicker significantly affected many brain regions that are related to stress. Also, 40 Hz induced gamma enrichment within 15 min after light flickers and impaired the expression of long-term potentiation (LTP), while facilitated the expression of long-term depression (LTD) in the hippocampal CA1. Furthermore, 40 Hz light flicker enhanced the expression of corticosterone, rendered well-formed place cells unstable and improved animal’s learning and memory in novel local recognition test, which could be blocked by pre-treatment with the LTD specific blocker Glu2A-3Y.
Conclusion:
These finding suggested that 40 Hz chronic light flicker contains stress effects, promoting learning and memory in wild-type mice via LTD.
INTRODUCTION
Alzheimer’s disease (AD) with progressive memory loss is the most prevalent type of dementia worldwide. AD patients are generally diagnosed by their presence of amyloid plaques and neurofibrillary tangles deposition. Recent animal studies suggest that 40 Hz visual or acoustic stimulation, specifically, could reduce amyloid-β in AD mice [1–3]. Several pilot studies have also reported that 40 Hz sound or light flicker exposure improves behavioral performance in 27 individuals [4, 5], showing that 40 Hz acoustic treatment improves mood, memory, and cognition scores in 9 individuals. Similarly, 40 Hz sensor stimulation twice a week over 6 weeks for a total of 6 times per treatment significantly impacted St. Louis University Mental Status Test scores in 18 AD patients. In addition, a feasibility trial in 10 prodromal AD patients showed that 4 to 8 weeks of 40 Hz flicker stimulation was safe and tolerable [6]. 40 Hz could induce gamma oscillations entrainment in many brain regions including visual, somatosensory, and prefrontal cortex, and hippocampal CA1 [1], suggesting visual sensor flicker could drive cognition-related hippocampal oscillation. However, whether 40 Hz flicker triggers brain remodeling, such as recruiting brain region inputs to the hippocampus, remains unclear.
The progression of memory deficit symptoms in AD is caused by amyloid-β accumulation and elevated tau levels in the brain [7, 8]. 40 Hz flicker not only improves learning and memory via the reduction of amyloid-β level in 5XFAD and APP/PS1 mice, but also decreases phosphorylated tau protein level in P301S tauopathy mice by microglia recruitment or astrocytes activation [2, 9–11]. Moreover, 40 Hz sensor stimulation modifies synaptic signaling and synaptic plasticity-related proteins in vivo [1]. Since synaptic plasticity is widely accepted as one of the major cellular mechanisms for learning and memory [12, 13], it is essential to investigate whether 40 Hz flicker remodels the expression of synaptic plasticity in wild-type mice.
Neural circuits and synaptic dysfunction lead to learning and memory deficits in AD [14, 15]. For better understanding and optimization of 40 Hz sensor stimulation treatment, it is essential to decipher the mechanisms of 40 Hz sensor stimulation on neural network and synaptic plasticity in wild-type mice.
MATERIALS AND METHODS
Animal
C57BL/6 mice were purchased from Beijing Weitong Lihua Experimental Animal Co., Ltd. Animals were housed at the animal facility of the Shenzhen Institute of Advanced Technology, Chinese Academy of Sciences in a 12-h light/dark cycle and temperature-controlled individually ventilated cages. Food and water were provided ad libitum. As shown in Figs. 1 and 2, 36 mice were treated with 40 Hz light stimulation, and 16 mice as control. 23 mice used in place cell recording, and 23 mice were used for behavior tests. All animal experiments were performed according to the protocol approved by the Institutional Animal Care and Use Committee of Shenzhen Institute of Advanced Technology, Chinese Academy of Sciences (approval number: SIAT-IACUC-210303-NS-YX-A1694).
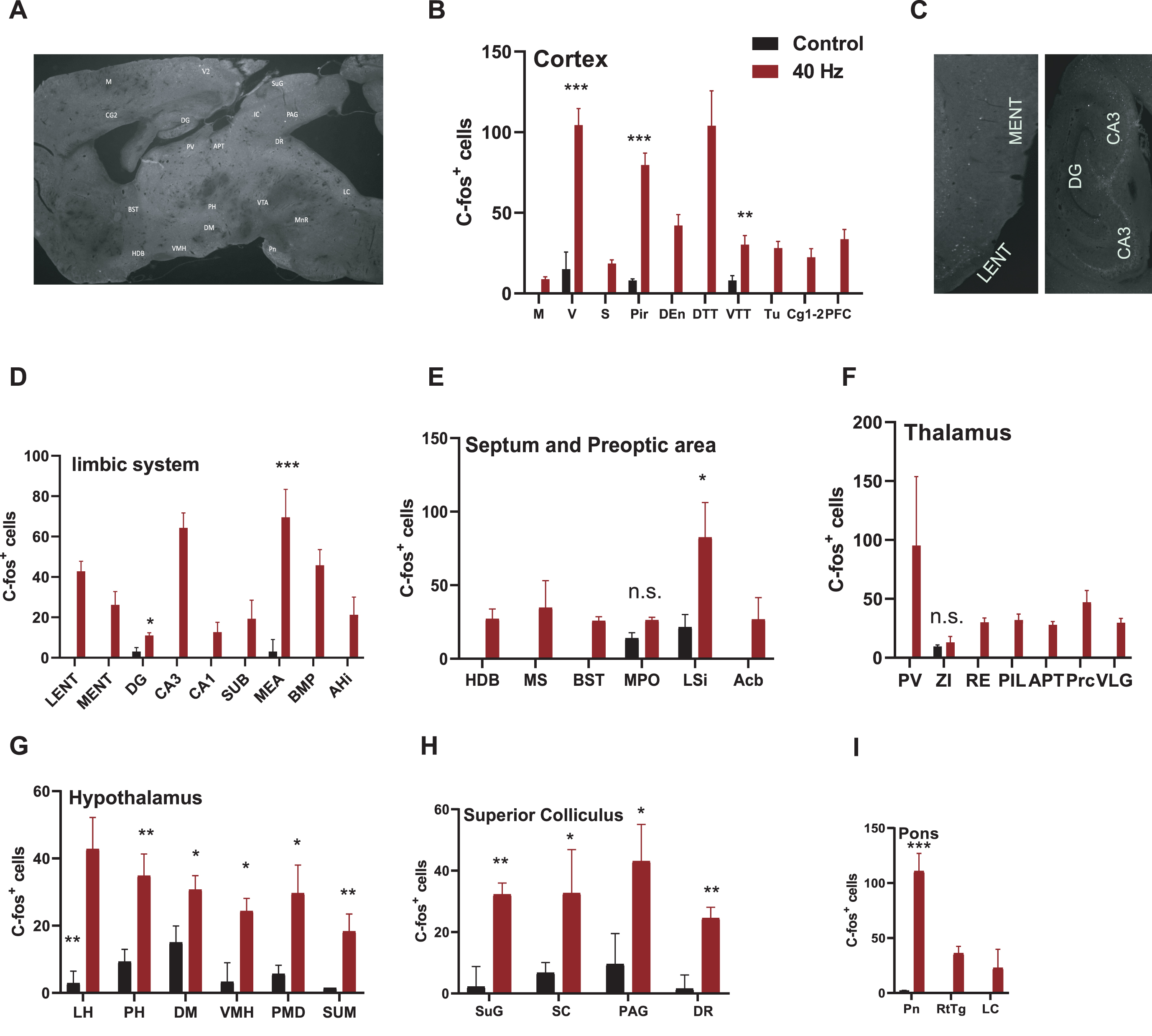
40 Hz light flicker affected many stress-related brain regions including hippocampal CA3. A) C57 mice received 40 Hz (N = 6) chronic visual stimulation for 1 h per day for 7 days. After 7 days, brain areas specifically activated by chronic 40 Hz visual stimulation were screened by c-Fos staining. Counting results showed the visual stimulation of 40 Hz (B–I). The brain regions with either dim or scattered signals were not taken in count and were shown as 0 in the graphs and were excluded from statistical analysis. Control versus 40 Hz, *p < 0.05, **p < 0.01, ***p < 0.001. n.s., no significant. Acb, accumbens nucleus; AHi, amygdalohippocampal area; APT, anterior pretectal nucleus; BMP, basomedial amygdaloid nucleus, posterior part; BST, bed nucleus of stria terminalis; CA1, field CA1 of hippocampus; CA3, field CA3 of hippocampus; CG1,2, cingulate cortex, area 1,2; DEn, dorsal endopiriformnucleus; DG, dentate gyrus; DM, dorsomedial hypothalamic nucleus; DR, dorsal raphe nucleus; DTT, dorsal tenia tecta; HDB, nucleus of the horizontal limb of the diagonal band; LC, locus coeruleus; LENT, Lateral entorhinal cortex; LH, lateral hypothalamicnucleus; LSi, lateral septal nucleus, intermediate part; M, motor cortex; MEA, medial amygdaloid nucleus, anterior part; MENT medial entorhinal cortex; MPO, medial preoptic nucleus; MS, medial septal nucleus; PAG, periaqueductal gray; PFC, prefrontal cortex; PH, posterior hypothalamic area; PIL, posterior intralaminar thalamic nucleus; Pir, piriform cortex; PMD premammillary nucleus, dorsal part; Pn, pontine nucleus; Prc, precommissural nucleus; PV, paraventricular thalamic nucleus; RE, reuniens thalamic nucleus; RtTg, reticulotegmentalnucleus of the pons; S, somatosensory cortex; SC, scaphoid thalamic nucleus; SUB, subiculum; SUG, superficial gray layer of the superior colliculus; SUM, supramammillary nucleus; Tu, tuberal nucleus; V, visual cortex; VLG, ventral lateral geniculate nucleus; VMH, ventromedial hypothalamic nucleus; VTT, ventral tenia tecta; ZI, zona incert.
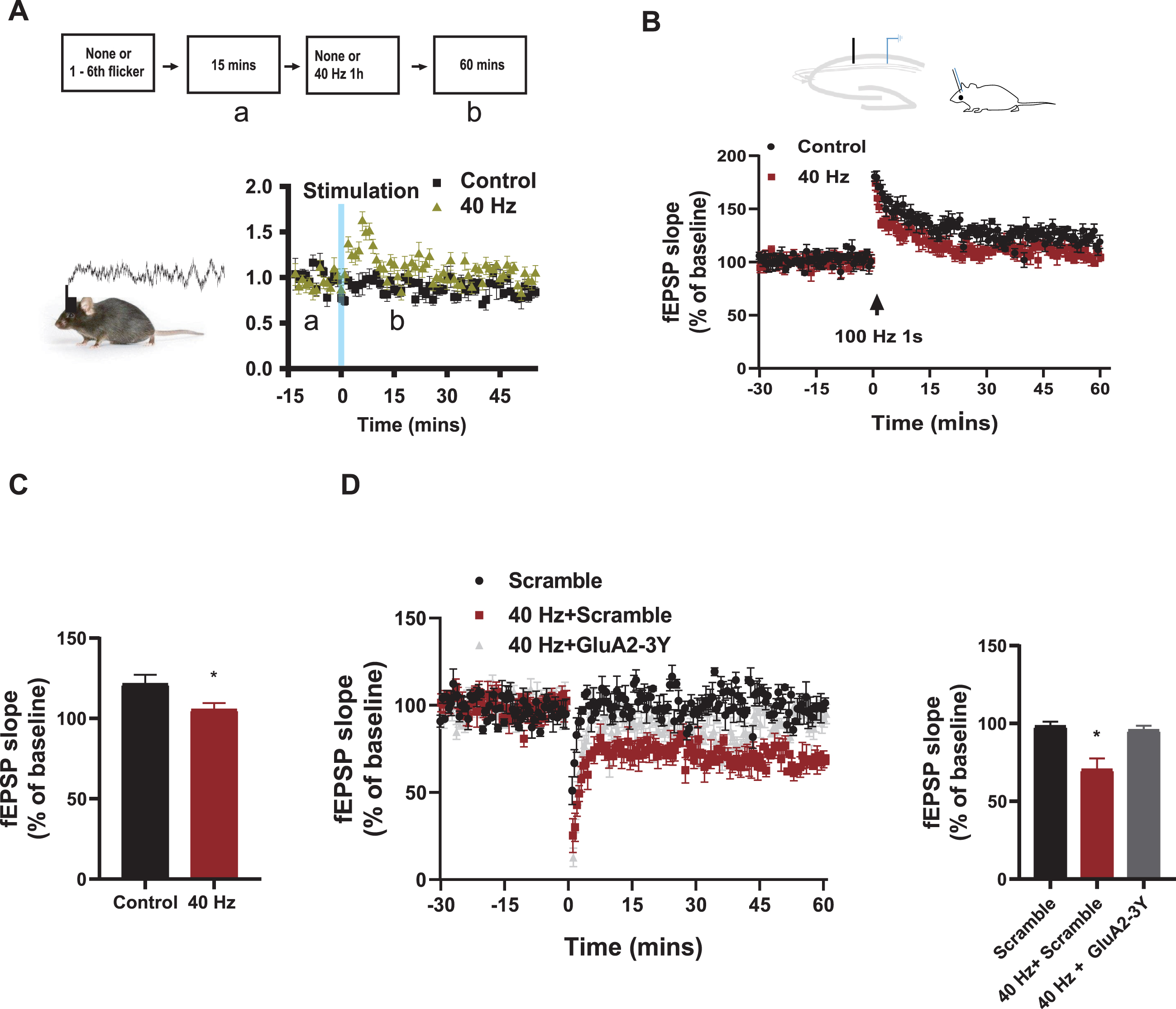
40 Hz light flicker remodeled hippocampal slow gamma oscillation and plasticity expression. A) Flow diagram of the experiment. Local field potential recorded in the CA1 area for 15 mins as baseline and 1 h after the 7th 40 Hz (N = 12) visual flicker, and then gamma oscillation isolated. Results showed that 40 Hz visual stimulation increased the power of gamma oscillations in the CA1 region (B). Chronic visual stimulation at 40 Hz (N = 8) in stimulated C57 mice inhibited LTP induction (C), compared to unstimulated controls (N = 5), Control versus 40 Hz, *p < 0.05. Chronic visual stimulation at 40 Hz (N = 5) in stimulated C57 mice promoted LTD expression (D), which were blocked by Tat-GluA2-3Y peptide pre-treatment in the 40 Hz + GluA2-3Y mice (N = 5) group, compared with mice treated with scramble (N = 4) and GluA2-3Y (N = 4) alone. Scramble versus 40 Hz + Scramble, 40 Hz + Scramble versus 40 Hz + GluA2-3Y, *p < 0.05.
Electrode complex implantation
Wild-type C57 mice were anesthetized with 2–5% isoflurane and fixed on a stereotaxic device (RWD, http://www.rwdls.com). After exposing the skull, a hole was drilled above the hippocampus for the preparation of the microinjection. Sixteen 17μm metal wires (California Fine Wire Company) were twisted into 2 strands and inserted into a homemade tiny driver to form an in vivo electrode complex with adjustable recording depth, which was then implanted over the hippocampal CA1. Approximately 2 g of the electrode complex was fixed to the mouse skull with dental cement and small screws. Support pain relief and anti-inflammatory treatment with 0.5 mg/kg meloxicam (Metacam) and 5 mg/kg Dexamethasone (Sigma). After completion of surgery, mice were returned to their home cages after their recovery from anesthesia.
LTP and LTD in vivo
After the animals were anesthetized with isoflurane, they were immobilized on a stereotactic device (RWD). According to the brain stereotaxic map, we inserted the stimulating electrode between CA3 and CA1 in the hippocampus (AP: –2.3 mm, ML: 2.5 mm, DV: 2.0) and placed the recording electrode at the end of CA1 (AP: –2.3 mm, ML: 1.35 mm, DV: 1.45 mm relative to bregma). The position of the stimulating and recording electrodes was determined by observing the typical postsynaptic responses recorded by the recording electrodes. The postsynaptic potential slope was recorded (A–M system). Fifteen minutes from baseline, we applied the protocols for inducing long term potentiation (LTP) and long term depression (LTD) (1 s 100 Hz and 1 Hz 900 pulses, respectively, software: WinLTP) to stimulate SC fibers through the stimulating electrode. The postsynaptic response was then recorded for 60 min. The expression of synaptic plasticity was evaluated by analyzing the slope change of the postsynaptic potential in the last 5 min and before stimulation. Tat-Glu2A-3Y or scrambled peptide systems were processed 30 min before baseline recording. Tat-GluA2-3Y with the sequence YGRKKRRQRRRYKEGYNVYG and its scrambled peptide with the sequence GRKKRRQRRVYKYGGYNE were synthesized by Tocris.
Place cell and neural oscillation
After mice were embedded with recording electrodes and recovered from surgery, we lowered the electrodes roughly to the CA1 region of the hippocampus and then lowered the electrode depth in 30μm steps while looking for firing of neurons in the CA1 region. At the same time, the firing of neurons in the CA1 area was recorded (Plexon) while the animal was moving freely in an open field. The recorded electrical signals were filtered by a 300 Hz low-pass filter to obtain the field electrical data. And then a 30–55 Hz band-pass filter was used to obtain slow gamma oscillations, and energy changes were obtained by current-voltage squared calculations. The firing signals of neurons were obtained by high-pass filtering, and the L-ratio (<0.2) or the distance between firing cells in the channel was greater than 20, which was considered as one unit firing. Place cell is a kind of cells that becomes activate when animal visiting a particular space in its environments. The place cells (firing rate maps) were analyzed with the commercial software NeuroExplorer 5 (Plexon) to obtain the firing map and the place field.
Visual flicker
We used a stimulator to trigger the light source (PSU-H-LED, QAXK-LASER-473, range from 0–120 mW) to generate 40 Hz light pulses. The 473±1 nm light came out of the source (the power out of port, 5.5–5.8 mW/mm2) with intensity 5 mW/mm2 on the tip of 200μm-diameter optical fiber (Ocean Optics). The animals were treated with the distance within ∼10 cm height for one hour per day in their home cages for 7 consecutive days.
Immunohistochemistry
Brain tissue was perfused with normal saline followed by 4% paraformaldehyde (PFA) solution, dehydrated with 30% sucrose and frozen sections to obtain 30μm sagittal sections with a cryostatmicrotome (Leica). After incubation with antibodies against c-Fos (Millipore, ABE457, 1:500), the brain slices were labelled with secondary antibodies (Alexa Fluor 488 Invitrogen™) to exhibit the expression of c-Fos in the brain. c-Fos+ cells were imaged under Zeiss Axio Zoom Macroscope. Only c-Fos+ cells that were apparent in the slices were counted. Signals(c-Fos+ cells) that were dim or scattered were not taken in count. The statistical analysis were only used for brain region with clear signals in control group mice.
New location recognition test
Animals were habituated in the behavioral room for 1 h before the training session. During the training, animals were exposed to a behavioral box (40×40 cm, RWD) containing two similar objects, each 5 cm from the edge of the box. Animals were allowed to freely access and familiarize with the two objects in the box for 20 s. Animals were returned to their home cages after the training.
The testing session was conducted 6 h after training. After randomly placing one of these two objects in a new position, animals were introduced to the behavioral box again. The time animal spent visiting the object in the new location was recorded, and the change in time were used for the assessment of its learning and memory.
Statistical analysis
Data are showed as mean±SEM. Parametric tests including Student t-test and two-way ANOVA were performed, and the statistical significance were defined as p < 0.05 in this research.
RESULTS
40 Hz light flicker increased c-Fos+ cells in many stress-related brain regions including the hippocampal CA3
To investigate the effects of 40 Hz visual stimulations in wild-type mice, we stained c-Fos after light flickers. 6-month-old wild-type mice received 1 h of 40 Hz (5 mW, 473 nm) visual stimulation for 7 days and were stained for c-Fos 90 min after the 7th stimulation. C-Fos positive cells were shown in 30μm contiguous sagittal sections spaced 0.2 mm from side to side (Fig. 1A). Similar to 40 Hz visual stimulation enrolled cells in V1, CA1, and prefrontal cortex by electrical recording [1], we found that compared to untreated control mice that did not receive the stimuli (N = 3), animals with visual stimuli at 40 Hz have significantly increased c-Fos-positive cells in the cortex (Fig. 1B), the limbic system (Fig. 1C, D), the septum and the preoptic area (Fig. 1E), the thalamus (Fig. 1F), the hypothalamus (Fig. 1G), the superior colliculus (Fig. 1H), and pons (Fig. 1I). The high level of c-Fos+ cells in the hippocampus and stress-related brain region in the hypothalamus [16] indicates that 40 Hz light flicker may relate to hippocampal function and stress/fear/anxiety.
40 Hz light flicker remodeled hippocampal slow gamma oscillation and plasticity expression
Considering the regional vulnerability of the hippocampus in mouse models of AD and human patients [17, 18], and experiences could alter neuroplasticity in the brain [19], especially in the hippocampus, we further tested the effect of chronic 40 Hz visual stimulation on hippocampal function. To avoid the complex effects of chronic stimulation and the baseline instability of prolonged recording in electrophysiological recordings, we compared the changes before and after the last 1 h of sensor stimulation. The electrical signal was recorded in CA1 for 15 min in 6-month-old mice before the 7th light stimulation, and then recorded for 1 h after the 7th visual stimulation (Fig. 2A). We found that compared to unstimulated mice, 40 Hz visual stimulation significantly enhanced slow gamma power 1.5 times within 15 min compared to the baseline and then decreased to baseline levels (p < 0.001, Fig. 2A), indicating that 40 Hz produced a prolonged effects in hippocampal function.
Next, we tested whether 40 Hz visual flicker induced plasticity changes in SC-CA1 synapses. After visual flicker treatment, metal stimulation and recording electrodes were implanted in the Schaeffer collateral and the CA1 of anesthetized mice. Classical protocols for inducing LTP at 100 Hz for 1 s and LTD at 1 Hz for 900 s were used. We showed that in contrast to the successful expression of LTP in control mice, where the slope of fEPSP reached 130% of baseline, LTP expressions were impaired in 40 Hz visual flicker-treated mice, and the slope of fEPSP after tetanic stimulation were comparable to baseline (Fig. 2B, C). However, the fEPSP slope in 40 Hz visual flicker-treated mice decreased to 75% of baseline, compared to control mice without LTD expression (Fig. 2D), suggesting that LTD induction is facilitated by those with chronic visual stimulation. To verify visual stimulus-induced LTD enhancement, the well-studied Tat-GluA2-3Y peptide was utilized as a specific blocker of LTD [20–22]. 3 mg/kg Scramble peptide or Tat-GluA2-3Y peptide was injected by systemic injection 30 min before stimulation. We found that Tat-GluA2-3Y peptide pre-treatment restored fEPSP levels to baseline in 40 Hz + GluA2-3Y mice (Fig. 2D), indicating that those promoting visual flicker-induced LTD expression were blocked by Tat-GluA2-3Y peptide pre-treatment.
40 Hz light flicker, acting as a stressor, remaps well-formed place cells in CA1 via LTD
40 Hz chronic visual flicker shift synaptic plasticity towards LTD in the hippocampus and increased the level of c-Fos+ cells in many brain regions including stress/fear/anxiety axis [16] and CA3, implying that an important component of a 7-day chronic visual flicker stimulation could act as a stressor. Therefore, we tested corticosterone secretion level after the final visual flicker and found that 40 Hz visual stimulation increased plasma corticosterone from a control level of 450.6±15 ng/ml (N = 4) to 600±70 ng/ml (n = 7; p < 0.05; Fig. 3A). Together, these data suggest that stress is a key component of 40 Hz visual flicker.
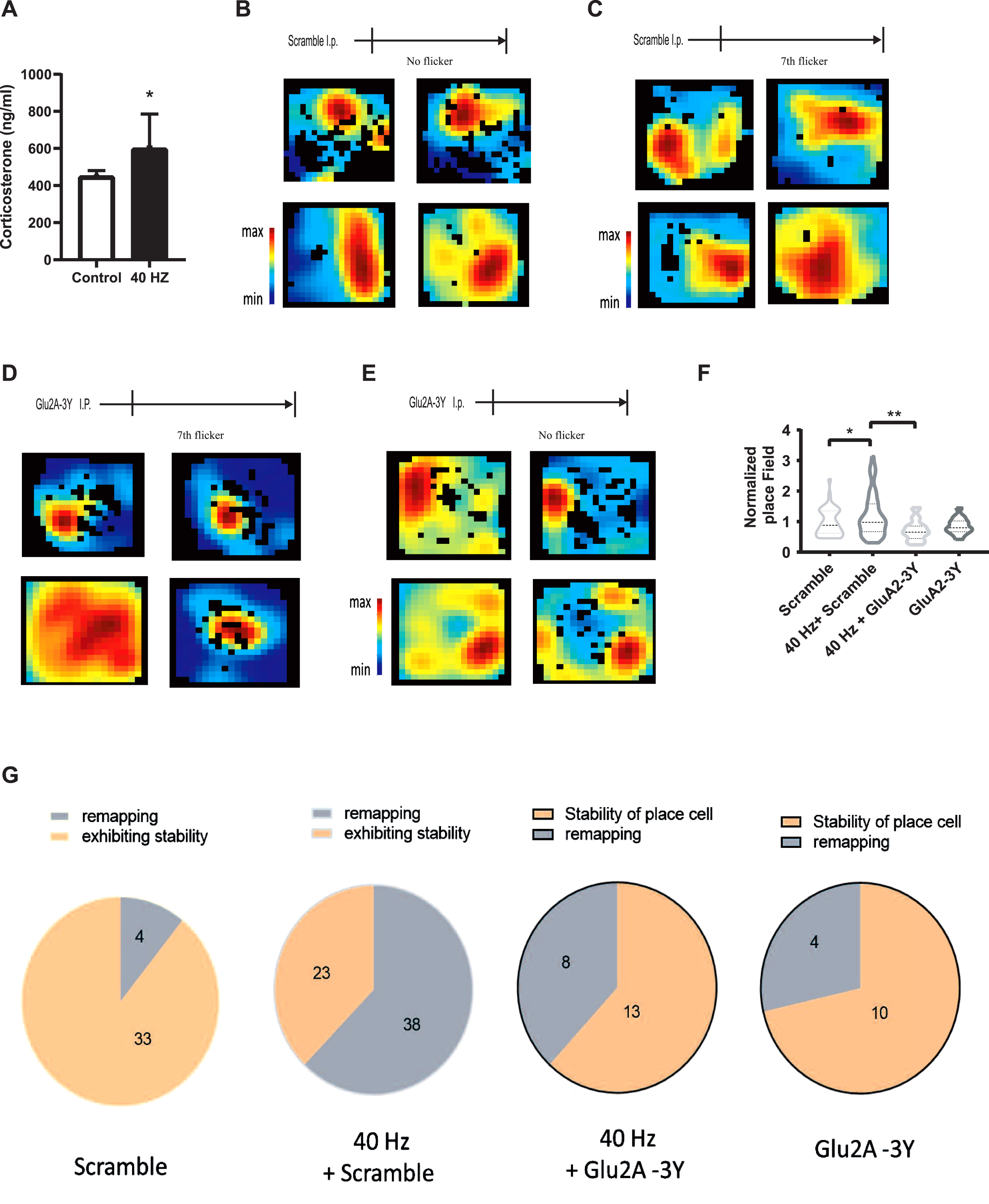
40 Hz light flicker, acting as a stressor, remaps well-formed place cells in CA1 via LTD. The secretion of corticosterone levels after 7 days with (N = 7) or without chronic light flickers (N = 4, A), Control versus 40 Hz, *p < 0.05. After well-formed place cells established during animal habituation within 3 days, mice underwent (C, D) or without (B, E) 6 days light stimulation, and followed by intraperitoneal injection with 3 mg/kg Tat-GluA2-3Y peptide (N = 12, D, E) or the same dose of scramble peptide (N = 11, B, C) 30 min before the last visual stimulation. Cell firing was recorded within 10 min after the habituation and the last visual flicker. F) Statistics for place field of each group, Scramble versus 40 Hz + Scramble, *p < 0.05, 40 Hz + Scramble versus 40 Hz + GluA2-3Y, p < 0.01. G) Pie graph showing the stability of place cells in CA1 in each group.
Place cells are directly modulated by LTP and LTD [23, 24] and the surrounding factors, including fear [24, 25]. Thus, we further investigated the effects of 40 Hz visual flicker on place cells before and after 40 Hz stimulation. The well-formed place cells were recorded in the mice hippocampal CA1 when animals were exploring freely in a box for 10 min after mice habituated in open box for 3–5 days, cell firing was recorded in the open box after seven days with or without chronic visual stimulation (Fig. 3C, D). We showed that compared with place cells fired in similar place without visual stimulation (33/37, Fig. 3B, E), 7 days 40 Hz visual stimulation induced many place cells re-localized into another space (38/61, Fig. 3C, G). However, 3 mg/kg Tat-Glu2A-3Y peptide pre-treatment blocked those remapping (Fig. 3D, G) in mice pretreated with Tat-Glu2A-3Y peptide and 40 Hz treatment (Fig. 3E, G), indicating that chronic 40 Hz visual stimulation destabilized place cells.
Place cells fire in a specific space of an environment, known as a place field. Since the stability of place field indicates the accuracy of place information. We further analyzed the changes of place field in each group. We found that the place field was significant increased by 40 Hz stimulation treatment (p < 0.05), whereas this enhancement was blocked by Glu2A-3Y peptide pre-treatment (p < 0.01; Fig. 3F). Thus, these data indicated that 40 Hz light stimulation unstabilized the well-formed place cells via LTD.
40 Hz light flicker enhances learning and memory in wild-type mice
Place cells mechanistic guide spatial learning and memory in animals and have been implicated in AD [26, 27]. The potential effects on neural plasticity and place cells implied that 40 Hz stimulation generally facilitates learning and memory in wild-type mice. We tested the effects of 40 Hz light flicker on spatial learning and memory in wild-type C57 mice by a weak learning and memory protocol: one trail novel location recognition. Mice were trained to recognize objects in a box during a training session, and then, tested in the probe test with the location of the object changed to a new place 6 h after the training. The time the animal spent on the new location was used to assess the strength of learning and memory (memory test, Fig. 4A). The membrane-permeable Glu2A-derived Tat-Glu2A-3Y peptide and a control scrambled peptide with an intact Tat domain but scrambled GluR2A-3Y sequence were used to test whether 40 Hz light flicker affects LTD. We found that mice received 40 Hz light flicker spent significantly more time in the probe test than control mice that received scrambled or Tat-Glu2A-3Y peptide pre-treatment alone. Also, 40 Hz enhanced animal performance could be blocked by Tat-Glu2A-3Y peptide pre-treatment in 40 Hz + Glu2A-3Y group mice (Fig. 4B), regardless of comparable level of speed and time spent on the two objects in the training session for these four groups of mice (Fig. 4C, D). This indicated that 40 Hz light flicker enhanced learning and memory via LTD in wild-type mice.
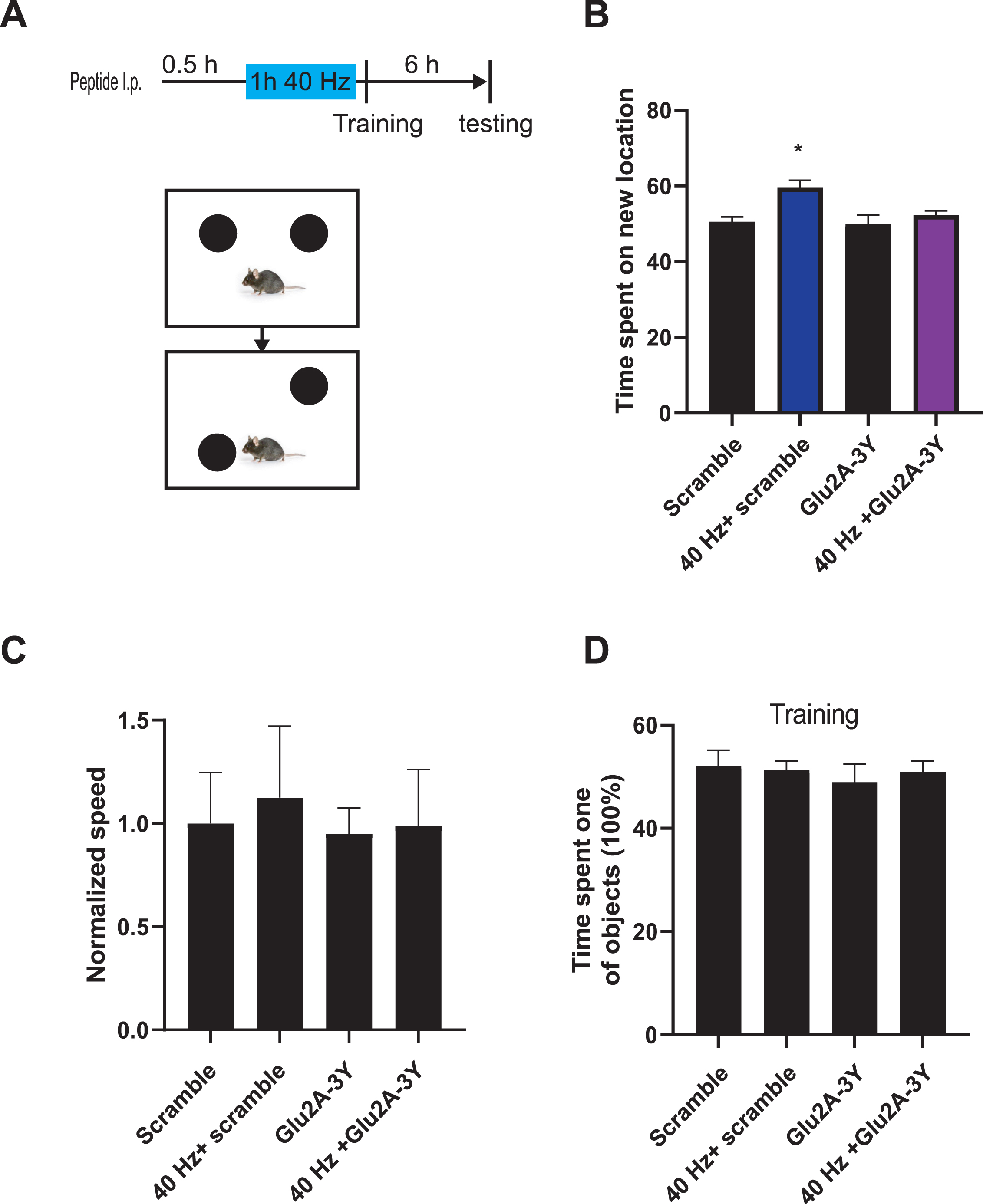
40 Hz chronic visual stimulation enhances the learning and memory in wild-type mice. A) Experimental flow chart. Intraperitoneal injection of wild-type mice with 3 mg/kg Tat-GluA2-3Y peptide (N = 11) or the same dose of scramble peptide (N = 12) 30 min before the last 40 Hz visual stimulation. Mice were then trained in one trail of new location recognition task, and the time the animal spent on new location of object was recorded in probe testing 6 h after training session. B) Compared with the C57 scramble group (N = 7) that only received the scramble peptide and the C57 mice treated with Tat-GluA2-3Y peptide (N = 6) alone, the 40 Hz treated mice coupled with scramble peptide (N = 5) showed significant improvement on behavior performance, and the pre-treatment of Tat-GluA2-3Y peptide (N = 5) could block those improvement induced by chronic visual stimulation in wild-type mice, Scramble versus 40 Hz + Scramble, 40 Hz + Scramble versus 40 Hz + GluA2-3Y, *p < 0.05. The speed (C) and time spent in two objects (D) in each group were comparable.
DISCUSSION
In the present study, we demonstrated that 7-days 40 Hz visual flicker contains stress effects, induces gamma enrichment, remodels synaptic plasticity, and promotes place cell remapping and learning and memory performance in wild-type mice. Taking into account that around 50% of the adverse reactions (including 25% dry eye,13% light sensitivity, 13% anxiety) occurred in chronic 40 Hz sensor stimulation treatment of AD patients and control participants [6, 28], this study points out the stress effects induced by 40 Hz in wild-type mice. These findings suggest that eliminating stress responses by dividing patients into stressed and non-stressed groups maybe essential in clinic trial. The stress level of the patient or animal taken into consideration should be more effectively to amplify or clarify the effects of chronic 40 Hz stimulation in disease.
For treated animals, the level of corticosterone are expected to increase for acute stress then decrease after repeats (but still higher than non-stressed ones) [29] or when the animals are away from the stressor [30]. However, the level of corticosterone stays in high level for chronic stress [31]. Unlike mouse models of neurological diseases, such as AD mice that originally come with high stress level [32], which stress induced by 40 Hz sensor treatment might been masked in disease model mice [3]. In our study, compared with non-treated groups, 40 Hz chronic light flicker acts as a stressor in wild-type mice. 40 Hz chronic light flicker enhances the c-Fos + cells in hippocampal CA3 and hypothalamic LH, PH, and VMH, which are all closely related to stress. Moreover, the c-Fos+ cell increased in CA3 indicating hippocampal functional remodeling was further validated by the changes in slow gamma oscillation and synaptic plasticity induced by 40 Hz flicker. Consistent with previous studies showing that stress impairs LTP but promotes LTD expression in the hippocampus [33–35], 40 Hz chronic light flicker also inhibited LTP expression and improved LTD expression in the hippocampal CA1. Since LTP stabilizes place cells [23] and forgetting is related to destabilizing well-formed place cells in the hippocampus [24, 36], these findings indicate that 40 Hz chronic light flicker destabilizes the CA1 well-formed place cell via LTP impairment and LTD facilitation. Furthermore, LTD is required for hippocampal dependent learning and memory [20–22] and stress, even chronic stress improves cognitive function [37]. For example, cold stress enhances animal spatial performance in Morris water maze [38]. In our study, wild-type animals without suffering from diseases treated with 40 Hz chronic light flicker shows stress effects, including activation of fear/anxiety related brain regions, the facilitation of LTD expression in hippocampal CA1, the destabilization of well-formed place cell, and the improvement of learning and memory performance.
Gamma oscillations (30–100 Hz) in the brain are relevant for neurocircuit function, behavior, and memory [39]. And the power of slow gamma (or theta locked power) is significant enhanced during LTP induction [40]. Consistent with benefits from 40 Hz in AD [1], it could be recruited by 40 Hz sensor stimulation and may link to neuroimmunology, effectively attenuating AD-related pathology [2]. However, synaptic plasticity dysfunction in AD is mainly LTP impairment in animals and patients [41, 42], and chronic TatGluA2-3Y treatment blocked LTD showed effectively intervention on memory loss in AD [43]. Our study showed that 40 Hz promoted LTD (10 Hz also did, data not shown) and learning and memory improvement may not be a good candidate mechanism to demonstrate potential ability of 40 Hz oscillation in AD therapy, but this mechanism may enhance the animals’ sensitivity to incoming danger situations and new information coding, which is important to increase its probability of survival via memory enhancement [44, 45].
Limitations
We showed that 40 Hz chronic visual flicker affects many brain regions, including CA3, leading to hippocampal functional remodeling in wild-type mice. However, limited by the level of accuracy in c-Fos staining in the whole brain and general activation of SC fiber in our LTP and LTD study, we do not exclude that 40 Hz non-invasive stimulation may affect some different and special neural circuits, or different subgroup of cell or engram cells/special circuits dependent plasticity to make benefit on animal performance. However, we retain our opinion that 7-days 40 Hz visual flicker facilitates learning and memory via LTD in wild-type mice.
CONCLUSION
Neural networks and neural plasticity remodeling are key to understanding brain adaptation and brain functional changes to external stimuli, such as 40 Hz sensor stimulation. Here, we showed that 40 Hz chronic visual flicker induces stress-related neural networks and hippocampal neuroplasticity remodeling in wild-type mice. Our study not only provides evidence suggesting that 40 Hz stimulation induces LTD facilitates memory, which could be a mechanism for animal survival, but also supports that compared to ordinary neuroplasticity, gamma power and neuroimmunity are more likely to be a mechanism for 40 Hz non-invasive sensory intervention in AD.