
Abstract
Background:
Mechanistic studies in animal models implicate a role for saturated fatty acids in neurodegeneration, but validation of this finding in human studies is still lacking.
Objective:
We investigated how cerebrospinal levels of sphingomyelins (SM) and phosphatidylcholine (PC)-containing saturated fatty acids, monounsaturated fatty acids, and polyunsaturated fatty acids associate with total tau and phosphorylated tau (p-tau).
Methods:
Cerebrospinal fluid (CSF) lipids were measured in two cohorts, a discovery and a confirmation cohort of older non-demented individuals from the University of Southern California and Huntington Medical Research Institutes cohorts. Lipid analysis was performed using hydrophilic interaction liquid chromatography, and individual PC and SM lipid species were measured using tandem mass spectrometry. In addition, CSF levels of Aβ42, total tau, and p-tau-181 were measured using an MSD multiplex assay.
Results:
The discovery cohort (n = 47) consisted of older individuals and more females compared to the confirmation cohort (n = 46). Notwithstanding the age and gender differences, and a higher p-tau, Aβ42, and LDL-cholesterol in the discovery cohort, CSF concentrations of dipalmitoyl-PC (PC32a:0) were significantly associated with p-tau in both cohorts. Similarly, total saturated PC but not mono or polyunsaturated PCs correlated with p-tau concentrations in both cohorts.
Conclusion:
Saturated PC species in CSF associate with early markers of neurodegeneration and are potential early disease progression biomarkers. We propose mechanisms by which saturated PC may promote tau hyperphosphorylation.
INTRODUCTION
Changes in the metabolism of sphingolipids and glycerophospholipids in the brain are consistently associated with preclinical Alzheimer’s disease (AD), prodromal AD, AD pathology, and tauopathies [1–5]. In addition, phosphatidylcholine (PC) and sphingomyelin (SM) lipid species are critical in forming lipid rafts, essential structural and functional domains for maintaining neuronal function [1, 6]. Tau is a microtubule-assembly associated protein that helps in cell transport, structure, and intracellular trafficking. In AD, tau protein becomes hyperphosphorylated and starts to aggregate as neurofibrillary tangles, preventing regular intracellular trafficking [7, 8]. Amyloid plaques, composed of amyloid-β peptides, and neurofibrillary tangles, composed of phosphorylated tau, are considered the hallmarks of AD pathology [9–12]. Tau phosphorylation is regulated by the balance of several kinases and phosphatases: kinases glycogen synthase kinase-3 (GSK-3), cyclin-dependent protein kinase-5 (cdk-5), extracellular regulatory kinase (ERK) 1/2, calmodulin-dependent protein kinase II (CaMKII), protein kinase A (PKA), the stress-activated c-Jun N-terminal kinase (JNK) and p38 kinase, and the phosphatases PP2A, PP2B and PP1 [13–15].
Changes in lipid rafts affect markers of AD pathology and might be reflected in the change in cerebrospinal fluid (CSF) lipids. The PCs present in the cell membrane are hydrolyzed by phospholipase A2 leading to the generation of lysophosphatidylcholine and free fatty acids. As a result, the intracellular calcium signaling via G-protein receptor (GPR132, G2A) is dysregulated, resulting in enhanced activity of CAMKII, in coordination with the ceramide–PP2A–GSK3β pathway, leading to tau hyperphosphorylation [1, 16]. Recently, saturated lipids contained in brain lipoprotein particles were shown to mediate astrocyte-induced neurotoxicity in cellular and animal models [17]. The purpose of this study was to investigate the association of CSF lipids with CSF tau phosphorylation, a marker of early AD pathology and brain tauopathies.
METHODS
Participants
All study participants provided signed informed consent and all the procedures were approved by the Ethics Committee. Cohort 1 participants were recruited between 2010 and 2016 at Huntington Medical Research Institutes (HMRI), Pasadena, CA (IRB # (HMH-99-09). Cohort 2 participants were recruited at the University of Southern California (USC) ADRC between 2016 and 2021 (USC IRB: HS-16-00888). Both cohorts had matching inclusion and exclusion criteria. Inclusion criteria were: English and Spanish-speaking men and women aged 55 and older with CDR score≤0.5 (indicating normality or mild cognitive impairment). All participants completed the National Alzheimer Coordinating Center Uniform data set (UDS 3.0) neuropsychology battery. Participants meeting mild dementia criteria (CDR = 1) [18] or clinical diagnosis of dementia were excluded. CSF and plasma were obtained from the study participants at the time of assessment. The lumbar puncture was performed the morning after an overnight fast.
Outcomes
The primary outcome was the measurement of PC and SM lipid species and their associations with AD biomarkers, specifically levels of phosphorylated tau (p-tau181) and total tau.
Measurements
APOE genotyping was assessed using qPCR with isoform-specific primers as described [19]. CSF levels of total tau and ptau (p-tau181) were measured using MSD multiplex assay as previously described [20]. For phospholipid measurements, a 400μL CSF aliquot was transferred into a reaction vial containing a 25μL aliquot of internal standard mix [21] [2 ng PC (11:0/11:0), 2 ng D4-PAF (16:0), 2 ng LPC(11:0), and 2 ng D4-LPAF(16:0), 2 ng D3-LPC (16:0))], 100μL PBS, and 475μL of water containing 1M NaCl and 0.9% formic acid. Phospholipids were extracted using a modified Bligh & Dyer procedure using the Gerstel Multipurpose Sampler Robotic machine [22]. In short, 2 mL methanol containing 0.2 mg/mL BHT and 1 mL chloroform was added to the CSF to create a mono-phase at room temperature. Following vigorous shaking for 10 min, 2 mL chloroform and 1 mL water containing 1 M NaCl and 0.9% formic acid were added to separate a lipid-rich chloroform layer, aspirated after shaking and centrifugation for 5 min, and transferred to clean reaction vials. This process was repeated once more, collecting all of the lipid-rich chloroform in one vial. 1.2 mL of HPLC-grade water was then added to the extracted CSF to remove any remaining salt. The lipid-rich chloroform layer was dried under a stream of N2 then resuspended in 1 mL of 1:1 CHCl3: MeOH (v/v). All 1 mL aliquots of resuspended CSF were transferred to VerexTM vials, dried under a stream of N2, and reconstituted in HPLC solvent for LC-MS/MS analyses. Phospholipid analysis was performed using hydrophilic interaction liquid chromatography (HILIC) with a TSK-Gel Amide-80 Column (2.0×150 mm) and positive electrospray ionization tandem mass spectrometry using a triple quadrupole mass spectrometer (TSQ Classic, Thermo Fisher Scientific, San Jose, CA), as described previously [23].
Data extraction
We obtained the masses of PC species containing saturated, monounsaturated, and polyunsaturated species from the LipidMaps database and the quantified PC and SM species were extracted using the Thermo Scientific Quant algorithm from the parent ion scan (m/z=184).
Statistical analysis
Data analyses
Using volcano plots, PC and SM levels were analyzed in association with p-tau and total tau measurements in the discovery and confirmation cohorts. Variables were compared using Fisher’s exact test and Wilcoxon’s two-sample test. p-values below 0.05 (two-sided) were considered statistically significant for the primary outcome. All analyses used R (https://www.R-project.org/).
RESULTS
Subject demographics
Subject demographics are shown in Table 1, stratified by cohort. All participants had CDR≤0.5 and were over 55 years old. Most participants were cognitively unimpaired and non-Hispanic white. There were no significant differences in total tau levels between cohorts. In the discovery group (n = 46), the majority of participants were female, while the confirmation cohort (n = 47) was predominantly male (Table 1). The cohorts differed significantly in age, sex, CSF levels of Aβ42, p-tau, and LDL cholesterol (Table 1). The discovery cohort had greater CSF p-Tau levels (Table 1).
CSF Demographic and Clinical Characteristics by Cohort
*Cut-off levels for Aβ42 + (< 190 pg/mL) and ptau + (> 78 pg/mL) were chosen based on increased AD risk [53]. 1Mean (SD); n/N (%). 2Wilcoxon rank sum test; Pearson’s Chi-squared test; Fisher’s exact test; Wilcoxon rank sum exact test
Individual lipid species in CSF
Individual PC and SM lipid species were measured in CSF using LC-MS/MS. All lipid species were analyzed against total tau and p-tau levels by cohort using volcano plots. Although 37 lipid species were significantly associated with p-tau in the discovery group, only 4 lipid species, PC32a:0, PC34a.1, PC34e.4/PC34p.3, and PC36e.6/36p.5 were significantly associated with p-tau in the confirmation cohort saturated lipid species (Fig. 1). Among those 4, PC32a:0, a PC lipid carrying two palmitic acids, was strongly associated with p-tau both cohorts (Fig. 2). As shown in Supplementary Figure 1, 44 lipid species were significantly associated with total tau in the discovery cohort. However, no lipid species were significantly associated with total tau in the confirmation cohort.
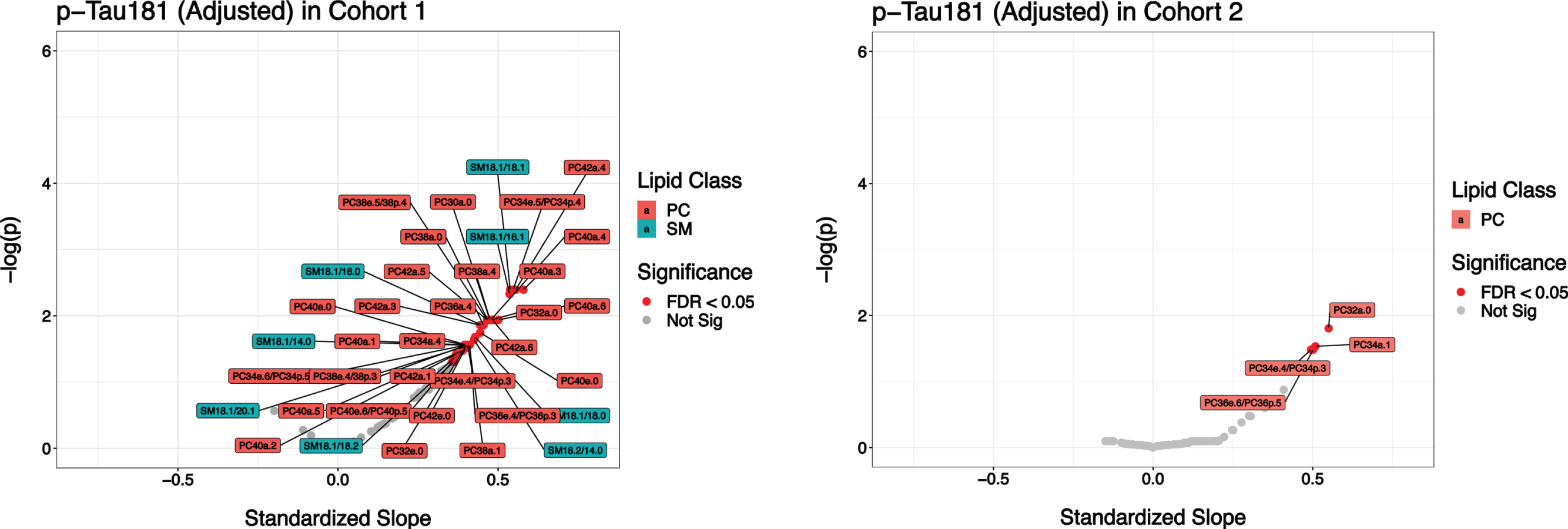
Association of CSF lipids with p-Tau181. Volcano plots showing the association of CSF lipids with p-tau181 in both the discovery and confirmation cohorts. In the discovery group, 37 lipid species were found to be significantly associated with p-Tau. However, in the confirmation cohort, only 4 lipid species, PC32a:0, PC34a.1, PC34e.4/PC34p.3, and PC36e.6/PC, 36p.5/PC were found to be significantly associated with p-Tau. The adjusted variables used in the linear regression model are age, sex, APOE4 status, and years of education.

PC32a:0 versus P-Tau. PC32a:0 in CSF, a PC lipid carrying two palmitic acids, was significantly positively associated with p-tau in both the discovery and confirmation cohorts.
Grouped lipid species in CSF
Individual PC species were grouped by saturation levels, or the number of double bonds, and analyzed for an association with CSF p-tau. In both discovery and confirmation cohorts, saturated PC lipids, or PC lipids with no double bonds known to be more rigid and supportive, were significantly positively associated with p-tau levels (Fig. 3). Monounsaturated PC lipids, or PC lipids with one double bond, were only significantly associated with p-tau in the confirmation cohort but not the discovery cohort (Supplementary Figure 2). Similarly, polyunsaturated PC lipids, or PC lipids with two or more double bonds, including PC lipids containing omega-3 and omega-6 fatty acids, had a significant positive association with p-tau in the discovery cohort but had no significant associations in the confirmation cohort (Supplementary Figure 2). One participant from the confirmation cohort (cohort 2) had a measurement of CSF p-tau greater than two standard deviations from the sample mean. Exclusion of this data point did not influence the observed associations (Supplementary Figure 3).

Total Saturated PC versus P-Tau. Saturated PC lipids, or PC lipids with no double bonds, were significantly positively associated with p-tau levels in both discovery and confirmation cohorts.
DISCUSSION
This report identifies a significant association between saturated PCs and p-tau, and underscores a role for the accumulation of brain saturated PC (i.e., PC 32a:0 species) in the pathogenesis of tauopathies. Previous research showed that saturated fats in rats affected hippocampal morphology and resulted in profound memory impairment through the neuronal death of the hippocampus due to the accumulation of hyperphosphorylated tau protein, NFTs, and amyloid plaques [24]. Therefore, to further investigate PC involvement in AD and p-tau, a robust neuronal cell loss biomarker, PC32a:0, and its association with p-tau in both cohorts were examined. PC32:0 or 1,2-dipalmitoyl-sn-glycero-3-phosphocholine is a PC containing two saturated (C16:0, palmitic acid) fatty acid moieties. While we are not aware of the cerebral function of PC32:0 in humans, PLA2 activity on PC32:0 releases palmitic acid that has several important functions. Palmitic acid influences membrane fluidity, cell growth, energy regulation, and is involved in signaling, and inflammation [25]. Post translational modification of receptors involving palmitoylation affects their internalization, localization, and influences other PTMs such as phosphorylation [26]. Diets rich in palmitic acid influence proteins (for example, alpha-synuclein) and neurotransmitters associated with changes in neurodegenerative risks [27]. Imaging data of rat brain show PC32:0 abundance in gray matter as compared with white matter, most strikingly in areas showing high plasticity [28]. The dominance of PC versus SM may reflect the structure and physical properties of membranes containing p-tau. Future studies are needed to demonstrate how lipid compositional differences mediate the association with p-tau.
Accumulation of saturated fat in the muscle and the liver plays a major role in the development of peripheral insulin resistance [29]. Here, we speculate that a similar process of saturated fat accumulation occurs in the brain thereby driving a phenotype of tau phosphorylation (Fig. 4). We propose that the hyperphosphorylation of tau in the presence of saturated PC may happen through one or more of three pathways. The first pathway involves PC-phospholipase C (PC-PLC) cleaving saturated PC into diacylglycerol (DAG). DAG temporal accumulation leads to an increase in protein kinase C (PKC) levels [30–32] and a decrease in insulin receptors (IR) [31]. The decrease of IR impairs the PI3K/AKT pathway [33, 34] leading to the hyperphosphorylation of p-tau [35] through phosphorylation of glycogen synthase kinase 3B (GSK-3B) [31, 37]. The second pathway relies on the preexisting relationship of mitochondrial dysfunction, reactive oxygen species (ROS) production, inflammation, and AD. Saturated fatty acids contribute to mitochondrial dysfunction [38] by enhancing oxidative stress, depolarizing mitochondrial inner membrane, causing calcium overload, and sarcoplasmic reticulum calcium leak [39]. When PC-PLC cleaves saturated PC into DAG, DAG and the saturated PC exacerbate mitochondrial dysfunction, thereby increasing ROS production and inflammation, leading to the hyperphosphorylation of tau [40]. A third pathway may involve lipid raft formation. PCs and SM lipid species are critical in forming lipid rafts, essential structural and functional domains for maintaining neuronal function [1, 6]. PCs with saturated and unsaturated long-chain fatty acyl groups positively influence the activity of the gamma-secretase enzyme by modulating cell membrane thickness and the lipid microenvironment of the enzyme [1, 41]. A significant amount of neuronal membrane reorganization takes place as part of memory encoding and neuroplasticity, particularly in the hippocampus [42]. This process is influenced by slight variations in membrane lipid composition, and can predispose to AD pathology [43]. Cholesterol and saturated fatty acids may promote the formation of Aβ42 through the complementary APP intracellular domain, causing excess tau phosphorylation [44]. In addition, the generation of lysophosphatidylcholine from membrane PC by both cytosolic phospholipase A2 Group IVA (PLA2G4A) in Land’s cycle [1, 45] as well as the secretory phospholipase A2 group IIA (PLA2G2A) can lead to dysregulation of intracellular calcium signaling in a G-protein receptor (GPR132, G2A) coupled manner. Dysregulated Ca2+ signaling can result in enhanced activity of CAMKII. In coordination with the ceramide–PP2A–GSK3β pathway, this enhanced activity results in tau hyperphosphorylation, leading to the generation of paired helical filaments and enhanced neurofibrillary tangle formation [1, 16] which contributes to AD.

Illustration showing different pathways that lead to hyperphosphorylation of tau. Saturated PC species hydrolyzed by lipases, PLC, or PLA2 release saturated DAG and saturated free fatty acids. DAG signaling involving PKC activation and insulin resistance may result in tau hyperphosphorylation. Similarly, saturated free fatty acids released by PLA2 activation are involved in oxidative stress and tau hyperphosphorylation.
Brain lipid composition is affected by the composition of the diet. C16:0 (palmitic acid) is the main product of mammalian fatty acid synthase, and readily enters into the brain after dietary consumption in it’s unesterified free fatty acid form [46]. A change in Western dietary patterns is associated with an increase in saturated fat consumption, contributing to the greater prevalence of metabolic syndrome [47]. Preclinical studies show that rodents fed on high saturated fat diets have impaired memory and learning [48] and altered dendritic growth [49]. High-fat diets also induce signaling pathways that promote tau pathology [50], increased tau expression, and tau phosphorylation [51, 52]. Thus, it is possible that dietary saturated fatty acids may increase PC32:0 and contribute to changes in tau phosphorylation observed in our study.
The main limitation of this study was the compositional heterogeneity across the two cohorts manifested by the differences in AD biomarkers in cohort 1 compared to cohort 2. In cohort 1, there was a higher level of p-tau in the CSF (71.40 pg/ml) compared to cohort 2 (59.70 pg/ml), but a lower percentage of amyloid positive individuals (17%) compared to cohort 2 (59%). These cohort differences may help explain the differences in lipid associations observed here with tau phosphorylation between AD and non-AD pathways, as several classes of lipids were associated with ptau in cohort 1, with only saturated PC lipids having an association in both. These data suggest that dipalmitoyl-PC is associated with AD pathology via tau phosphorylation independent of amyloid biomarker status. Our cross-sectional study does not lend itself to determining whether altered saturated lipids result in tau phosphorylation or vice versa. Although we show an association between PC32a:0 content and tau phosphorylation, studies regulating saturated fatty acids will be required to address this fundamental question. For example, dietary modification studies show that saturated fat induces signaling pathways that promote tau pathology [50–52]. Thus, it is likely that fatty acid saturation precedes higher tau phosphorylation. We excluded participants with dementia as it will be difficult to disentangle the effect of AD pathology and CSF lipids, where greater pathology (for example, tau phosphorylation) may be present independent of lipid saturation or composition status.
In summary, we identified an association between saturated phospholipids and p-tau in CSF, providing insights into early processes that drive neurodegeneration. Understanding the mechanisms that promote the accumulation of saturated fats in the brain is relevant to the eventual therapeutic management of brain tauopathies.