
Abstract
Background:
There exists considerable interest in the identification of molecular traits during early stages of Alzheimer’s disease (AD). Mild cognitive impairment (MCI) is considered the closest prodromal stage of AD, and to develop gradually from earlier stages although not always progresses to AD. Classical cerebrospinal fluid (CSF) AD biomarkers, amyloid-β peptides and tau/p-tau proteins, have been measured in prodromal stages yet results are heterogeneous and far from conclusive. Therefore, there exists a pressing need to identify a neurochemical signature for prodromal stages and to predict which cases might progress to AD.
Objective:
Exploring potential CSF biomarkers related to brain oxidative and inorganic biochemistry during prodromal stages of the disease.
Methods:
We have analyzed CSF levels of lipoxidative markers (MDA and 8-isoF2α), biometals (Cu, Zn, Se, Mn, and Fe), iron-transport protein transferrin (TFER), antioxidant enzymes (SOD and GPx4), detoxifying enzymes (GST and BuChE), as well as classical amyloid-β and total and phosphorylated tau, in cognitively healthy controls, patients with MCI, and subjects exhibiting subjective memory complaints (SMC).
Results:
Inter-group differences for several variables exhibit differentiable trends along the HC ⟶ SMC ⟶ MCI sequence. More interestingly, the combination of Se, Cu, Zn, SOD, TFER, and GST variables allow differentiable fingerprints for control subjects and each prodromal stage. Further, multivariate scores correlate positively with neurocognitive In-Out test, hence with both episodic memory decline and prediction to dementia.
Conclusion:
We conclude that changes in the CSF biochemistry related to brain oxidative defense and neurometallomics might provide more powerful and accurate diagnostic tools in preclinical stages of AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is the most common cause of dementia in the elderly. Epidemiological evidence support the notion that AD has a long preclinical period, including an interim phase known as mild cognitive impairment (MCI) characterized by measurable memory and cognitive declines [1 –3]. It is estimated that evolution from MCI to AD occurs at a rate of 10–15% per year, with 80% conversion by the sixth year of follow-up [4, 5]. It is accepted that MCI is associated with a long phase of neuropathological changes before cognitive decline is diagnosed [6]. Since once developed, AD is irreversible and has no cure, it is of enormous importance the development of biomarkers for monitoring neurochemical or neuroimaging abnormalities linked to disease progression from MCI to AD. The lack of effective treatments for overt AD urges the discovery of biomarkers for early diagnosis, i.e., in subjects with MCI or prodromal AD.
It is widely accepted that oxidative stress plays a critical role in the pathophysiology of AD and that this phenomenon is closed related to the formation and reactivity of amyloid plaques (SPs), a main histopathological hallmark of the disease [7 –9]. Cerebral tissue is particularly vulnerable to oxidative-stress derived reactive species because of its high content of polyunsaturated fatty acids susceptible for oxidative modification, its high oxygen requirements for aerobic metabolism, and its low content of antioxidants [7 , 11]. Oxidative damage caused by reactive oxygen and nitrogen species (ROS and RNS, respectively) affects nearly all structures and biochemical components in the cell, impairing homeostasis and leading to nerve cell death [10 , 13]. A number of studies have reported molecular signs of oxidative damage in postmortem AD brains and cerebrospinal fluid (CSF) of AD patients [7 , 14–18]. By-products derived from lipid oxidation include reactive di-aldehydes such malondialdehyde (MDA), α,β-unsaturated aldehydes such 4-hydroxy-2-trans-nonenal (HNE), 4-hydroxy-2-trans-hexenal, and 2-propene-1-al (acrolein), and γ-ketoaldehydes formed via the isoprostane pathway from arachidonic acid peroxidation, have been consistently found elevated in AD brain regions related to cognition and also in the CSF of AD patients [1 , 19–22]. These reactive carbonyl species originating from oxidative lipid degradation have been found to generate advanced lipoxidation end products. Advanced lipoxidation end products are protein modifications formed by the reaction of nucleophilic residues of proteins (mainly Cys, Lys, His, and Arg) with reactive carbonyl species to form Michael adducts [23 –25], which are commonly detected during neurodegenerative progression and, noticeably, also during aging [26 –28]. These findings occur in parallel with reduced activity of antioxidant and detoxifying systems which are essential for reducing excess of oxidative and nitrosative by-products [14 , 29–31]. Noticeably, the evidence accumulated so far in MCI patients suggests that oxidative imbalance is already present in this precursor phase of AD, with patients showing increased levels of oxidative markers in the same brain regions and also in the CSF [19 , 22].
In close connection with ROS generation and AD is neurometallomics. A number of studies have reported chemical signatures of iron, copper, zinc, and selenium in AD. Four main roles of trace elements are related to AD. First, ionic zinc, copper, and iron facilitate amyloid-β aggregation and increase toxicity by ROS generation from the ternary complexes [32 –35]. Second, may induce ferroptosis, a recently discovered lipoperoxide-related apoptotic death pathway, which is triggered by elevated iron and/or reduced selenium levels in brain parenchyma [36 –38], Third, key enzymes from classical glutathione and thioredoxin antioxidant systems in nerve cells are metalloproteins, and display absolute requirements of metals (Cu, Fe, Zn, or Mn) for enzyme activity [33 , 39], or even for protein biosynthesis, such selenium (a non-metal required for selenocystein synthesis) in selenoproteins [38 , 41]. Consequently, biometal dyshomeostasis impacts brain metabolism at different levels which converge in the development and very likely in the onset of neurodegenerative diseases [36 , 43].
One of the intrinsic limits of most neuropathological studies on AD is that they are carried out mainly postmortem or in overt stages of dementia, giving no information about the early changes of the disease. Another pitfall is that neurochemistry of ROS and trace metals is not specific for AD, but rather shared by other neurodegenerative diseases, such Parkinson’s disease, amyotrophic lateral sclerosis, and Lewy body disease, among others [44].
In the present study, we have aimed to determine levels of lipoxidative markers, trace elements, transport proteins, and antioxidant and detoxifying enzymes associated with neurodegenerative diseases, including AD, in the CSF of healthy subjects and patients with prodromal and preclinical stages of AD. We have compiled simultaneous data from subjects in control and early stages of cognitive impairments. We have used a multivariate approach to generate a matrix of alterations which might provide a signature of disease progression in the CSF of subjects in the AD continuum. The results point to a multiparameter panel which provides a potential pathological scheme of AD progression from preclinical stages. The changes in this multivariable approach in the CSF could help to understand the biochemical processes underlying the early pathology associated with AD.
MATERIALS AND METHODS
Participants
A cohort of 53 subjects were consecutively recruited at Hospital Universitario de Gran Canaria Dr. Negrin and Hospital Universitario de Canarias (Spain) in accordance with ethical principles stated in the Declaration of Helsinki, as well as with approved national and international guidelines for human research.
The inclusion criteria were consecutive outpatients at the Neurology service, older than 59 years, able to read and write, and consulting for memory complaints. All individuals provided informed consent and underwent an evaluation that consisted of medical history, physical and neurological examinations, laboratory tests, and neuropsychological assessments. The patients were classified into the three study groups described below.
Healthy control (HC) patients were recruited at the orthopedic surgery service scheduled for a CSF study. These patients are characterized by alternative diagnoses but do not show any neurological signs or positive marker profiles indicative of neurological disease. Subjective memory complaints (SMC) group includes patients who refer for cognitive complaints but had normal performance in a full neuropsychological evaluation without impact in daily living. Finally, the MCI group was classified following the criteria from the report of the MCI working group of the European Consortium on Alzheimer’s disease (EADC) [45].
Subjects were assigned to each group using the Global Dementia Scale (GDS). This scale classifies HC as GDS1 for those participants exhibiting neither cognitive complaints nor dementia; GDS2 for patients of group SMC, who refer cognitive complaints but had normal performance in neuropsychological evaluation; and GDS3 for patients of MCI group, presenting a low performance in at least one cognitive function (more than 1.5 standard deviations below the mean, according to the Spanish data from NORMACODEM and NEURONORMA in not demented patients [46 –49]. A comprehensive overview of cohort data is summarized in Table 1.
General description of group characteristics used in this study
Data are expressed as mean±SD. M, men; W, women; MMSE, Mini-Mental State Examination. Different letters in a variable indicate statistically significant differences with p < 0.05 (post hoc Tukey’s HSD).
The exclusion criteria were: suspected focal or diffuse brain damage due to clinical conditions different to AD; uncontrolled systemic diseases or delirium in the last 30 days; history of drug addiction or alcoholism; being under treatment for AD; history of major depression or being under treatment with two or more antidepressants; more than one dose per day of benzodiazepines; severe perceptive or motor disorders. Patients with severe anxiety and depression symptoms were excluded according to the Hospital Anxiety and Depression Scale evaluation [50, 51].
CSF samples
CSF samples were obtained by lumbar puncture in polypropylene tubes following a standard procedure. Once collected, samples were centrifuged at 2,000 g for 20 min at 4°C and frozen at –80°C until analysis.
Determination of selenium, iron, copper, zinc, and manganese
CSF samples were thawed slowly at 4°C before being diluted 1:20 in a solution of 5% HNO3 with 1000 ppm of Rh as internal standard to determinate total concentration of Se, Cu, Fe, and Mn. Then, proteins were removed by filtering samples through a 0.45-micron filter. In the inductively coupled plasma mass spectrometry analysis, a lyophilized serum control for trace elements (ClinChek® Controls, Germany) was used to ensure the accuracy of the analytical methods and results. Concentrations of metals were determined by interpolation of the absorbance value into a multitrace standard curve and were expressed by μmol/ml CSF. Elemental analysis was performed by inductively coupled plasma mass spectrometry, using the Agilent 7500ce collision–reaction cell system (Agilent Technologies, Tokyo, Japan), with helium of high-purity grade (> 99.99%) as collision gas.
SOD, GST, GPx4, and BuChE activities
Superoxide dismutase (SOD) activity was measured using the pyrogallol method following by Mesa-Herrera and collaborators (2019) [52]. One unit was defined as 50% inhibition of the rate of autoxidation of pyrogallol. The activity of SOD is expressed as mUnits/mg protein.
Glutathione-S-transferase (GST) activity was determined following the conjugation of GSH with CDNB (1-chloro-2,4-dinitrobenzene) at 340 nm [53] and expressed as nmol GS-DNB/hr.mg protein.
Total phospholipid-hydroperoxide glutathione peroxidase (GPx4) activity was measured using the glutathione reductase-NADPH methods described by Lawrence and Burk (1976), by monitoring the rate of decrease in the concentration of NADPH as recorded at 340 nm. GPx4 activity was expressed as μmol NADPH/hr.mg protein [54].
Butyrylcholinesterase (BuChE) activity was determined using the method develop by Ellman et al., 1961 and adapted by Wheelock and collaborators (2005) [55]. The enzyme activity was determined by following the increase in absorbance at 412 nm produced by the formation of the compound resulting from the reaction of thiocholine with DTNB. BuChE activity was expressed as μmol thiocholine/hr.mg protein.
Spectrophotometric measurements were carried out in a VICTORTM X5 spectrophotometer (Perkin Elmer). Enzyme activities units were normalized to mg protein, which were determined by the method of Bradford [56].
Transferrin levels
CSF transferrin was determined using an ELISA assay. Briefly, 10μg of CSF proteins were immobilized using carbonate-sodium bicarbonate buffer (0.07M NaHCO3, 0.03M Na2CO3, pH 9.6 in double-distilled water) to the substrate residing at the bottom of the ELISA plates and were incubated for 16–18 h under agitation at 4°C. Wells were washed with 0.05% PBS-Tween20 buffer. Subsequently, wells were incubated with specific primary antibodies diluted in a 5% PBS-BSA solution during 16–18 h at 24°C. The primary antibody was recognized by a secondary peroxidase-bound antibody (anti-rabbit or anti-mouse, diluted in PBS-T). Then, secondary peroxidase-bound antibodies were oxidized using 1-StepTM Ultra TMB-ELISA (Thermo Fisher) during 30 min and the reaction was stopped by adding 1N H2SO4. Absorbance measurements were carried out at 450 nm wavelength in a VICTORTM X5 spectrophotometer (Perkin Elmer).
Malondialdehyde concentration
MDA lipoperoxide was determined by the thiobarbituric acid reacting substances (TBARs) method [57], using TMP (1,1,3,3 tetramethoxypropane) as standard for calibration curves. TBARS were measured fluorimetrically with 485 nm (excitation)/535 nm (emission) wavelengths using an Appliskan Microplate Reader (Thermofisher). TBARS contents were expressed as nmol/ml CSF.
8-Isoprostane-F2α concentration
8-isoprostane-F2α concentration was determined using OxiSelect 8-isoprostaglandin- F2α commercial ELISA kits (Cell biolabs) following the manufacturer’s instructions. Concentration was expressed as pg/ml of CSF.
Amyloid-β, total tau, and phosphorylated tau concentrations
CSF concentrations of amyloid-β1 - 42 peptide, total-tau, and phospho-tau (p-tau181p) were determined using appropriate ELISA-kits (INNOTEST®, Fujirebio) following the manufacturer’s instructions. Concentration were expressed as pg/ml CSF.
Statistics
Data were initially assessed by analysis of variance (ANOVA) followed by Tukey’s or Games-Howell post hoc tests, where appropriate. Kruskal-Wallis and Mann-Whitney U tests were used for variables where normality was not achieved. Multivariate statistics was performed using principal component analyses (PCA), hierarchical cluster analyses, and discriminant function analysis (DFA). Predictive variables in DFA were chosen according to the number of cases in each group to fulfill the assumptions of discriminant analysis. Canonical scores for each case were calculated and plotted along with group centroids to indicate adscription to a particular group. One-way univariate and multivariate ANOVA were performed with age, gender, amyloid-β, total tau, and phosphorylated tau as covariates. Pearson correlation analyses and lineal regression analyses were performed to assess the significance of bivariate relationships between variables within each group.
RESULTS
Lipoxidative markers
Results from MDA and Iso-F2α analyses in the three groups are shown in the boxplots in Fig. 1. For MDA, ANOVA analyses indicate differences between groups (F2,45 = 3.75, p = 0.032). A significant increase in MDA concentration was observed in the MCI group as compared to the SMC. However, differences were not significant when comparing HC with SMC or MCI (Fig. 1A). On the other hand, no changes were observed for isoprostane F2α levels (Fig. 1B) between groups (F2,43 = 0.29, p = 750).
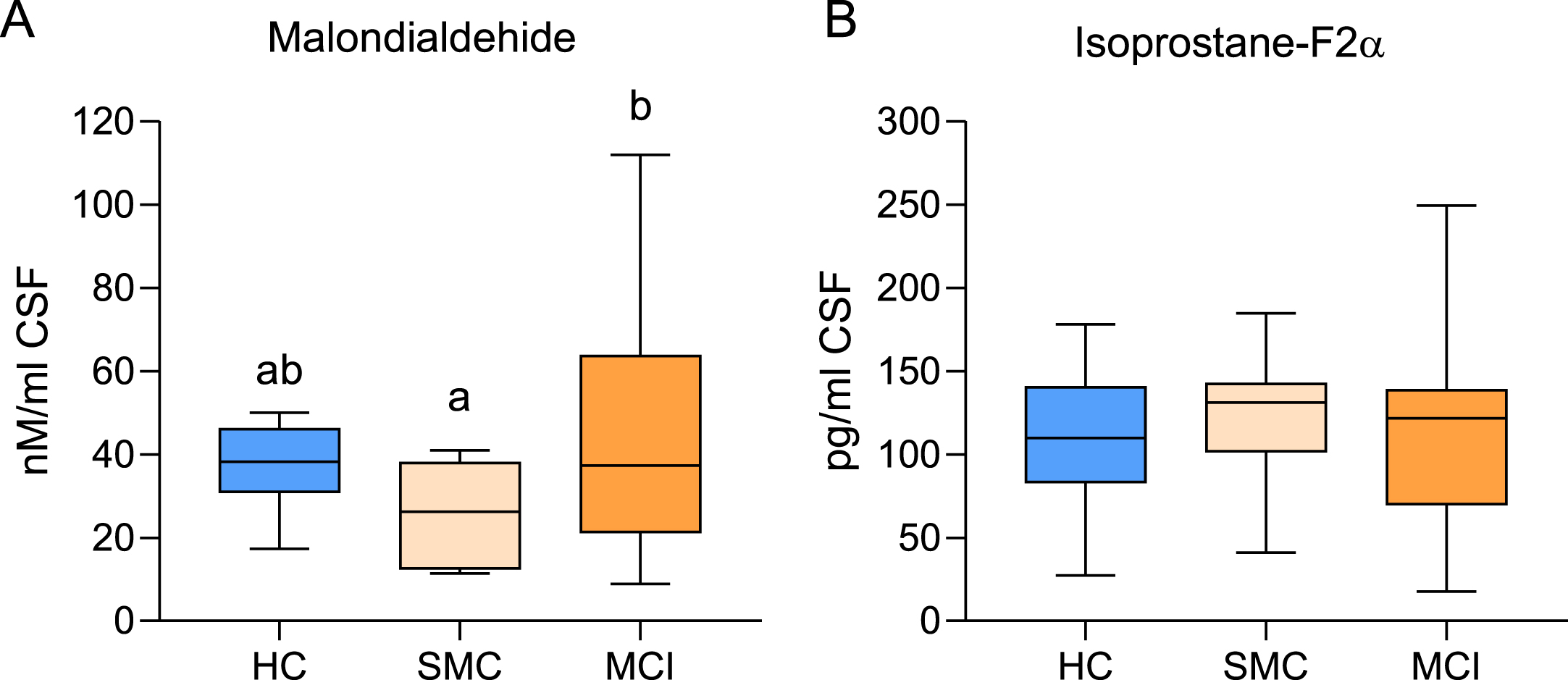
Box plots for MDA (A) and Isoprostane-F2α (B) concentrations in the CSF of healthy controls (HC, n = 15), subjective memory complaints (SMC, n = 8), and mild cognitive impairment (MCI, n = 25) groups. Different letters in the plots indicate statistically significant differences with p < 0.05 (post hoc Tukey’s HSD).
Trace biometal concentrations
The concentrations of the trace metals for each of the groups are shown in Table 2. A common feature for all biometals is the reduction of CSF levels in MCI subjects. However, ANOVA results indicate that differences between groups were significant for Se (F2,50 = 12.70, p = 0.000), Zn (F2,46 = 42.33, p = 0.000), Cu (F2,45 = 4.20, p = 0.021), and Mn (F2,45 = 2.82, p = 0.046). In the case of selenium and zinc, concentrations decrease gradually in the HC ⟶ MCI transition, with SMC group showing intermediate levels which, in turn, differ significantly from both HC and MCI groups. Conversely, in the case of copper and Mn, maximal reduction occurs earlier, in the SMC group, with no further changes towards the MCI stage.
Concentration of trace biometals in the CSF in the three groups
Concentrations are expressed in μmol/ml CSF. IC95% : 95% confidence interval for the mean. n, sample size. Different letters within rows indicate statistically significant differences at p < 0.05. Letters followed by # indicate statistical differences reached at p < 0.10 (post hoc Tukey’s HSD).
Transferrin and iron
The iron-transporting protein transferrin (TFER) was detected in the CSF of all groups, and the statistics revealed inter-group differences (F2,52 = 3.69, p = 0.034). Levels of TFER increased gradually in the HC-to-MCI transition and were significantly higher in the MCI group (Fig. 2A). Thus, we found worthwhile assessing CSF iron levels in the same subjects. The results indicate a near significant difference between groups (F2,46 = 2.86, p = 0.068). Inter-group comparisons suggests that Fe levels decreased to a minimum in SMC subjects and remained similar in MCI group (Fig. 2B).

Box plots for transferrin levels (A) and iron (B) concentration in healthy controls (HC, n = 15), subjective memory complaints (SMC, n = 9), mild cognitive impairment (MCI, n = 28) groups. Different letters within a plot indicate statistically significant differences with p < 0.05 (post hoc Tukey’s HSD).
Antioxidant and detoxifying activities
We next have analyzed the enzyme activities of a selection of enzymes essential for buffering reactive species and inactivation/removal of oxidized phospholipids and lipid adducts. These include antioxidants and detoxifying enzymes SOD, GPx4, GST, and BuChE. Results from ANOVA indicate significant differences for SOD (F2,45 = 3.47, p = 0.039), GST (F2,49 = 2.72, p = 0.076), and BuChE (F2,52 = 2.34, p = 0.061). Results shown in Fig. 3A reveal the increase in SOD activity in MCI cases compared to age-matched control, which was significantly higher than for SMC. No statistical differences were observed between MCI and HC groups, though SOD activity in this later group was 39.5% lower than in MCI. With regards to GST activity, results in Fig. 3B indicate a significant increase in SMC compared to HC (413.6%). This increase in GST activity compared to HC was slightly reduced in MCI group (264.3%). No statistically significant differences were observed for GPx4 activity between groups (Fig. 3C). Finally, BuChE activity decreased in the SMC group when compared to controls (48.4%) and remained similar in MCI group (Fig. 3D).
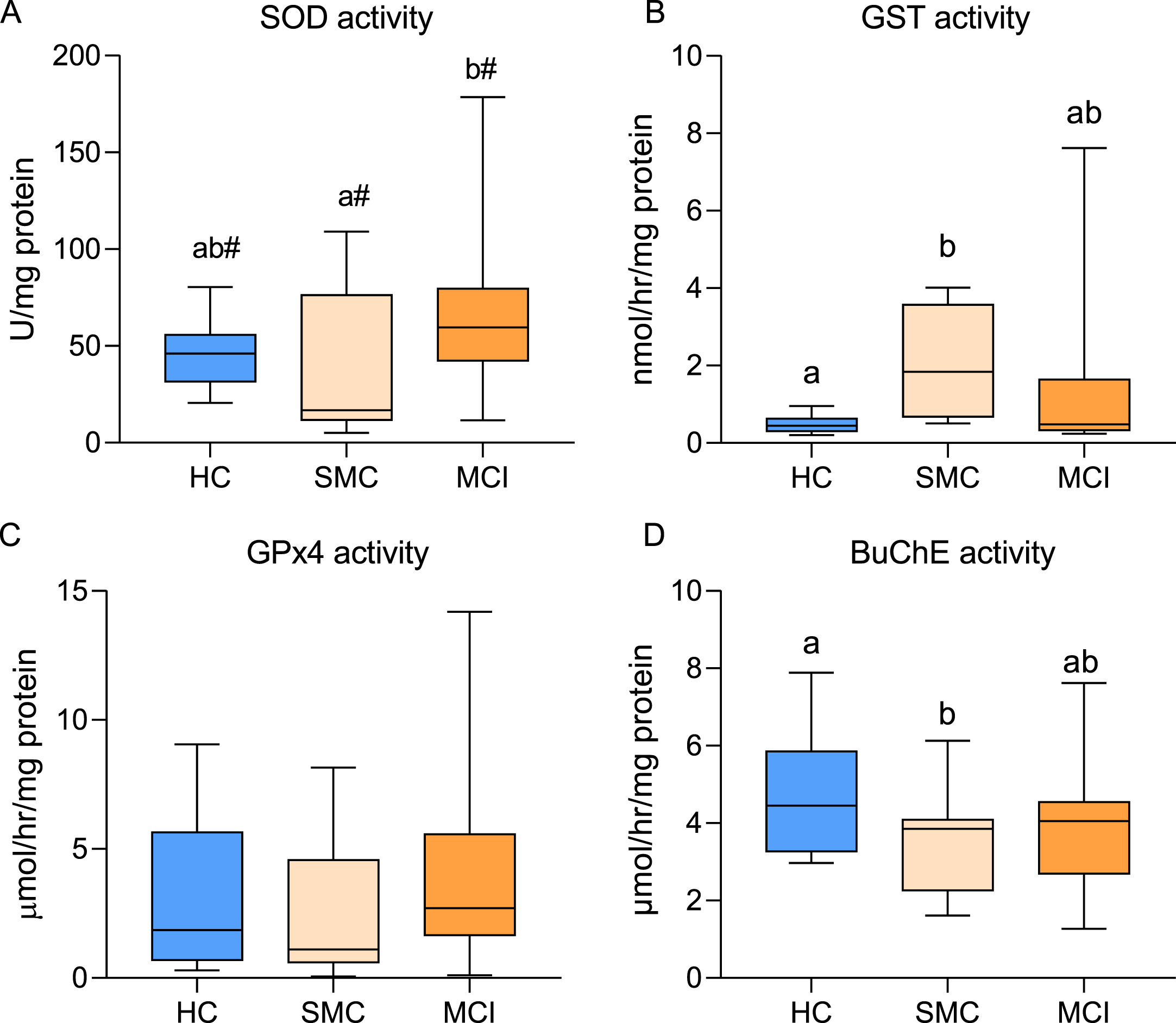
Box plots for superoxide dismutase (SOD, A), glutathione-S-transferase (GST, B), glutathione peroxidase 4 (GPx4, C), and butyrylcholinesterase (BuChE, D) activities in healthy controls (HC, n = 15), subjective memory complaints (SMC, n = 9), and mild cognitive impairment (MCI, n = 29) groups. Within each plot, different letters indicate statistically significant differences with p < 0.05. #indicates differences at p < 0.10 (post hoc Tukey’s HSD).
Bivariate relationships
Once the changes in the different variables in the three study groups were assessed, we proceeded to evaluate the intrinsic intra-group relationships between variables, and whether they might be altered along the disease continuum. We initially obtained the correlation matrixes for all groups, and the results are shown in Fig. 4.
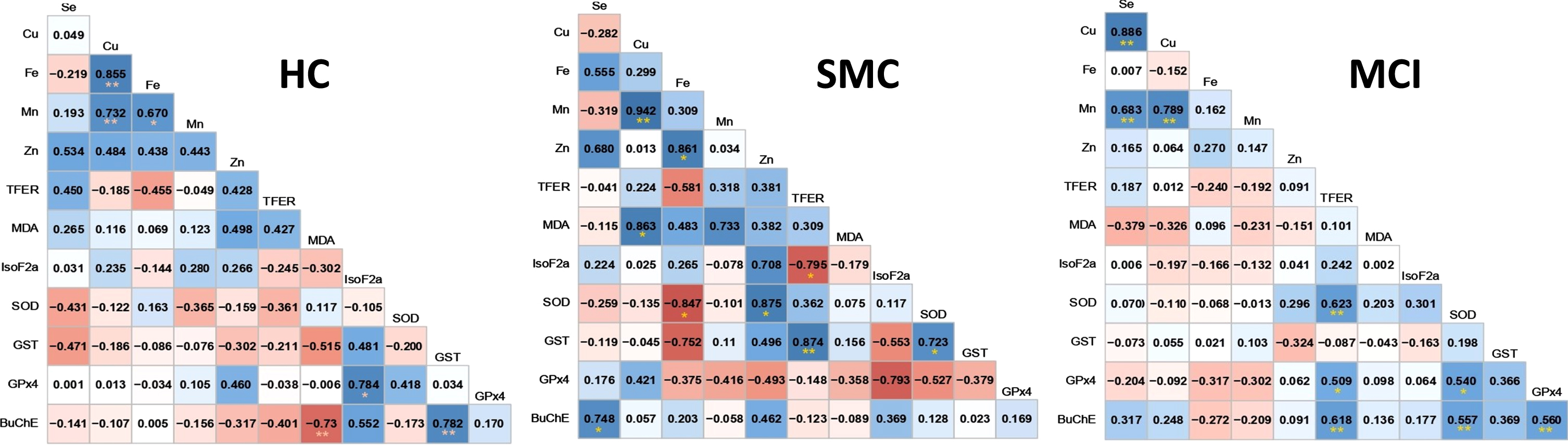
Correlograms for healthy controls (HC) subjective memory complaints (SMC), and mild cognitive impairment (MCI) groups. Significant relationships (p < 0.05) are indicated with * and p < 0.01 with **. Positive correlations are colored in blue and negative correlations in red.
For HC group, significant positive correlations were found for Cu/Fe (r = 0.855; p = 0.000), Cu/Mn (r = 0.734; p = 0.003), Fe/Mn (r = 0.670; p = 0.017), and Iso-F2α/GPx4 (r = 0.784; p = 0.021) pairs. In this group the only negatively correlated pair was MDA/BuChE (r = –0.730; p = 0.020). For the SMC group, most of the observed relationships between the variables for the HC group vanished (with the exception of Cu/Mn, r = 0.94; p = 0.000) and new relationships between the variables appeared. Thus, trace metals appeared correlated positively with different enzyme activities and metabolites, i.e., Se/BuChE (r = 0.748: p = 0.020), Zn/SOD (r = 0.862; p = 0.022), and Cu/MDA (r = 0.861; p = 0.028), and negatively to Fe/SOD (r = –0.848; p = 0.033). In SMC group, transferrin comes into scene and correlates negatively with IsoF2α (r = –0.793; p = 0.033), and positively with GST (r = 0.873; p = 0.005). Additionally, in this group, SOD and GST are also correlated (r = 0.722, p = 0.043) and copper with Mn (r = 0.940; p = 0.002). Finally, in the MCI group there is drastic decrease in the number of significant correlations (R-squared above 0.49). In this group, only Cu/Se (r = 0.887; p = 0.000) and Cu/Mn (r = 0.788; p = 0.000) fulfilled these criteria. However, as essentially relevant, BuChE becomes more important and relates with TFER (r = 0.618; p = 0.000), SOD (r = 0.557; p = 0.003), and GPx4 (r = 0.560; p = 0.005). Transferrin correlates positively with SOD (r = 0.623; p = 0.001) and GPx4 (r = 0.509; p = 0.016) but, unexpectedly, was unrelated to Fe. Finally, SOD also appear correlated positively with GPx4 (r = 0.541; p = 0.011).
Relationships with amyloid-β, total tau, and p-tau
We also assessed the relationships between the different variables under study and classical biomarkers for AD determined in the same subjects (Table 3). As can be seen only few of the set variables in this study are correlated to classical AD biomarkers, mostly in the HC group (tau isoforms positively with copper, and negatively with GST and BuChE), which, in turn, vanished in SMC and MCI groups. For SMC group only selenium was related to amyloid-β (negatively) and to p-tau (positively). Finally, for MCI group only amyloid-β was negatively related to Zn.
Pearson correlation values and p-values for each variable pairs in healthy control (HC), subjective memory complaints (SMC), and mild cognitive impairment (MCI) groups
Significant Pearson correlation coefficients (r) are shadowed. * p < 0.05. ** p < 0.005.
We next performed univariate ANOVA on Se, Cu, Fe, Mn, Zn, TFER, MDA, IsoF2a, SOD, GST, GPx4, and BuChE using amyloid-β, t-tau, and p-tau as covariates. The results indicated that neither test variable was significantly influenced by classical biomarkers. Overall, these data suggest that the changes observed for the variables under study in prodromal stages have to occur before classical AD biomarkers become significantly altered.
Multivariate assessments
We next used multivariate approaches to determine what set of variables would allow a definition of the different groups. We first used PCA. The PCA results for the 11 variables are illustrated in Fig. 6. We found that two principal components explained majority of overall variance (59.64%). In the component matrix, PC1 and PC2 explain 33.43% and 26.21% of total variance, respectively. Differences between groups were mainly attributed to the variables with higher coefficients, namely, Se, Cu, Zn, and Mn for PC1 and SOD and TFER, GST, and MDA for PC2 (Fig. 5A). The analyses revealed that the alterations in the levels of these variables provide sufficient variance (F2.25 = 18.35. p = 0.000 for factor score 1 and F2.17 = 2.72. p = 0.087 for factor score 2) to allow a high degree of segregation between groups (Fig. 6B). Indeed, analysis of factor score 1 indicated that the three groups are statistically different from each other (Fig. 5C).
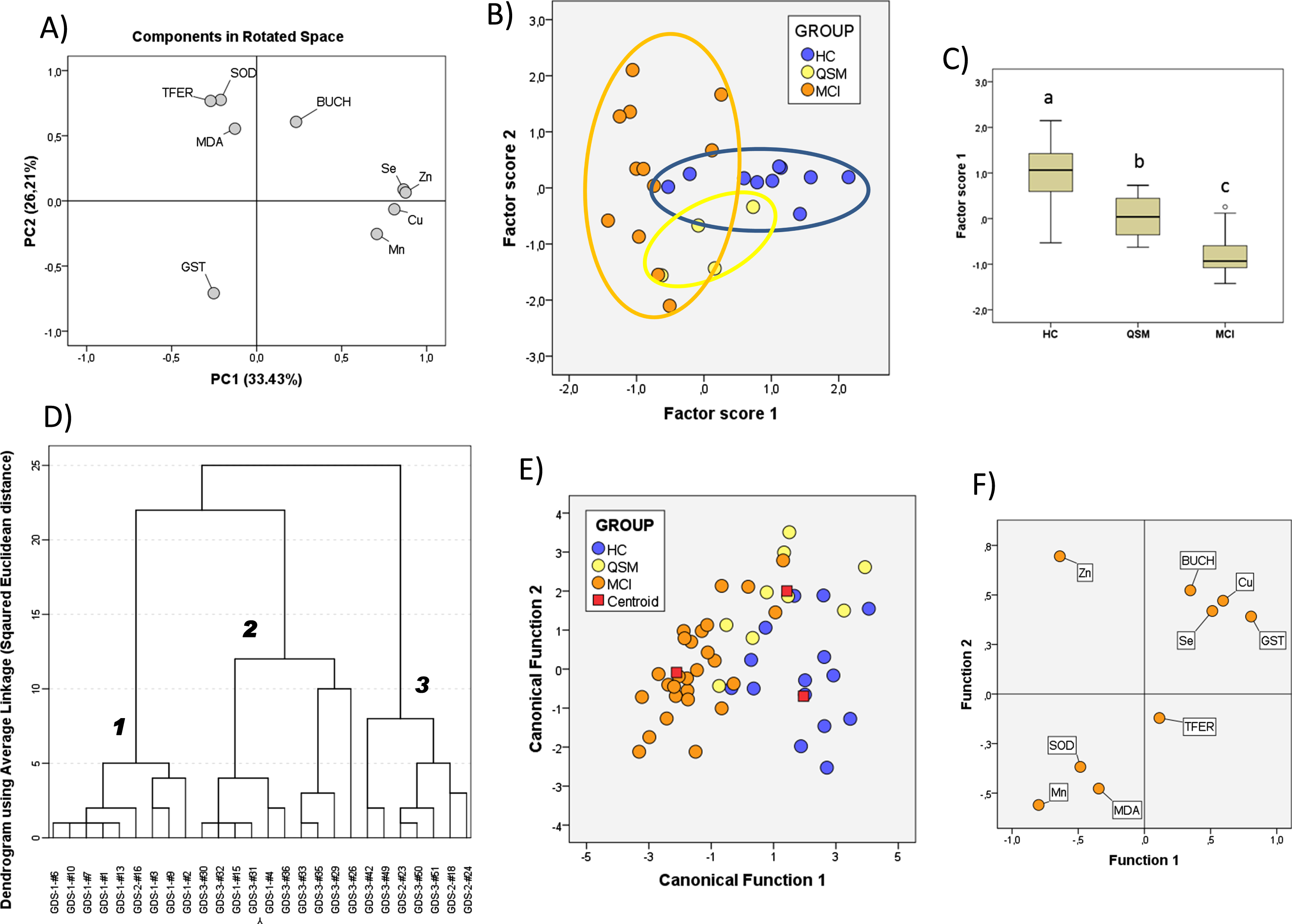
Principal component analyses of the CSF variables included in this study. A) PCA components in rotated space. B) Score plots for all subjects from HC, SMC, and MCI groups. C) Box-plot representation of factor scores from PC1. In C, different letters indicate statistically significant differences with p < 0.05 (post hoc Tukey’s HSD). D) Cluster analyses for factor scores from PCA. Bold numbers identify separated clusters. E, F) Canonical functions of CSF variables for the three groups obtained from DFA (E) and plot of matrix structure (F). For details, see the Results section.
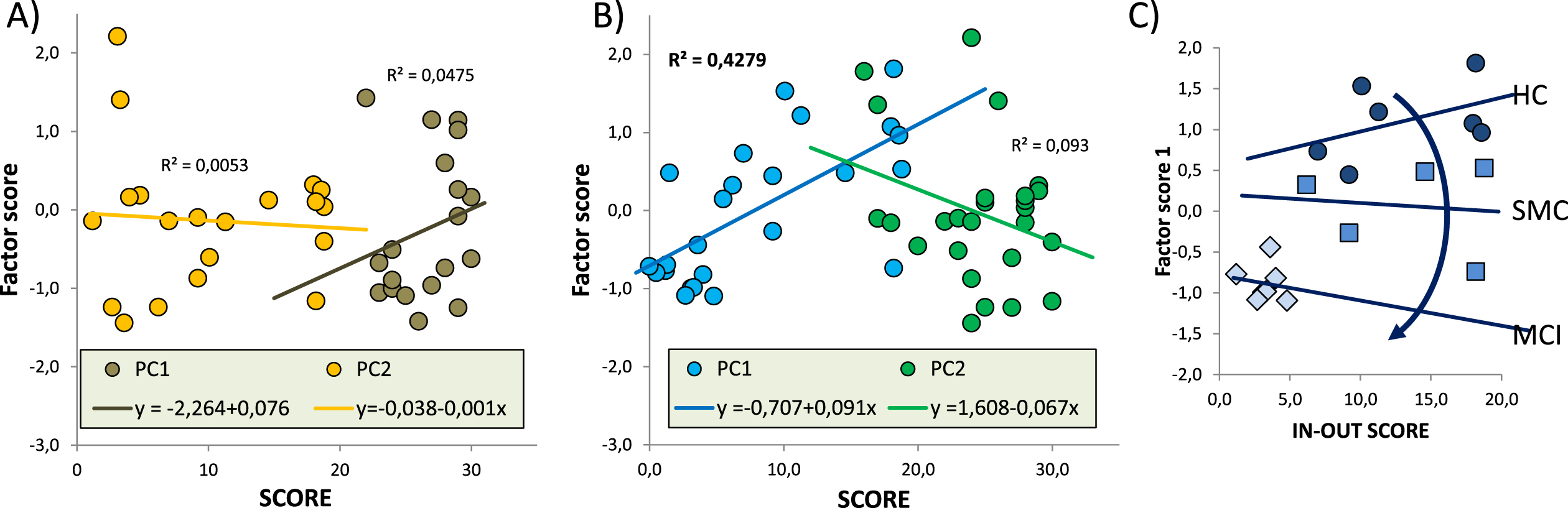
Associations of neurocognitive tests and factor scores from PCA shown in Fig. 5. A) Lineal relationships between PC1 or PC2 scores and MMSE along the whole range of data. B) Lineal relationships between PC1 or PC2 scores and IN-OUT test along the whole range of data. Regression equations are indicated in the legend box. Determination coefficients (R2) are indicated for each regression line. C) Lineal relationships between factor score 1 and IN-OUT scores in healthy controls (HC) and prodromal SMC and MCI stages.
We next used factor scores as input matrix to perform hierarchical cluster analyses in order to detect hidden groups/clusters based on inter-group similarities without a priori assumptions. As shown in Fig. 5D, three defined clusters are identified at similar Euclidean distances corresponding to HC (cluster 1, mostly GDS1), MCI (cluster 2, mostly GDS3), and cluster 3 including most SMC (GDS2) and few MCI subjects (GDS3). Thus, the two factors scores can efficiently define the group structure of the present cohort without assumptions of subject classification.
We have also applied DFA to determine the coefficient matrixes defining each group and to calculate the probabilities of each subject belonging to a particular group. Given that a number of individuals without the whole set of values in SMC was low in the SMC groups, thereby limiting the number of discriminating variables; we used the three groups of subjects replacing missing values with variable means within each group. The results are shown in Fig. 5E. The first discriminant function has a high capacity to discriminate the groups (λ (Wilks) = 0.098. χ² = 44.17 p = 0.001) and contains 82.7% of overall variance while the second canonical function represents the remaining 17.3% (λ (Wilks) = 0.523. χ 2 = 12.30. p = 0.14). According to the matrix structure coefficients (Fig. 5F), variables with highest loadings (correlations for a particular variable within the discriminant function) for the first discriminant function are Zn, Se, and Cu while for the second discriminant function, main variables were GST, SOD, and MDA, although the latter contributed to group discrimination to a lesser extent.
Relationships between neurocognitive tests and principal components
We finally used the results from PCA to evaluate potential relationships with the data of neurocognitive tests, Mini-Mental State Examination (MMSE) and In-Out test, indicated in Table 1 for each group. The results of these analyses are shown in Fig. 6. Neither PC1 nor PC2 were related to MMSE scores in the whole range of data (Fig. 6A). However, for the In-Out test, a significant lineal positive relationship was detected for PC1 (R2 = 0.428, p < 0.05), revealing that larger In-Out punctuations the higher the scores in PC1 (Fig. 6B). Further, a more detailed intra-group analysis of PC1 and In-Out test score association suggests a gradual decrease of regression coefficients (β) as the disease progresses (Fig. 6C). As computation of each factor score requires non missing values in every variable included in the principal component, the number of valid cases was considerably reduced, therefore no further statistical analyses were performed.
DISCUSSION
AD is a multifactorial disease triggered by different processes which ultimately lead to neuronal destruction and death. Numerous studies have linked uncontrolled oxidative processes, metal dyshomeostasis, and aberrant detoxifying mechanisms with the appearance of histopathological markers of the disease: SPs and neurofibrillary tangles. In this study, we have addressed the changes of different parameters, i.e., metals, markers of oxidative stress and antioxidant and detoxifying activities (as well as classical markers amyloid-β, total tau, and phosphorylated tau) in the CSF in subjects from preclinical and asymptomatic stages of AD.
There is general agreement that oxidative stress is a critical factor in the pathogenesis and progression of AD and may be associated with its earlier stage, amnestic MCI [9 , 59]. In the present study, we observed an increase in MDA concentration in the MCI group but not in the SMC stage. Such an increase was not detected for the lipoperoxide 8-iso-F2α in either stage. Paralleling these findings, we observed that extracellular SOD activity was substantially higher in the CSF of MCI patients compared to HC and SMC groups. This finding agrees with the observations showing high levels of SOD activity in the brain tissue of AD patients [14, 17], that we previously corroborated in the CSF of advanced AD patients [52]. Recently, CSF SOD1 activity has been proposed as biomarker for tau but not amyloid-induced neurodegeneration in AD [60].
Interestingly, we observed no changes in GPx4 activity between groups, though a clear trend to increase activity is observed in MCI subjects. This enzyme is the main responsible of reducing oxidized phospholipids in cell membranes, and is essential for preserving docosahexaenoic acid, the most abundant fatty acid in nerve cells [41 , 62]. This result is in line with subtle of changes in the CSF levels of complex lipoperoxide 8-iso-F2α, suggesting that the subtle changes in the levels of oxidative stress in preclinical stages are not high enough to induce uncontrolled oxidative stress and nerve cell membrane damage. In support of this, we observed that SOD and GPx4 activities are positively correlated in HC group and this relationship is retained in the MCI group, while the positive relationship 8-iso-F2α/GPx4 present in HC group disappears in MCI. However, this 8-iso-F2α/GPx4 relationship becomes negative in SMC group, meaning that the higher the activity of GPx4, the lower the concentration of 8-iso-F2α, and suggesting a role for GPx4 in early stages of cognitive decline.
Another component of glutathione antioxidant system is GST, an essential family of enzymes involved in detoxifying mechanisms associated to reactive LPO and adduct formation in different tissues and physiological conditions, including neurological disorders [31]. It has been known that GSTs can reduce lipid hydroperoxides through their Se-independent glutathione-peroxidase activity and that these enzymes can also detoxify lipid peroxidation (LPO) end-products such as HNE, one of the main end products of LPO which react with lysine and cysteine residues proteins such tau or LRP1 [18 , 64]. We provide here the first evidence that GST activity is significantly increased in the CSF of preclinical stage SMC, which suggests that GST activation is part of a very early strategy to cope with deleterious consequences of LPO generation in nerve tissue. However, different studies in brains and CSF have shown that GST activity decreases in AD patients [29, 30], which has been interpreted as secondary to oxidative modification of the enzyme. Indeed, significantly decreased activity of GST in AD brain correlates with HNE-induced modification of the enzyme [18]. Thus, it seems that a compensatory antioxidant strategy based on increasing SOD and GST activities may provide a degree of antioxidant defense in preclinical stages. Further studies will be necessary to better understanding the changes in SOD and GST activities and their relationships with molecular events underlying AD onset and progression.
Numerous studies have linked changes of brain oxidative status with trace elements [42 , 66]. Further, there exist direct evidence that dysregulation of trace metal levels in the brain associate with a range of neurological and neurodegenerative diseases from amyotrophic lateral sclerosis to AD [42 , 66–72].
In our present study in the CSF of pre-AD stages, we have observed significant changes of all metals with the sole exception of iron. Changes could be assigned to two major patterns depending on their stage of appearance and progression as 1) early-onset and stable and 2) progressive developing. In the first pattern, observed for Cu and Mn, minimal levels are detected already in the SMC group and remain without further changes in the MCI group. Bivariate analyses indicate a positive relationship between the two metals which is maintained along stages, though their relationship with lipoperoxydes varies. Thus, both Cu and Mn were unrelated to LPOs in HC but became positively correlated to MDA in the SMC group. In agreement, it is widely accepted a link between Cu and AD as postmortem studies indicate that AD brains are characterized by an excess of extracellular Cu, since it accumulates in SPs, a finding which is paralleled by decreased intracellular Cu compared to control brains [33, 69]. Similarly, reduced Mn levels have been reported in the frontal and occipital lobes of AD brains in the CSF of AD patients [67] suggesting a potentially contributory role for Mn deficiency in disease development [73]. Following the same reasoning discussed for Cu, Mn reduction in the CSF must correspond to an early pathogenic event well before SPs are formed.
The second pattern is exhibited by Se and Zn and is characterized by a progressive reduction of CSF levels and reaching a minimum in the MCI group. Most bivariate relationships between variables in the HC group disappear in this later group with the exception of Cu/Mn which does not change through stages. It is interesting to note that despite the significant reductions of Se and Cu in MCI subjects, these two metals display a very significant positive relationship. However, in the SMC group, a particular behavior is observed for Zn, which correlates positively with 8-iso-F2α, SOD, GST, and BuChE. As in the case of Cu, Zn also accumulates in SP in AD [29 , 67], where it interacts with amyloid-β promoting precipitation and giving rise to protease-resistant unstructured aggregates [74, 75] which promotes further oxidation. In addition, Zn sequestering into SP likely reduces Zn availability at the synaptic cleft, hence leading to altered neuronal signaling and synaptic plasticity, in particular in glutamate- and zinc-releasing neurons, which represent almost half of all of glutamatergic synapses [39].
Selenium is the essential metal for the synthesis of selenocysteine, an amino acid very similar to cysteine, but incorporates selenium instead of sulfur, as part of reaction center of selenoproteins, including glutathione peroxidases [76]. Different studies have shown that Se deficiency drastically reduces GPx4 enzyme activities and the cellular antioxidant capacity, usually accompanied by disruption of membrane integrity by membrane lipid oxidation [40 , 68]. A decrease in CSF levels of Se has been also observed in AD patients [67] and in the plasma and CSF of subjects suffering from MCI [77]. Further, some studies have suggested that the content of Se in the elderly is significantly reduced, and this apparently accelerates the aging process [40]. Other studies have reported a positive association between Se status and cognitive performance in older adults, and is being accepted that alterations in Se status contribute to cognitive impairment [38, 78]. Our current results expand on the importance of Se in cognitive performance and suggests that Se reduction in CSF (and concomitant decline in brain levels) occurs at initial stages of cognitive deficits, irrespective of whether it will progress to AD.
An interesting issue arising from this study is that CSF iron levels were not modified in any of the groups under study whereas CSF transferrin levels were elevated in prodromal stages SMC and MCI respect healthy controls. This finding was unexpected as much of the literature reports increased Fe levels and Fe binding proteins in the AD brain [33 , 69] though the mechanisms leading to Fe accumulation remain unclear [32, 66]. We hypothesize that increased transferrin levels in the CSF reflect global brain iron burden, and it is likely that iron elevation in specific brain areas affected during AD, and more obvious in prodromal AD, might be too confined regionally to be reflected in CSF. In this sense, TFER might be a suitable candidate for exploring preclinical AD.
Accumulation of trace metals in senile plaques have received considerable attention and the amyloid-β-[Fe, Cu, Zn] complexes either bound or acting synergistically has been considered the main source for the generation of oxidative stress in AD [37 , 79]. The question arises of whether metal accumulation is prior or subsequent to plaque formation, and whether their accumulation in brain tissue is an upstream cause or a downstream consequence of plaque formation. Studies in transgenic mice AD models have shown that metals accumulation depends on the specific metal, may occur during early stages, and independently of senile plaques formation. Indeed, using the PSAPP mouse model of amyloid plaque formation, Leskovjan and coworkers (2011) [80] obtained the time-course of Fe, Cu, and Zn contents in the cortex and hippocampus. They observed that Fe was significantly higher in the cortex than in controls at stages corresponding to the commencement of plaque formation, and that elevated iron was not associated with the amyloid plaques. Further, neither Cu nor Zn was elevated in amyloid plaques until the latest stage, when amyloid plaques are present throughout the cortex [80]. Thus, evidence of metal dyshomeostasis in prodromal stages of AD are growing steadily, and current reports, including this study, suggest that brain trace metal alterations are key players in the development of AD well beyond ROS generation from the ternary complexes [7, 32, 37].
The set of changes observed here for the different variables in the CSF along prodromal stages of AD, encouraged us to perform multivariate approaches in the pursuit for a panel of potential biomarkers at the interface of lipoxidative metabolites-trace metals-antioxidant/detoxifying enzymes. We found a high degree of segregation between HC, SMC, and MCI groups in the Factor score 1, which was positively related to Se, Zn, Cu, and Mn. Further separation was provided by incorporation of TFER and EC-SOD (both positively related) and GST activity (negatively related) to Factor score 2, respectively. Cluster analyses revealed the presence of three clusters associated to HC, MCI, and a third group including SMC+MCI subjects. Discriminant function analyses were also performed, and the outcomes were also indicative of group discrimination, being the main canonical function positively associated to GST, Cu, and Se, and negatively to Zn, Mn, and EC-SOD, hence similar to PCA results. The three multivariate methods used here (PCA, CA, and DFA) provided evidence that HC and MCI groups are sufficiently different as to allow their neat segregation, while SMC subjects appear as an intermediate group without a clear separation between the previous and next stages of the cognitive process. This inaccuracy is surely due to the low number of SMC subjects in the study, but most likely to the existence of a continuum in the transition from healthy cognition to cognitive impairment.
We have finally assessed the potential relationship of multivariate outcomes with neuropsychological evaluation of this cohort. We found that In-Out test punctuations (but not MMSE) were positively related to principal component 1 from PCA, and that a progressive change in the degree of their association was apparent in the HC ⟶ SMC ⟶ MCI transition. These results are outstanding as they indicate that CSF variables included in PC1 might predict cognitive alterations, specifically in episodic memory, and more interestingly, in predicting conversion to dementia [81].
Though promising, a major drawback of the present study is the limited size of the cohort and the uncertainty on the progression of the cases analyzed here towards more advanced stages in the AD continuum. In this sense, we have undertaken a 5-year follow-up study with the patients included in this study. Nonetheless, there is a clear need for larger cohort and multicenter studies on the association of CSF trace metals and antioxidant/detoxifying enzyme activities with the onset and early progression of AD.
Conclusions
Overall, the results in the present study indicate that prodromal stages of AD may be identified by the altered CSF values of trace metals and antioxidant and detoxifying activities, to a much larger extent than lipoxidative metabolites and even classical markers of AD-related proteinopathies. A significant outcome if this study is that combination of several of these CSF variables allows envisaging a scenario for early changes in the development of AD. Indeed, the multivariate approach of such potential biomarkers allows the definition of a molecular signature for prodromal stages of AD, in particular for subject suffering subjective memory complaints, perhaps the earliest recognizable neurocognitive trait in AD progression.
Footnotes
ACKNOWLEDGMENTS
FM-H wishes to thank Fundación La Caixa-Caja Canarias (Spain) for financing her predoctoral fellowship. We would like to thank Prof. José Luis Gomez-Ariza and Prof. Luis Galindo for providing the necessary support and equipment to carry out the metallomics experimentation. We also thank the staff at the Neurology and Orthopedic surgery services from Hospital Universitario de Gran Canaria Dr. Negrin for excellent collaboration and technical assistance.
This study was supported by grants ProID2020010075 from Agencia Canaria de Investigación, Innovación y Sociedad de la Información (ACIISI, Gobierno de Canarias, Spain) and SAF2017-84454-R (MINECO, Spain).