
Abstract
DAPI is conventionally used as a nuclear stain for cells in culture or tissue. Here we demonstrate that it binds specifically to the β-sheet core of amyloid-β plaques but not diffuse amyloid-β at the plaque periphery. The specific DAPI induced blue fluorescence is much stronger than amyloid plaque autofluorescence. DAPI staining of fibrillar amyloid deposit may yield a misleading impression of damaged or dying cells. On the other hand, it provides a handy and low-cost means of staining compact amyloid plaques together with cell nuclei in double or triple immunofluorescent studies.
INTRODUCTION
DAPI (4’,6-diamidino-2-phenylindole) is a lipid soluble fluorochrome which has high affinity to DNA and a lower affinity to RNA. It binds to the minor groove of the DNA double-strand that is rich in A-T nucleotide pairs, while in the case of RNA it binds to an A-U sequence in the single nucleic acid strand [1, 2]. Typical use of DAPI is visualization of cell nuclei of cultured cells or tissue sections, being present in the culture/mounting medium.
We made a perplexing observation that a substantial proportion of amyloid-β (Aβ) plaque cores in the 5xFAD transgenic mouse, carrying mutation human APP and PSEN1 genes, stain positive for DAPI. A quick literature search brought up two papers from recent years with a similar observation [3, 4]. Two kinds of explanation were offered: first, DAPI binding to released chromatin from the nuclei of dying cells in or around the amyloid plaque [3], and second, plaque core autofluorescence [4]. Indeed, both DAPI and amyloid plaque core have the strongest autofluorescence in the blue range [5]. In addition, a recent protein chemistry study reported that various amyloid fibrils have a high affinity to DAPI, in some cases even higher than to nucleic acids [6]. Since the interpretation of DAPI positivity in amyloid plaques plays a crucial role in our understanding of amyloid plaque formation, we set out a series of histochemical experiments to test these three explanations.
METHODS
Animals
Male and female mice of the 5xFAD line (Jackson Laboratory, Stock #34840-JAX) that carries two APP and three PSEN1 mutations were sacrificed at 3 months of age. In addition, mice carrying mutated human APPswe and PSEN1dE9 transgenes (from here on APP/PS1 mice) [7] were crossbred with CX3CR1-GFP mice (Jackson Laboratory, Bar Harbor, ME, USA, Stock #005582) to obtain mice with amyloid plaques surrounded by GFP expressing microglia. Female mice sacrificed at 13 months of age were used. Mice were kept in individually ventilated cages in a controlled environment with 12/12 h light/dark cycle and ad libitum access to water and feed. Under deep Equithesin anesthesia, the mice were perfused transcardially with saline to rinse blood from brain vessels. The brains were fixed with 4% PFA perfusion or immersion fixation (4 h), dehydrated in 30% sucrose overnight and stored in a cryoprotectant at –20°C until cut into 35μm or 100μm coronal sections with a freezing microtome. All experiments were conducted in accordance with the EU directive 2010/63/EU for animal well-being and approved by the Animal Experiment Board of Finland.
Human samples
Paraffin-embedded, 4% PFA fixed, frontal cortical sections (5μm) of a 76-year-old female Alzheimer’s disease patient (Alzheimer’s Disease Neuropathological Change/Braak VI) were obtained from the tissue archives of Department of Pathology, Kuopio University Hospital. The use of the human brain samples was approved by the Research Ethics Committee of the Northern Savo Hospital District.
Immunofluorescence
The 35μm sections of 5xFAD mice brains were left overnight a Na3PO4 (0.1 M, pH 7.6) buffer and then the sections were rinsed three times for 5 min in the buffer. The native sections were mounted on gelatin coated glass slides using Vectashield mounting medium with or without DAPI (Vector Laboratories, Burlingame, CA, USA). For amyloid immunostaining, the sections were pretreated with citrate buffer (0.05 M, pH 6.0) at 85°C for 30 min for antigen retrieval. Thereafter, the sections were cooled down in Na3PO4 buffer followed by three washing steps of 5 min in the buffer. Sections were incubated overnight at room temperature with primary antibody MoAb-2 (1:5000, Biosensis). MoAb-2 is a mouse monoclonal antibody that recognizes residues 4–10 of human Aβ. It shows aggregated, oligomeric and fibrillar forms of Aβ42, and aggregated Aβ40. Three rinse steps of 5 min with TBS-T (0.5 M, pH 8.6) were done to wash the primary antibody. A secondary antibody coupled to Alexa Fluor 488 (1:500, ThermoFisher) was then added in a TBS-T solution and sections were incubated for 2 h. Again, sections were washed three times for 5 min in TBS-T. The sections were then placed in Na3PO4 buffer and mounted on gelatin coated glass slides using Vectashield mounting medium with DAPI (Vector Laboratories).
The 100μm sections of APP/PS1+ GFP+ mice were stained by rabbit monoclonal anti-human amyloid-β (D54D2, 1:10000 for mouse sections, 1:100 for human sections, Cell Signalling Technology) followed by secondary goat anti-rabbit antibody coupled with Alexa Fluor 594 (1:400). Sections were mounted on the glass with VectaShield with DAPI (Vector Laboratories). As a control for the specificity of DAPI in mounting medium, we stained sections from one animal with DAPI (1:10,000, 20 min) in TBST pH 7.6 and mounted them with VectaShield without DAPI (Vector Laboratories). For the Thioflavin-S and DAPI staining, 35μm sections of APP/PS1+ GFP- were first mounted on glass with VectaShield with DAPI. After imaging mounting medium was rinsed off with phosphate buffer pH = 7.6 and stained with Thioflavin-S. Finally, VectaShield with DAPI was used for mounting.
Image analysis
The 35μm sections were visualized first with fluorescence microscope Zeiss Axio Imager 2 (Zeiss, Gottingen, Germany) or confocal microscope Zeiss LSM 880 with Airyscan detector (Zeiss, Gottingen, Germany), both cases with a 63x oil-immersions lens. Image processing and 3D rendering of the confocal images of 100μm sections was performed in ZEN 3.3 and 3.2
RESULTS
To assess whether autofluorescence can explain the occasionally visible blue Aβ plaque core in DAPI stained sections, we analyzed the intensity of blue autofluorescence of amyloid plaques of 5xFAD mice with different exposure times. As seen in Fig. 1A, dense core amyloid plaques can be detected with a fluorescent microscope as blue fluorescence with a long exposure time (144 ms). However, with a regular exposure (128 ms) that does not show background, the plaques are barely visible (Fig. 1B). The peak fluorescence is within the blue range and not visible in green (Fig. 1C). Furthermore, native fluorescence of amyloid plaque cores was an order of magnitude stronger with DAPI in the mounting medium (Fig. 1E) than in the absence of DAPI (Fig. 1F). As an additional control to exclude the possibility that DAPI+ signal of amyloid plaque cores results from autofluorescence, we pretreated the sections with 0.3% Sudan Black to remove lipofuscin-related autofluorescence. The pretreatment effectively reduced the background fluorescence and attenuated but did not remove the DAPI+ signal of compact plaques (Fig. 1G,H).
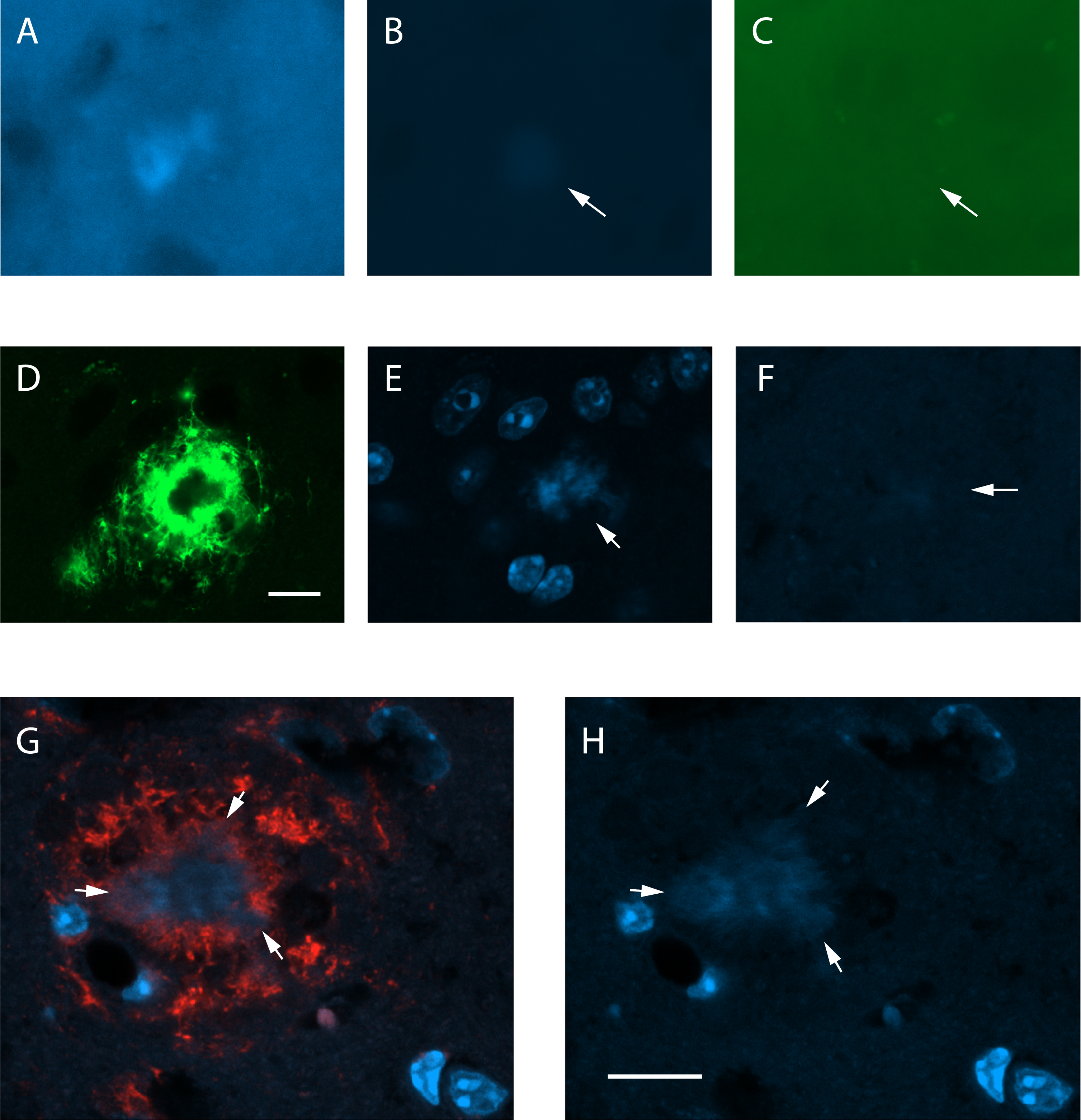
Autofluorescence of dense-core amyloid-β plaques in a 5.5-month-old 5xFAD mouse. A) A amyloid-β plaque-like deposit from a non-stained brain section as seen in the blue channel (excitation 365 nm, emission band-pass 445/50 nm) of a fluorescent microscope at the LED exposure time of 144 ms. B) The same deposit is barely seen at a lower LED exposure time (128 ms) used for sections with DAPI. dC) It is totally absent in the green channel (excitation 475/30 nm, emission band-pass 550/100 nm) at the same lower exposure time. Arrows point to the deposit. D) A confocal microscope image showing a dense-core amyloid-β plaque stained with the antibody MoAb-2 against Aβ. The plaque core remains unstained. The laser power for the green channel was 0.2%. E) DAPI staining of the same plaque at a laser power of 0.4% for the DAPI channel (358 nm). The plaque core appears DAPI positive (arrow). F) Another amyloid-β plaque of the same mouse in a non-stained section. The blue channel at 2% laser power barely shows the plaque core (arrow). G) A compact plaque double-stained with anti-Aβ antibody W02 (red) and DAPI with 0.3% Sudan Black pretreatment to remove autofluorescence. H) The compact amyloid-β (arrows) remains DAPI positive despite the pretreatment. Scale bar 10μm in A –F, 20μm in G-H.
To further evaluate the source of the DAPI+ signal we sampled high-resolution confocal images of larger amyloid plaques in 100μm sections of 13-month-old APP/PS1+ GFP+ mice and rendered 3D images offline. The GFP signal in the microglia allowed us to identify chromatin structure of neurons versus microglia around the amyloid plaques and compare it to the pattern of DAPI+ plaque cores. We observed that the plaque core in DAPI staining had a fibrillar structure with no resemblance to the chromatin pattern of the cell nuclei (Fig. 2AB). This finding was compatible with the notion that DAPI stains fibrillar amyloid.
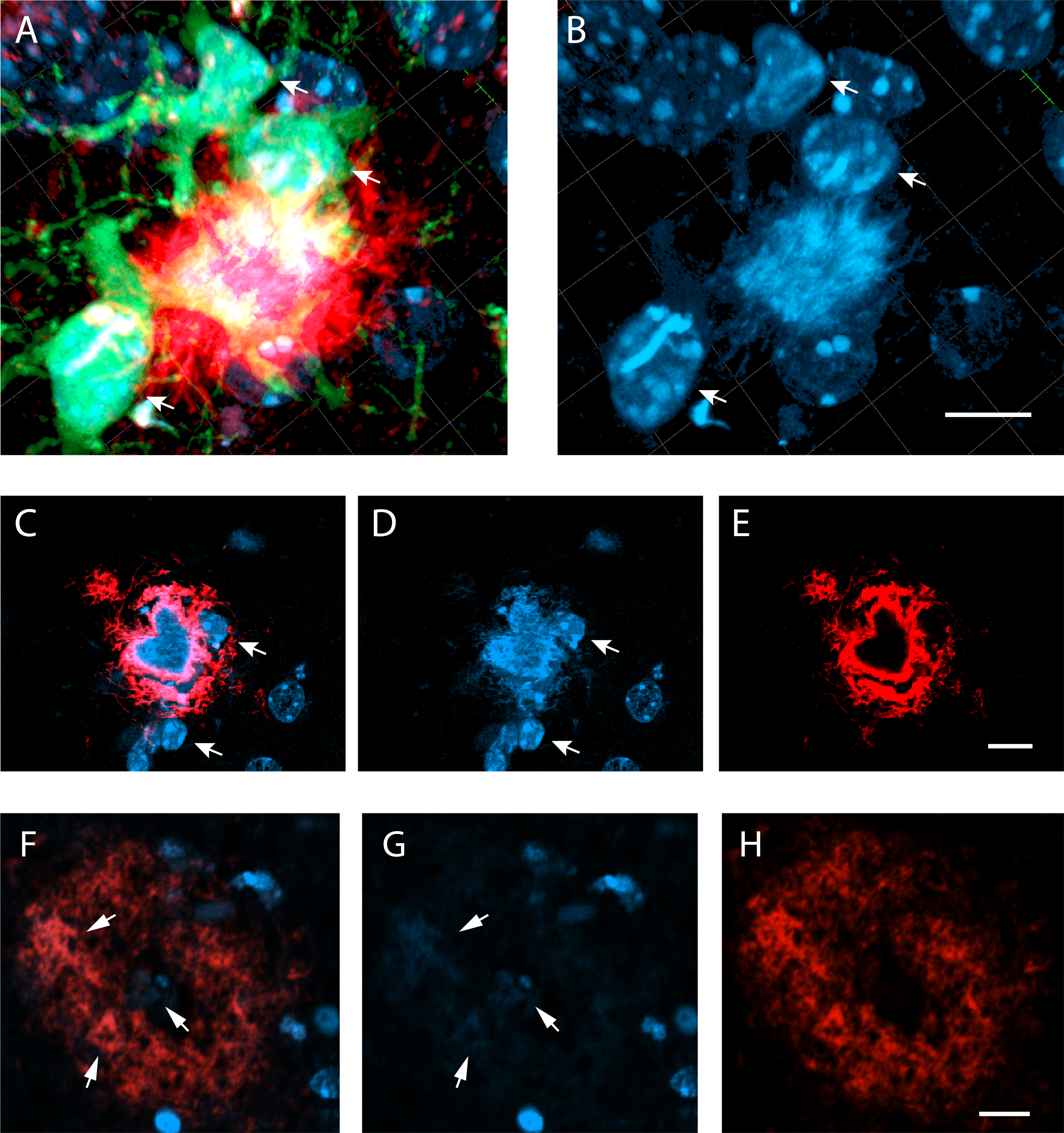
A) A confocal image (merged z-stack) on an amyloid-β plaque stained with anti-Aβ D54D2 (red) and surrounded by GFP+ microglia (green). B) The DAPI channel reveals the nuclei of the microglia (arrows) and some nuclei of GFP- cells, most likely neurons. The amyloid-β plaque core stands out by its hairy texture. C) Another compact amyloid-β plaque stained with D54D2 with DAPI in the mounting medium. A single z-plane is shown. The heart-shape plaque core is D54D2 negative and distinct from the surrounding microglia nuclei (arrows) by its texture. DAPI does not stain the diffuse Aβ in the plaque periphery or the small satellite. D) Blue channel alone, E) red channel alone. F) A compact amyloid-β plaque in the frontal cortex of an Alzheimer’s patient brain (ADNC VI) stained with anti-Aβ D54D2 (red) and DAPI. G) Compact amyloid regions are faintly DAPI positive (arrows), H) Red channel alone. Scale bar in all images 10μm.
To further characterize the nature of DAPI+ material in the amyloid plaque, we did double staining for DAPI and D54D2 anti-human Aβ antibody (1:10,000, Cell Signaling Technology). This double staining revealed that some of the larger plaques lacked D54D2 staining in the plaque core, giving them a donut-like appearance (Fig. 2CE), while the hollow center stained exclusively with DAPI and had a fibril-like texture (Fig. 2D). However, the DAPI signal also partially overlapped with D54D2 (Fig. 2CE), showing that DAPI can also stain less condensed Aβ fibrils but not the diffuse Aβ on the plaque surface. To test whether DAPI also stains compact plaques in human Alzheimer’s disease brains, we double-stained paraffin-embedded 5μm cortical sections of an Alzheimer’s disease patient brain for D54D2 (1:100) and DAPI. In compact plaques, identified by a donut shape in D54D2 staining and thioflavin-S positive cores, the core and denser D54D2+ regions showed also DAPI positive fibrillar texture (Fig. 2F-H). The DAPI intensity was an order of magnitude fainter than in the compact plaques of 5xFAD mice, which can at least partly be ascribed to the section thickness (mouse 35μm, human 5μm).
Finally, to verify the specific staining of amyloid fibrils by DAPI, we compared its staining pattern with the widely used nonimmunologic Thioflavin-S staining for dense core amyloid plaques. First, we stained a 35μm section of a 13-month-old APP/PS1+ GFP- mouse with Thioflavin-S alone. Thioflavin-S stained the core of the plaques, which was visible equally with green (Fig. 3A) and blue (Fig. 3B) fluorescence. Then we stained other sections with DAPI in the mounting medium and imaged one dense core plaque in the hippocampus with defined landmarks such as the granule cell layer and blood vessels (there are only few plaques but occasionally large ones in APP/PS1 mice at this age) (Fig. 3C). Thereafter, we removed the cover slip, flushed the mounting medium away and performed Thioflavin-S staining with DAPI in the mounting medium of the same section. Then we identified and imaged the same plaque that was previously stained only with DAPI (Fig. 3DE) and finally compared the staining pattern with the original DAPI only staining (Fig. 3F). Again, the Thioflavin-S staining looked identical in the green and blue channels and overlapped with the DAPI staining.
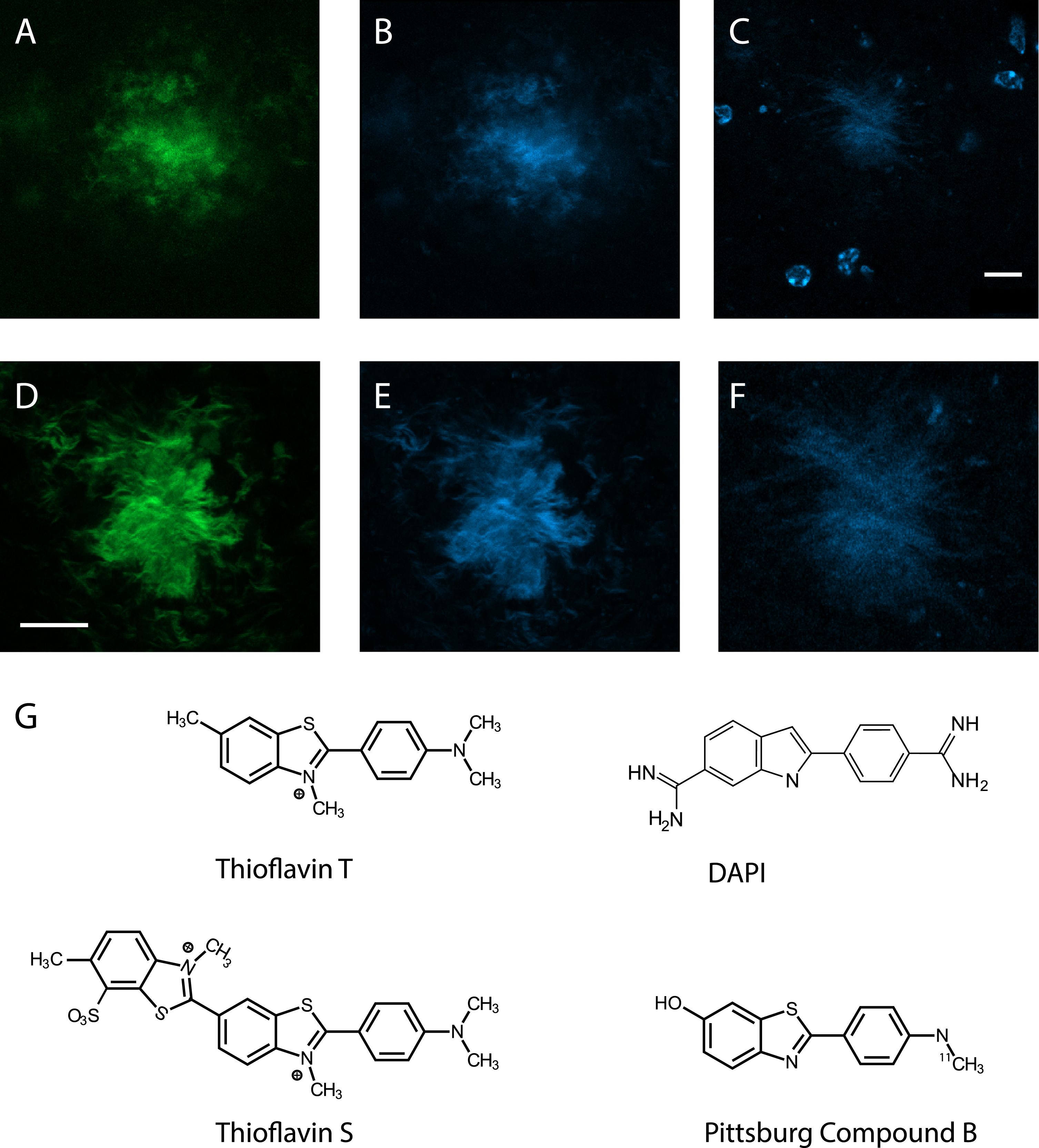
DAPI and Thioflavin-S both stain fibrillar Aβ in the amyloid-β plaque core. A) A amyloid-β plaque stained with Thioflavin-S only, green channel. B) The same plaque but blue channel, showing identical pattern. C) A amyloid-β plaque surrounded by microglia, stained with DAPI in the mounting medium. Note the strongly stained nuclear chromatin of the microglia. D) The same plaque as in C after washing the mounting medium and re-staining with DAPI+ Thioflavin-S, the green channel. E) The same as in D but blue channel. F) A close-up of the DAPI only stained plaque in C. All images are taken with a confocal microscope. Scale bar = 10μm. G) The chemical structure of DAPI resembles that of Thioflavin-T and its derivatives, Thioflavin-S and Pittsburg Compound B.
DISCUSSION
The present findings demonstrate that DAPI shares similar staining properties for Aβ plaques as Thioflavin-T and -S and their derivatives such as Pittsburg Compound B. It stains the plaque core that consists of Aβ in the β-sheet structure but not diffuse Aβ in the plaque periphery. This finding is consistent with the recently reported high affinity of DAPI to fibrillar protein aggregates [6]. Overlapping staining between Thioflavins and DAPI is not surprising considering the basic structural similarity between these compounds (Fig. 3G). However, DAPI is an indole while Thioflavin-T derivatives are benzothiazoles and they have amino groups instead of methyl groups at their ends. These differences may account for the lower staining intensity of DAPI.
The strong affinity of DAPI to fibrillar Aβ provides the best explanation to observations that some of the senile plaques have DAPI+ cores [3, 4]. Autofluorescence of Aβ plaques is well established [8], but we found that it is an order of magnitude weaker than the fluorescence induced by added DAPI in the mounting medium. However, in the fixed brain tissue the DAPI fluorescence of the amyloid plaque cores did not exceed or even reach the level of nuclear staining as was reported to be the case in a solution [6].
DAPI binding to amyloid plaque cores is a possible confound in histochemical studies assessing Aβ plaque toxicity and related cellular damage, and therefore worth knowing in the Alzheimer’s disease research community. On the other hand, DAPI provides a handy and low-cost method of visualization of Aβ plaques in double or triple fluorescent staining. DAPI staining is tissue friendly, uses the often-unutilized blue channel, and in many cases provides a two-in-one staining for the nuclear chromatin structure and amyloid plaque core.
Footnotes
ACKNOWLEDGMENTS
We thank Pasi O. Miettinen from A. I. Virtanen Institute for technical assistance in histological staining. We also thank Janne Capra from Cell and Tissue Imaging Unit at UEF for technical assistance with ZEISS. The study was supported by Academy of Finland. M.G. salary was covered by the Doctoral Programme for Molecular Medicine at the University of Eastern Finland.