
Abstract
Background:
Recent studies have revealed that atrial fibrillation (AF) patients have a high risk of developing cognitive impairment, vascular dementia, and Alzheimer’s disease (AD). Some reports suggest that the application of oral anticoagulant with an appropriate dose may have a preventive effect on AD. However, which oral anticoagulant drug is more appropriate for preventing AD and the underlying mechanism(s) is still unknown.
Objective:
The aim of the present study was to assess the treatment effect of rivaroxaban administration as well as investigate the roles of PAR-1 and PAR-2 in the AD + CAA mice model.
Methods:
In the present study, we compared a traditional oral anticoagulant, warfarin, and a direct oral anticoagulant (DOAC), rivaroxaban, via long-term administration to an AD with cerebral amyloid angiopathy (CAA) mice model.
Results:
Rivaroxaban treatment attenuated neuroinflammation, blood-brain barrier dysfunction, memory deficits, and amyloid-β deposition through PAR-1/PAR-2 inhibition in the AD + CAA mice model compared with warfarin and no-treatment groups.
Conclusion:
The present study demonstrates that rivaroxaban can attenuate AD progress and can be a potential choice to prevent AD.
Keywords
INTRODUCTION
The prevalence of atrial fibrillation (AF) increases with age. AF is now regarded as a risk factor for cognitive impairment and dementia among the elderly patients [1, 2]. Alzheimer’s disease (AD) is the most common cause of dementia, involving multiple pathogenic factors and causing profound impairments in learning and memory functions [3, 4]. Cerebral amyloid angiopathy (CAA), which is one of the three pathological hallmarks of AD, is present in more than 80% of AD patients, is characterized by the accumulation of amyloid-β (Aβ) in cortical and leptomeningeal blood vessels, and can directly contribute to cognitive decline and dementia by causing vascular lesions [5 –8]. Oral anticoagulant, such as warfarin and rivaroxaban, showed beneficial effects for the prevention and treatment of stroke associated with AF [9, 10]. However, some reports indicated that the application of anticoagulant may increase the risk of intracerebral hemorrhages in patients with both CAA and atrial fibrillation [9 –12]. Moreover, warfarin had a higher risk of cognitive impairment than the direct oral anticoagulant (DOAC) [13, 14].
Warfarin has vitamin K antagonist function and causes an inhibition of the vitamin K-dependent coagulation factors II, VII, IX, and X [15 –17]. Rivaroxaban is a DOAC that directly inhibits FXa and reduces thrombin production. [18 –20]. FXa activates Protease-activated receptor-1 (PAR-1) and PAR-2. PARs include 4 members (PAR-1, PAR-2, PAR-3, and PAR-4), which belong to the family of G protein-coupled receptors [21 –24]. Previous papers showed that the activation of PAR-1and PAR-2 induced by FXa could be inhibited by rivaroxaban, which attenuated vascular damage and inflammation [25, 26]. However, the treatment effect of rivaroxaban against cognitive impairment in AD was still unknown [25]. Therefore, the aim of the present study was to assess the influence of rivaroxaban treatment, compare it with warfarin treatment, and investigate the roles of PAR-1 and PAR-2 induced by FXa in the AD + CAA mice model.
MATERIALS AND METHODS
Animals
All procedures were conducted in accordance with the guidelines of the Animal Committee of the Graduate School of Medicine and Dentistry of Okayama University (OKU-2018-364). The transgenic mice model of AD, which expresses human vasculotropic Swedish mutant amyloid precursor protein (APP23 mice), presents both parenchymal senile plaque and CAA. Wild type (WT) C57BL/6J mice were used as the control. Mice were divided into four groups: WT group (WT + sham surgery, n = 10), the APP23 plus CCH group (APP23 + CCH, n = 5), the APP23 plus CCH plus Warfarin group (APP23 + CCH + Warfa, n = 5), and the APP23 plus CCH plus Rivaroxaban group (APP23 + CCH + Riva, n = 6). To establish the CCH mice model, ameroid constrictors (ACs) with an inner diameter of 0.75 mm (Research Instruments NW, Lebanon, OR, USA) were applied to bilateral common carotid arteries at 4 months (M) of age [27]. We measured cerebral blood flow (CBF) at 1, 3, 7, 14, and 28 days by using a laser-doppler flowmeter (FLO-C1, Omegawave, Tokyo, Japan). From 15 day after surgery, warfarin (0.2 mg/kg/day) and rivaroxaban (60 mg/kg/day) were mixed with liquid gel (MediDrop® Sucralose, ClearH2O, Westbrook, ME, USA), and were administrated orally though bottled drinking water [28]. The optimal dosage of warfarin and rivaroxaban were determined by pre-test, as below. All mice were housed in a 12 h day/night cycle with controlled temperature and ad libitum access to liquid gel (MediDrop® Sucralose) and a standard laboratory diet (MF; Oriental Yeast, Tokyo, Japan).
Pre-test (PK/PD study)
To make sure mice in warfarin and rivaroxaban groups received same anticoagulant effect, prothrombin time-international Normalized Ratio (PT-INR) target range were controlled as 2–4 in warfarin group and 1.5–3 in rivaroxaban group, measured by Coaguchek® XS (Sekisui Medical Co. Ltd, Tokyo, Japan) [16 , 29]. These PT-INR target ranges were determined based on inhibition of clot formation by 70% in the rodent venous thromboembolism model [30 –32]. Gel water intake of a C57BL/6J mouse weight 30 g was regarded as 4 g/24 h [33], then warfarin group received 0.2 mg/kg/day of warfarin and rivaroxaban group received 60 mg/kg/day rivaroxaban, respectively [16 , 29].
Behavioral analysis
The 8-arm radial maze test was performed when mice were 3, 7, and 10 months old. The 8-arm radial maze test evaluates behavioral memory, mainly for working memory [34, 35]. For each trial, a mouse was allowed to make arm choices until either consumed all 4 pellets or 5 min elapsed. Re-entry into the baited arms previously visited was scored as a working memory error. Before formal 8-arm radial maze test, mice were placed under fasting 7 days to reduce weight to 85% of each and were allowed to explore the maze for 10 min in 2 days for adaptation.
Tissue preparation
At 10 months of age, all mice were deeply anesthetized by intraperitoneal injection of pentobarbital (40 mg/kg), then transcardially perfused with ice-cold phosphate-buffered saline (PBS, pH 7.4) and 4% ice-cold paraformaldehyde (PFA) in 0.1 mol/L phosphate buffer. Brains were removed and post-fixed in 4% PFA overnight, then brain tissues were transferred into 10, 20, and 30% (w/v) sucrose in PBS for 24 h. Brains were then rapidly frozen in liquid nitrogen and stored at –80°C. Coronal sections (20μm) were prepared on a cryostat at –22°C and mounted on silane-coated glass slides.
Immunofluorescent histochemistry
To determine vessel damage, double immunofluorescence of mice sections was performed. A TrueBlack lipofuscin autofluorescence quencher (Biotium, San Francisco, CA, USA) was used to block lipofuscin autofluorescence. For immunofluorescent histochemistry, sections were incubated overnight at 4°C with the following primary antibodies: goat anti-GFAP antibody (1:500, Abcam, Cambridge, UK) and rabbit anti-Collagen IV antibody (1:200, Abcam). The antibodies against GFAP and Collagen IV were detected with a suitable secondary antibody conjugated with Alexa Fluor (Invitrogen, Carlsbad, CA, USA). Control sections were stained in the same manner without the primary antibody. Finally, sections were covered with Vectashield mounting medium containing DAPI (Vector Laboratories) and fluorescent signals were visualized under a confocal microscope (LSM-780; Zeiss, Jena, Germany). The investigators who performed the immunohistochemical analyses were blinded to the WT, APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva assignments.
Histochemistry and immunohistochemistry
To determine microbleeds or microhemorrhages in the brains, Prussian blue staining was performed by using an Iron Stain Kit (Abcam, Cambridge, UK), according to manufacturer’s instructions.
For single immunohistochemistry, intrinsic activity of peroxidases was blocked with 0.6% hydrogen peroxide/methanol for 30 min, followed by treatment with 5% bovine serum albumin. Sections were stained overnight at 4°C with the following primary antibodies: mouse anti-amyloid β 40 (Aβ40) antibody (1:500, Fujifilm, Osaka, Japan), mouse anti-amyloid β, 17–24 antibody (4G8) (1:500, Biolegend, San Diego, CA, USA), biotin-labeled rabbit anti-mouse IgG antibody (1:500; Vector Laboratories, CA, USA), rabbit anti-MMP-9 antibody (1:100, Merck Millipore, MA, USA), rabbit anti-Iba1 antibody (1:200, Fujifilm), goat anti-GFAP antibody (1:500, Abcam), mouse anti-Thrombin R (PAR-1) antibody (1:50, Santa Cruz Biotechnology, Texas, USA), and mouse anti-PAR-2 antibody (1:50, Santa Cruz Biotechnology). After washing with PBS, antibodies against Aβ40, 4G8, IgG, MMP-9, Iba1, GFAP, PAR-1, and PAR-2 were detected with suitable secondary antibodies (1:500; Vector Laboratories) at room temperature for 2 h. Brain sections were then incubated with ABC Elite complex (Vector Laboratories) for 30 min and visualized with 3,3′-diaminobenzidine (Fujifilm). Negative control sections were stained in the same manner without the primary antibody. The investigators who performed the immunohistochemical analyses were blinded to the WT, APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva assignments.
Quantitative analysis
For each measurement, three separate sections per brain and four randomly selected regions were analyzed. We evaluated the detachment of the astrocyte end-feet from basement membrane by performed collagen IV/GFAP double immunofluorescence of mice sections. To calculate the vascular dissociation index, we measured the area between the basement membrane and the astrocyte end-feet of blood vessels, as well as the length of blood vessels [36]. We evaluated the inner diameter of collagen IV/GFAP double positive vessels and analyzed basement membrane /astrocyte remodeling in the neurovascular unit (NVU).
Statistical analysis
All data were analyzed in GraphPad Prism (version 8.3, GraphPad Software Inc., San Diego, CA, USA), and expressed as the mean±SD or median (interquartile range). For mice data, one-way analysis of variance (ANOVA) was used to compare the four groups (WT, APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva) followed by the Tukey-Kramer test. p < 0.05 was considered statistically significant.
RESULTS
ACs decreased CBF after CCH
After ACs application, the CBF gradually decreased in APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva groups 1 day after CCH surgery, reached a minimum point 7 days after surgery. The administration of warfarin and rivaroxaban started 15 days after surgery, but there was no significant difference between these 3 groups at 28 days (Fig. 1A).
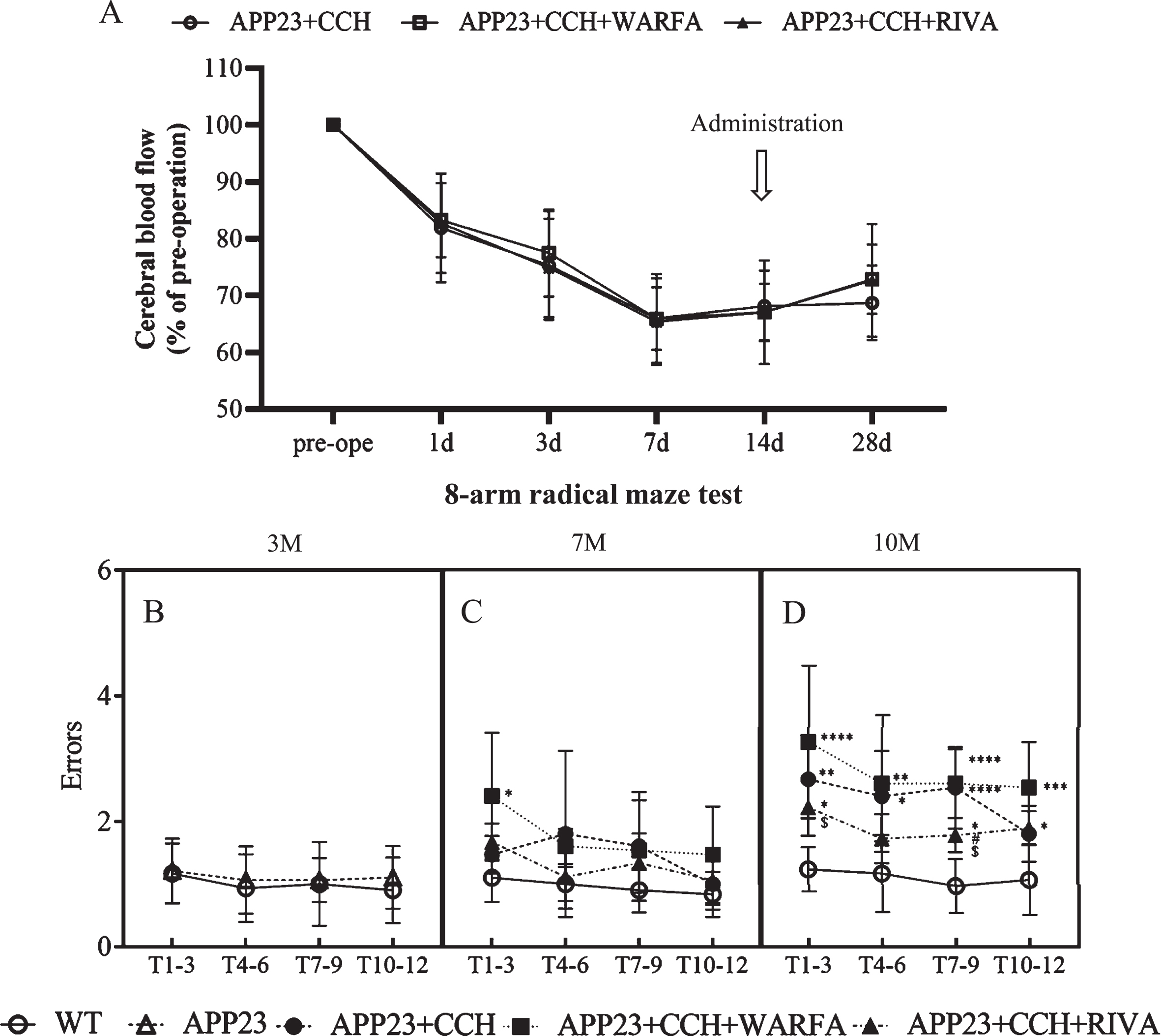
Rivaroxaban significantly improved memory deficits of AD mice model with CCH. A) Temporal profile of the cerebral blood flow measured by Laser-Doppler flowmeter. B-D) Quantitative analysis of working memory by 8-arm radial maze test (* p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 versus WT; # p < 0.05 versus APP23 + CCH; $ p < 0.05 versus APP23 + CCH + Warfa).
Rivaroxaban improved memory deficits
Before the CCH surgery, the result of 8-arm radial maze test had no differences in errors between the WT group and the APP23 group at 3 months (Fig. 1B). At 7 months, error times significantly increased in the APP23 + CCH + Warfa group compared to the WT group (Fig. 1C, * p < 0.05 versus WT, Tukey’s multiple comparison test). At 10 months, the APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva groups showed significant working memory impairment in errors compared to the WT group (Fig. 1D, * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 versus WT, Tukey’s multiple comparison test). Meanwhile, rivaroxaban treatment rescued this dysfunction compared with the APP23 + CCH and APP23 + CCH + Warfa groups at 10 months of age (Fig. 1D, # p < 0.05 versus APP23 + CCH, $ p < 0.05 versus APP23 + CCH + Warfa, Tukey’s multiple comparison test).
Warfarin aggravated microhemorrhages
We labeled ferric iron for evaluating the area of cerebral microbleeds and microhemorrhages (Fig. 2A). Iron staining positive area significantly increased around vessel in the cortex of the APP23 + CCH + Warfa group compared with the WT and APP23 + CCH + Riva groups at 10 months old (Fig. 2B, *** p < 0.001 versus WT, $ p < 0.05 versus APP23 + CCH + Riva, Tukey’s multiple comparison test). There were no significant differences between other groups.
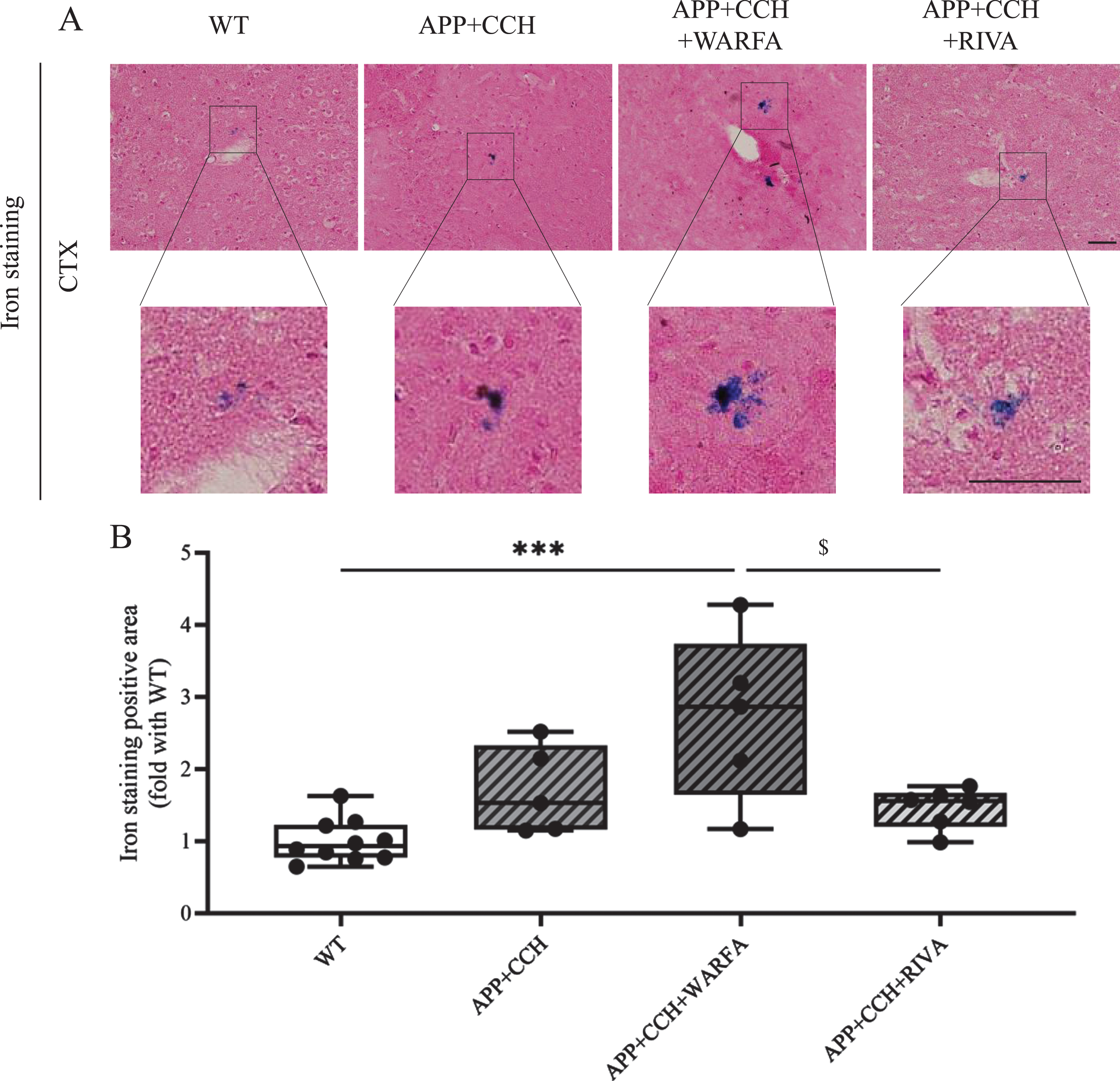
A) Iron staining in the cortex of AD plus CCH mice model. The positive area of iron staining mainly was located close to vessel. Scale bar = 50μm. B) Quantitative analysis of iron staining positive area. Note the significant decrease of iron staining positive area in the APP23 + CCH + Riva group compared to the APP23 + CCH + Warfa groups (*** p < 0.001 versus WT; $ p < 0.05 versus APP23 + CCH + Warfa).
Rivaroxaban improved both IgG leakage and vessel damage
IgG leakage was mainly found near the small vessel in the cortex of the APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva groups at 10 months old (Fig. 3A). Moreover, compared with other groups, the pixel intensity of IgG was significantly increased in APP23 + CCH + Warfa group (Fig. 3C, **** p < 0.0001 versus WT, # # # # p < 0.0001 versus APP23 + CCH, $$$$ p < 0.0001 versus APP23 + CCH + Riva, Tukey’s multiple comparison test).
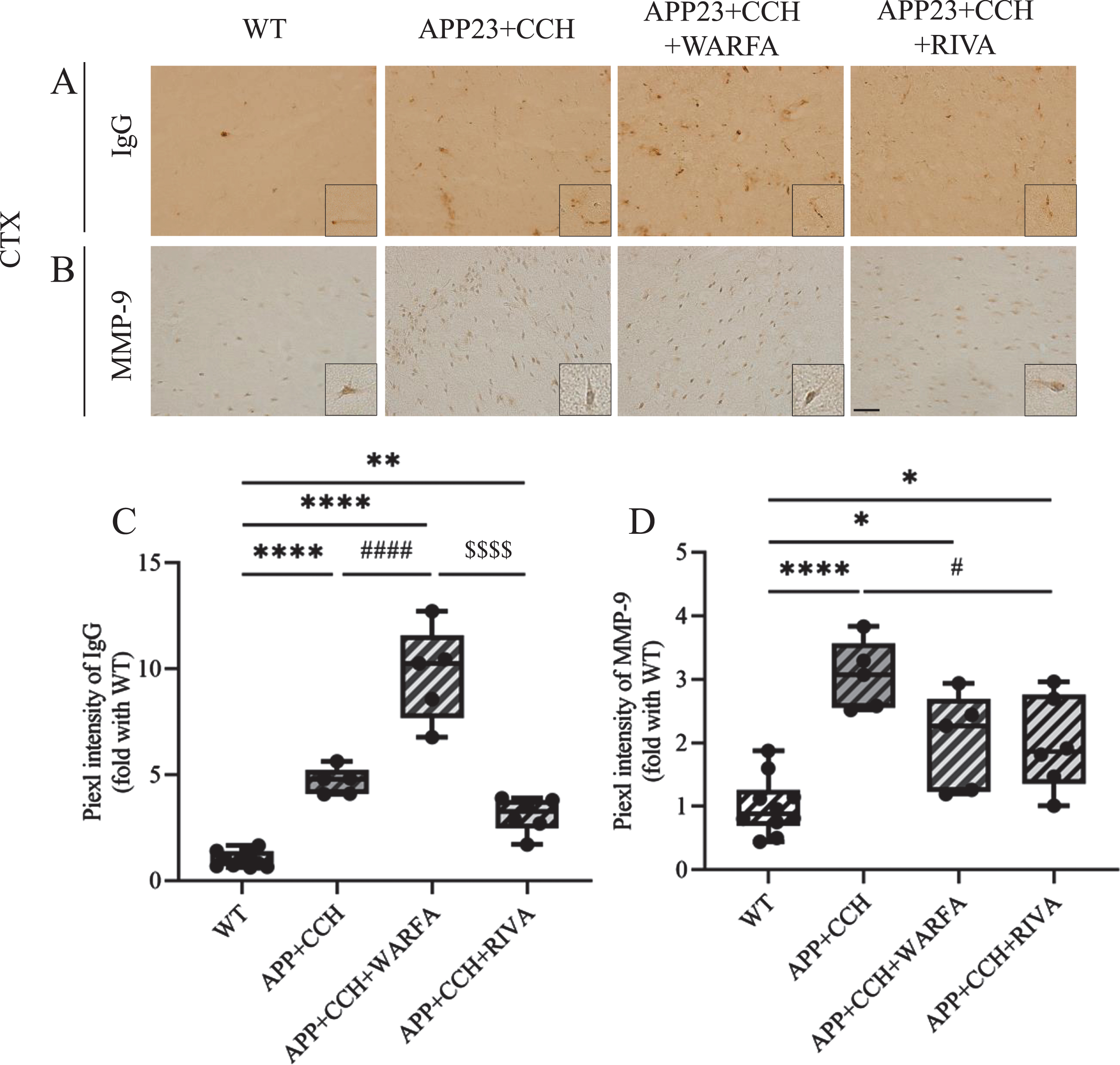
A, B) Immunochemical staining of IgG and MMP-9 in the cortex of AD plus CCH mice model. Scale bar = 50μm. C, D) Quantitative analysis of IgG and MMP-9. Note the significant increase of IgG expression in the APP23 + CCH + Warfa group compared to both APP23 + CCH and APP23 + CCH + Riva groups as well as the significant decrease of MMP-9 expression in the APP23 + CCH + Riva group compared to the APP23 + CCH group (* p < 0.05, ** p < 0.01, and **** p < 0.0001 versus WT; # p < 0.05 and # # # # p < 0.0001 versus APP23 + CCH; $$$$ p < 0.0001 versus APP23 + CCH + Warfa).
MMP-9 were widely but weakly expressed in the mice brain. Single immunohistochemistry of MMP-9 revealed that the pixel intensity of MMP-9 significantly increased in the cortex of the APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva groups compared with WT group at 10 months old (Fig. 3B, D, * p < 0.05 and **** p < 0.0001 versus WT, Tukey’s multiple comparison test). However, warfarin and rivaroxaban treatment reduced the activation of MMP-9, the pixel intensity of MMP-9 significantly decreased in the APP23 + CCH + Riva group compared with APP23 + CCH group (Fig. 3D, # p < 0.05 versus APP23 + CCH, Tukey’s multiple comparison test).
Double immunofluorescent of the basal lamina (Collagen IV) and astrocyte foot processes (GFAP) showed that the inner diameter of a part of small vessels significantly increased in the APP23 + CCH + Warfa groups compared with the APP23 + CCH group at 10 months old (Fig. 4A, B, # # # p < 0.001 versus APP23 + CCH, Tukey’s multiple comparison test). Meanwhile, rivaroxaban treatment showed significantly rescue enlargement of the inner diameter compared to both APP23 + CCH and APP23 + CCH + Warfa groups (Fig. 4B, # p < 0.05 versus APP23 + CCH, $$$$ p < 0.0001 versus APP23 + CCH + Warfa, Tukey’s multiple comparison test). At 10 months, vascular dissociation indexes significantly increased between the basal lamina and astrocyte foot processes in the APP23 + CCH + Warfa group compared to the WT group (Fig. 4C, ** p < 0.01 versus WT, Tukey’s multiple comparison test).
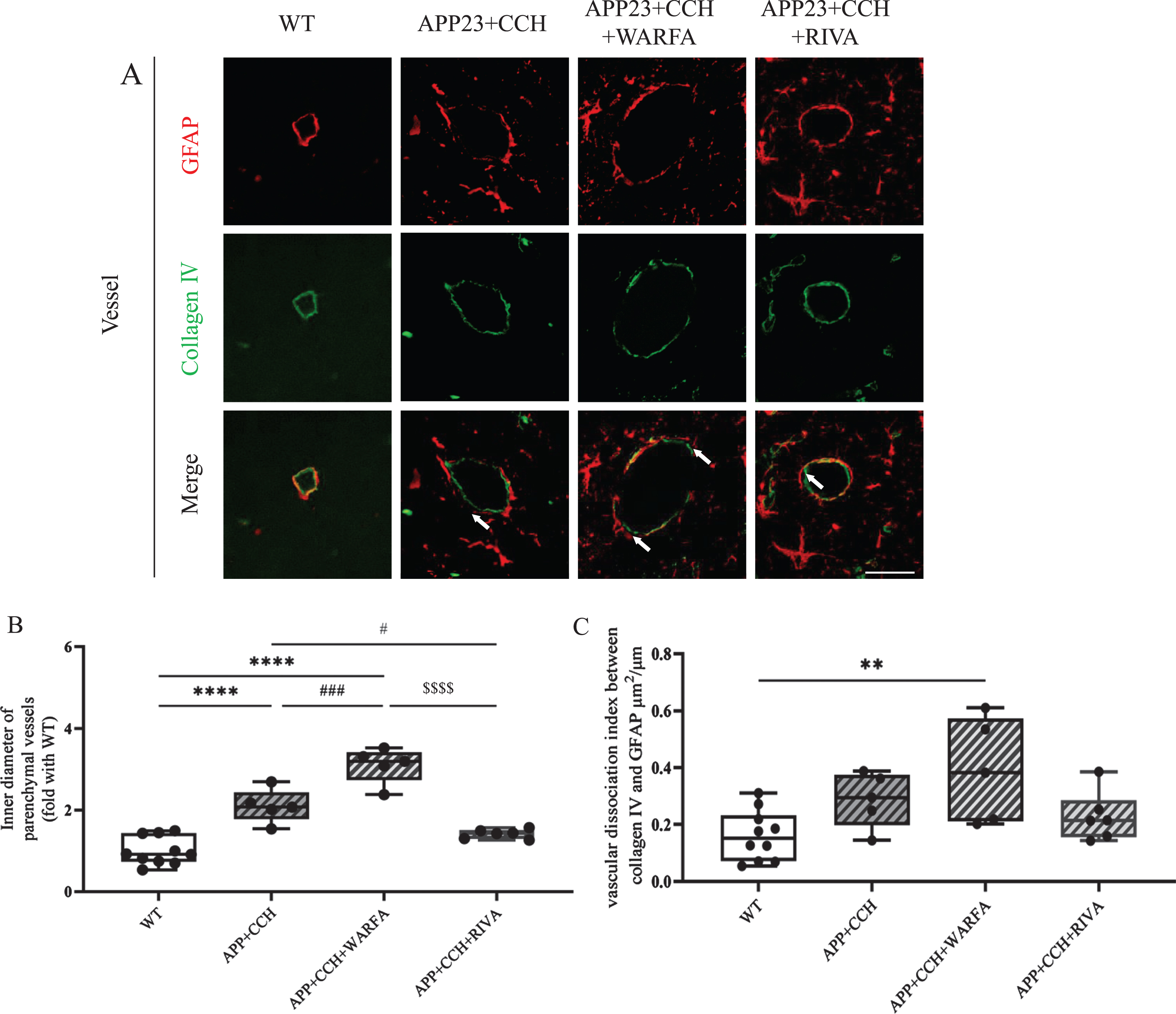
A) Double immunostaining of GFAP and Collagen IV in the AD plus CCH mice model. Arrowheads point the dissociation of Collagen IV- and GFAP-positive vessel. Scale bar = 50μm. B, C) Quantitative analysis of inner diameter of parenchymal vessels and vascular dissociation index. Note the significant decrease of inner diameter of parenchymal vessels in the APP23 + CCH + Riva group compared to both APP23 + CCH and APP23 + CCH + Warfa groups (** p < 0.01 and **** p < 0.0001 versus WT; # p < 0.05 and # # # p < 0.001 versus APP23 + CCH; $$$$ p < 0.0001 versus APP23 + CCH + Warfa).
Rivaroxaban attenuated neuroinflammation
The results showed the pixel intensity of a microglia marker (Iba-1) and an astroglia marker (GFAP) significantly increased in both cortex and hippocampus of the APP23 + CCH + Warfa group, compared with the APP23 + CCH group at 10 months old (Fig. 5A-D, # p < 0.05, # # p < 0.01 versus APP23 + CCH, Tukey’s multiple comparison test). However, rivaroxaban treatment greatly attenuated the activation of microglia and astrocyte in the cortex and hippocampus compared with the APP23 + CCH + Warfa (Fig. 5C, D, $ p < 0.05 and $$$$ p < 0.0001 versus APP23 + CCH + Warfa, Tukey’s multiple comparison test). Moreover, rivaroxaban treatment significantly attenuated the activation of microglia in the hippocampus as well as astrocyte in the cortex (Fig. 5C, D, # # p < 0.01 and # # # p < 0.001 versus APP23 + CCH, Tukey’s multiple comparison test).

A, B) Immunochemical staining of Iba-1 and GFAP in the cortex and hippocampus of AD plus CCH mice model. Scale bar = 50μm. C. D) Quantitative analysis of Iba-1 and GFAP. Note the significant decrease of Iba-1 and GFAP in the APP23 + CCH + Riva group compared to both APP23 + CCH and APP23 + CCH + Warfa groups (** p < 0.01, *** p < 0.001, and **** p < 0.0001 versus WT; # p < 0.05, # # p < 0.01, and # # # p < 0.001 versus APP23 + CCH; $ p < 0.05 and $$$$ p < 0.0001 versus APP23 + CCH + Warfa).
Rivaroxaban suppressed the expression of PAR-1 and PAR-2
PAR-1 and PAR-2 were widely but weakly expressed in the cell and vessel of mice brain. The results showed PAR-1 and PAR-2 accumulation significantly increased in the APP23 + CCH + Warfa group compared with the APP23 + CCH group at 10 months old in the cortex and hippocampus (Fig. 6A-D, # # p < 0.01 versus APP23 + CCH, Tukey’s multiple comparison test). However, rivaroxaban treatment strongly inhibited the accumulation of PAR-1 and PAR-2 in the cortex and hippocampus compared with the APP23 + CCH and APP23 + CCH + Warfa groups (Fig. 6C, D, # p < 0.05, # # p < 0.01, and # # # p < 0.001 versus APP23 + CCH, $$$ p < 0.001 and $$$$ p < 0.0001 versus APP23 + CCH + Warfa, Tukey’s multiple comparison test).
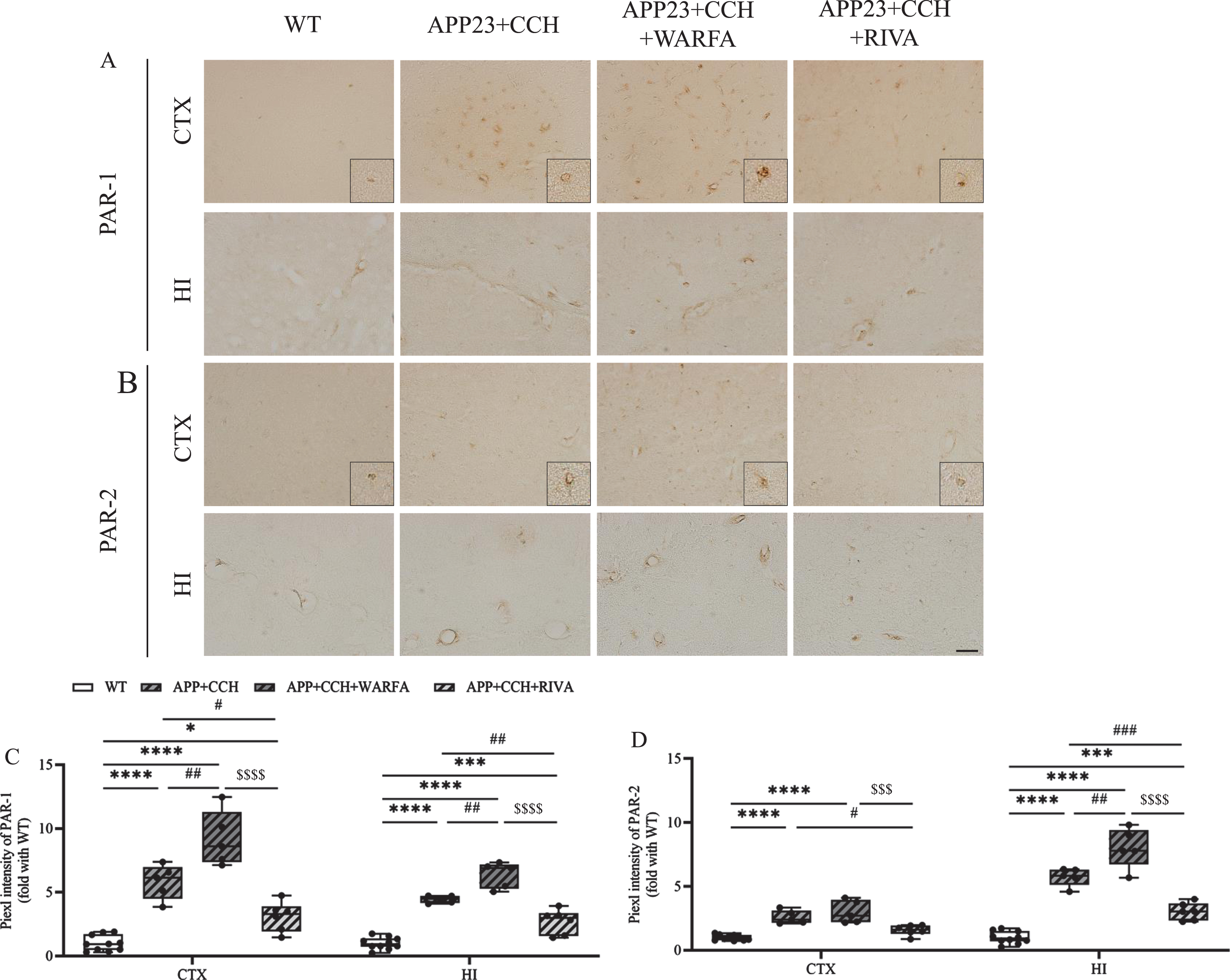
A, B) Immunochemical staining of PAR-1 and PAR-2 in the cortex and hippocampus of AD plus CCH mice model. Scale bar = 50μm. C, D) Quantitative analysis of PAR-1 and PAR-2. Note the significant decrease of PAR-1 and PAR-2 in the APP23 + CCH + Riva group compared to both APP23 + CCH and APP23 + CCH + Warfa groups (* p < 0.05, *** p < 0.001, and **** p < 0.0001 versus WT; # p < 0.05, # # p < 0.01, and # # # p < 0.001 versus APP23 + CCH; $$$ p < 0.001 and $$$$ p < 0.0001 versus APP23 + CCH + Warfa).
Rivaroxaban decreased intraneuronal and vascular Aβ deposition
The results showed more obvious differences of Aβ40 in vessel between the APP23 + CCH, APP23 + CCH + Warfa, and APP23 + CCH + Riva groups. The deposition of 4G8 and Aβ40 significantly increased in the APP23 + CCH + Warfa group compared with the APP23 + CCH group at 10 months old in both cortex and hippocampus, respectively (Fig. 7A-D, # p < 0.05 and # # # # p < 0.0001 versus APP23 + CCH, Tukey’s multiple comparison test). Rivaroxaban treated group showed significant improvement on the deposition of 4G8 and Aβ40 compared with the APP23 + CCH + Warfa groups (Fig. 7C, D, $ p < 0.05 and $$$$ p < 0.0001 versus APP23 + CCH + Warfa, Tukey’s multiple comparison test). Moreover, rivaroxaban attenuated Aβ40 deposition in the hippocampus compared to the APP23 + CCH group (Fig. 7D, #p < 0.05 versus APP23 + CCH, Tukey’s multiple comparison test).
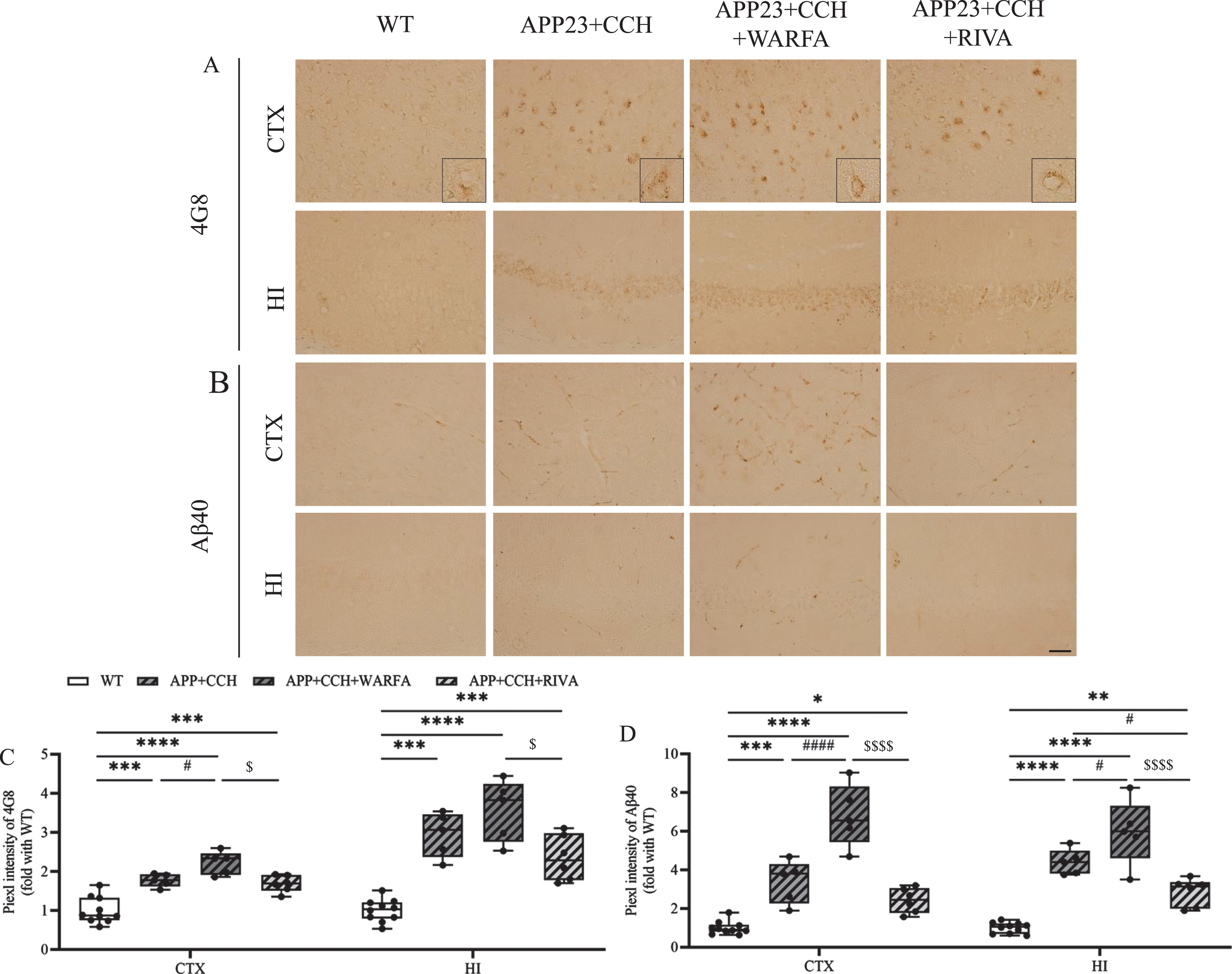
A, B) Immunochemical staining of 4G8 and Aβ40 in the cortex and hippocampus of AD plus CCH mice model. Scale bar = 50μm. C, D) Quantitative analysis of 4G8 and Aβ40. Note the significant decrease of 4G8 and Aβ40 in the APP23 + CCH + Riva group compared to the APP23 + CCH + Warfa groups (* p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001 versus WT; # p < 0.05 and # # # # p < 0.0001 versus APP23 + CCH; $ p < 0.05 and $$$$ p < 0.0001 versus APP23 + CCH + Warfa).
DISCUSSION
In the present study, we applied long-term treatments of warfarin and rivaroxaban to a novel AD + CAA mice model to evaluate their effects on the AD + CAA pathology (Figs. 1 –7). Warfarin treatment had aggravating effects on the AD + CAA mice model when mice were 10 months old (Figs. 1 –7). In contrast, rivaroxaban treatment attenuated memory decline, rescued blood-brain barrier (BBB) leakage, neuroinflammation, and Aβ deposition in the mice brain (Figs. 1 and 3 –7).
The increasing of glial cells in the central nervous system (CNS) is a result of neuroinflammation response in AD. In consequence, the increased glial cells promote the generation of cytokines, such as tumor necrosis factor α and IL-1β, and further enhance inflammation and BBB damage [37, 38]. PAR-1 and PAR-2 can cause the activation of glial cells, express throughout the CNS, including vascular endothelial cells, microglia, astrocytes, and neurons [39 –43]. Previous reports showed that the inhibition of PAR-1 and PAR-2 could prevent the activation of microglia cells and astrocytes in the CNS [44 –47], indicated that the activation of PAR-1 and PAR-2 may be related to inflammation around vessels and BBB damage. In the present study, PAR-1 and PAR-2 decreased in the APP23 + CCH + Riva group at 10 months old, accompanied by a decreased activation of microglia cells and astrocytes (Figs. 5 6), which suggested that the proliferation of astrocyte and microglia were suppressed through the inhibitory effect of rivaroxaban to PAR-1 and PAR-2. Therefore, rivaroxaban attenuated neuroinflammation and showed a protective effect in BBB damage. Increased expression of MMP-9 was observed in AD patients, which had a correlation with increased BBB permeability and degradation of extracellular matrix components [48, 49]. Our results showed the activation of MMP-9 significantly decreased in the APP23 + CCH + Riva group compared to APP23 + CCH group (Fig. 3), indicated that this decreased MMP-9 activation may relate to the inhibition of PAR-1 [50]. The activation of glial cells and increased expression of PAR-1/PAR-2 were observed in the APP23 + CCH + Warfa group; however, the MMP-9 expression was unchanged in the present study.
In the APP23 + CCH + Warfa group, the increasing of vessel remodeling and vascular dissociation suggested an aggravated NVU dysfunction. However, vessel remodeling was dramatically improved in the rivaroxaban group (Fig. 4). Warfarin treatment inhibited multiple coagulation factors, leading to PAR-1 and PAR-2 increasing, then aggravated vessel inflammation, which could be a reason for reversing the positive effect on thrombin inhibition and aggravated NVU and BBB dysfunction and neuroinflammation in AD mice [51, 52]. In contrast, rivaroxaban inhibited the expression of PAR-1 and PAR-2, which were induced by FXa, prevented astrocyte- and microglia-induced inflammation, and also prevented microvascular hyperpermeability by decreasing MMP-9 expression, suggesting that rivaroxaban protects NVU in AD + CAA mice [53, 54]. Moreover, thrombin-mediated PAR-1 activation can increase endothelial barrier permeability through mitogen-activated protein kinases. Meanwhile, activated protein C (APC)-mediated activation of PAR-1 provides an endothelial barrier-protective effect [51 , 54]. As a hypothesis, rivaroxaban decreased neuroinflammation by inhibiting FXa-mediated activation of PAR-1 but had less inhibition to APC-induced endothelial barrier protective effect in contrast with warfarin, this suppressed inflammation may cause less NVU dysfunction.
Our previous paper showed a relative downregulation of PAR-1 and PAR-2 by rivaroxaban compared with warfarin pretreatment in a stroke model, which might be related to the reduction of hemorrhagic complications [25]. In the present study, PAR-1 and PAR-2 decreased in the APP23 + CCH + Riva group (Fig. 6) and showed less microhemorrhages and IgG leakage compared to warfarin treatment (Figs. 2 3), which suggested rivaroxaban might decrease microhemorrhages and IgG leakage through inhibition of PAR-1 and PAR-2 in AD mice. According to our results, these effects may be caused by the protective effect of rivaroxaban to NVU and BBB. Meanwhile, 8-arm radial maze test showed that error numbers in the APP23 + CCH + Riva mice was decreased at 10 months old, which indicated that rivaroxaban significantly recovered memory decline in the APP23 + CCH mice at 10 months old (Fig. 1). The results of single immunohistochemistry of 4G8 and Aβ40 suggested that rivaroxaban decreased Aβ accumulation in the APP23 + CCH mice model, more obvious change of Aβ accumulation was observed in small vessels than cells (Fig. 7). Recently, some studies suggested that neuroinflammation is an early event in AD as well as one of the important factors promoting AD pathogenesis. CAA pathogenesis is associated with multiple factors, such as NVU and BBB dysfunction and hemorrhage [5 , 8]. Moreover, neuroinflammation, BBB and NVU dysfunction, and CAA pathology may form a positive feedback loop in a milieu enriched by the accumulation of Aβ [55 –57]. Intracellular Aβ in AD can activate astroglia and microglia, increasing the inflammatory effect on NVU and BBB. The cytokines generated by reactive astrocytes and microglia may cause the dysfunction of NVU and BBB, involving vascular wall thickening, vascular remodeling, a reduction in smooth muscle cells, and vascular dissociation. This subsequently causes a reduction in CBF and a defect in the clearance of Aβ, then triggers an increase in the accumulation of cerebrovascular Aβ to form CAA, leading to more severe neuroinflammation and dysfunction of the BBB and NVU. This feedback loop could lead to the loss of neurons and cognitive deficits [8 , 58–60]. However, rivaroxaban treatment reduced this feedback effect by inhibiting the expression of astrocytes and microglia. This decrease in neuroinflammation may attenuate the dysfunction of NVU and BBB, finally reducing the accumulation of Aβ in the vessel. These results suggested that rivaroxaban treatment attenuated CAA pathology, which may be a cause leading to memory decline improvement.
In summary, the present study demonstrated that rivaroxaban treatment attenuated neuroinflammation, BBB dysfunction, memory deficits, and Aβ deposition in a novel AD + CAA mice model. The inhibition of PAR-1 and PAR-2 induced by rivaroxaban may play an important role in the attenuation of neuroinflammation and in CAA pathology. Therefore, rivaroxaban might, compared to warfarin, reduce the risk of causing AD + CAA when applied to old patients. Moreover, rivaroxaban reduced the progress of AD, indicating that rivaroxaban could be a potential choice to prevent AD + CAA.