
Abstract
Background:
Interleukin-10 (IL-10) is a classic anti-inflammatory cytokine that exerts its effects via the receptor complexes IL-10RA and IL-10RB. Loss of IL-10RB results in many diseases. Moreover, IL-10RB is closely associated with neuronal survival and synaptic formation. However, the regulation of IL-10RB gene expression remains elusive.
Objective:
To investigate whether the expression of IL-10RB gene is increased in brain of Alzheimer’s disease (AD) and its transcriptional regulation.
Methods:
We examined the gene expression of AD patient brain from public database and detected the protein expression of AD model mouse brain by western blot. We constructed a variety of reporter gene plasmids with different lengths or mutation sites, tested the promoter activity and defined the functional region of the promoter with the luciferase reporter assay. The protein-DNA binding between transcription factors and the promoter was analyzed using chromatin immunoprecipitation (ChIP) and electrophoretic mobility shift assay (EMSA).
Results:
We found that the IL-10RB is elevated in the brain of AD patient and AD model mice. The minimal promoter of the IL-10RB gene is located in the –90 to +51 bp region (relative to the transcriptional start site) and is sufficient for high-level expression of the IL-10RB gene. Transcription factors Sp8 and Sp9 bind to the IL-10RB promoter in vitro. The overexpression or knockdown of Sp8 and Sp9 affected the IL-10RB promoter activity and its gene expression.
Conclusion:
Our study functionally characterized the promoter of the IL-10RB gene and demonstrated that Sp8 and Sp9 regulated its expression.
INTRODUCTION
Alzheimer’s disease (AD) is a progressive neurodegenerative disease characterized by extracellular deposition of amyloid-β (Aβ) plaques, intracellular hyperphosphorylation of tau protein, activation of microglia, and neuronal cell death [1]. The amyloid cascade hypothesis has been widely accepted as the centerpiece of AD research. As a result, amyloid has served as a major therapeutic target, although the achieved therapeutic effect has not been satisfactory [2, 3]. The amyloid cascade-inflammatory hypothesis is a new and promising hypothesis [4], which suggests that excessive deposition of Aβ and hyperphosphorylation of tau protein in the brain interfere with neuronal function and trigger an inflammatory microglial response. Activated microglia release numerous inflammatory cytokines to further induce Aβ deposition and finally form a vicious cycle [5–8]. In addition, multiple studies have suggested that nonsteroidal anti-inflammatory drugs (NSAIDs) (the most widely used anti-inflammatory drugs) have a substantial sparing effect on AD, which further supports the neuroinflammatory hypothesis for AD pathogenesis [9].
IL-10, as a canonical anti-inflammatory cytokine, inhibits the synthesis and release of proinflammatory cytokines, including tumor necrosis factor (TNF)-α, IL-1β, IL-6, IL-8, and IL-12 [10]. In the central nervous system, IL-10 profoundly inhibits the IFN-γ-induced expression of MHC class II antigens on microglial cells and suppresses the expression and secretion of proinflammatory cytokines [11]. Additionally, IL-10 reduces inflammation-mediated neuronal degeneration through the inhibition of microglial activation [12]. In AD, IL-10 suppresses Aβ-induced inflammatory factors, such as IL-1α, IL-1β, IL-6, and TNFα, to protect neuronal tissue from Aβ-induced cognitive impairment [13, 14]. In contrast with these findings, studies in AD model mice suggest that IL-10 inhibited the phagocytosis of microglia for Aβ through the JAK-STAT3 signal pathway to accelerate the deposition of Aβ plaques in the brain of mice, while inhibition of IL-10 restored the phagocytosis of pathological microglia for Aβ and improved the cognitive impairment of mice [15–17]. These conflicting results highlight the complication of IL-10 effects in AD. We surmise that IL-10 may play different roles in different stages of AD progression. More studies should be carried out to determine the link between IL-10 expression and AD prognosis. The effects of IL-10 are mediated by the corresponding receptor complexes IL-10RA and IL-10RB, in which the main role of IL-10RB is to activate the downstream JAK/STAT signaling pathway [18, 19]. The human IL-10RB gene is constitutively expressed in most human tissues [10]. Deletions or mutations of the IL-10RB gene is relevant to various diseases, such as inflammatory bowel disease, hepatitis B, coronavirus disease 2019 (COVID-19), and ischemic stroke [20–24]. In the central nervous system, the IL-10RB gene is co-expressed with genes involved in synaptic formation [25]. Additionally, Runge et al. have demonstrated that CD4+ T cells are capable of crossing the blood-brain barrier and inducing the expression of the central IL-10RB gene to mediate neuroprotection. CD4+ T cells lacking IL-10RB showed reduced differentiation into regulatory CD4+ T cells, which enhanced T cell co-stimulation, excessive microglia activation, and synaptic elimination [26]. However, the underlying transcriptional regulatory mechanism of the IL-10RB gene remains elusive.
Gene transcription is tightly controlled by the interaction between promoter elements and transcription factors. Transcription factors of the Sp family are highly conserved DNA-binding proteins that have zinc finger DNA domains to recognize GC box sequences (GGGGCGGGG) [27]. Sp proteins play a vital role in AD and other neurodegenerative diseases [28, 29]. Moreover, Sp proteins influence the pathological process of AD by regulating the transcription of APP, BACE1, BACE2, and Tau genes related to AD [30–33]. In addition, Sp proteins regulate the expression of other genes to influence neuronal survival and synaptic plasticity in AD [34–37]. Sp8 and Sp9 are two closely related button-like transcription factors with high homology that are widely expressed in the embryonic ganglionic eminences [38]. Sp8 and Sp9 coordinately regulate the differentiation, migration, and survival of olfactory bulb (OB) interneurons, dopamine receptor Drd2-expressing medium spiny neurons (D2-MSNs), and caudal ganglionic eminence (CGE)-derived cortical interneurons [39–41].
In this study, we analyzed the expression level of IL-10RB in AD-related database and AD model mice, characterized the IL-10RB promoter, and elucidated the molecular mechanism of IL-10RB gene transcription regulation. We first found that the IL-10RB is elevated in the brains of AD patient and AD model mice. The minimal promoter of the IL-10RB gene is located in the –90 to +51 bp region (relative to the TSS) and is sufficient for high-level expression of the IL-10RB gene. In addition, we demonstrated that the transcription factors Sp8 and Sp9 play a crucial role in the regulation of IL-10RB gene expression.
MATERIALS AND METHODS
Plasmids, siRNAs, and site-directed mutagenesis
A 3176 bp fragment and a series of deletion fragments of the 5′-flanking region of the IL-10RB gene from –3075 to +101 bp were amplified by PCR using genomic DNA extracted from human embryonic kidney 293T (HEK293T) cells as a template. Primers were designed using the SnapGene software with restriction enzyme sites compatible with the multi-cloning site of pGL4.10 vector (Promega, USA). The PCR reaction was performed as follows: 94°C for 1 min, 34 repeated cycles at 98°C for 10 s, 63°C for 30 s, and 72°C for X min (X is defined by the fragment length in kbp), followed by 7 min at 72°C. A total of eight deletion fragments were cloned into the promoterless luciferase reporter plasmid pGL4.10 at the KpnI and HindIII restriction sites by homologous recombination. Site-directed mutations based on pIL-10RB-90+51 were generated using overlap extension PCR (SOE PCR) [42]. The mutations were then confirmed by sequencing. The CDS sequences of human Sp1, Sp3, Sp4, Sp8, and Sp9 genes were amplified and inserted into the pcDNA4 vector (Promega) at the BamHI and XhoI sites. Sp8 siRNA and Sp9 siRNA were designed and synthesized by the Ribobio Company (Guangzhou, China). The siRNA sequences are as follows: si-Sp8: GCCGCTACCTGTAATAAGA; si-Sp9: CAGTCGGCCTTCATTTCCA. All primers are listed in Table 1.
Primer sequences for IL-10RB and SP8/9 gene cloning
The mutation sites were underlined in the bold italic face.
Cell culture, transfection, and luciferase assay
HEK293T and U251 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco, USA) containing 10% fetal bovine serum (FBS, Gibco) in a humidified incubator at 37°C with 5% CO2. Cells were vaccinated and grown to approximately 70–80% confluence for transfection using Lipofectamine 2000 (Invitrogen, USA) according to the supplier’s protocol. Then, 24μg of plasmids were transfected into a 10-cm dish for nuclear protein extraction, 3μg of plasmids or siRNAs (50 nM) were transfected per well of 6-well plates for RNA and proteins extraction, and 270 ng of reporter plasmids were transfected per well of a 48-well plate for a luciferase reporter assay with 30 ng of pRluc as an internal control. One hundred and thirty-five ng of expression plasmids or siRNAs (50 nM) were transfected per well of a 48-well plate for a luciferase reporter assay, together with 135 ng of luciferase reporter plasmids and 30 ng of control pRluc plasmids. The cells were harvested after 36 h, and firefly and Renilla luciferase activities were measured using a dual-luciferase reporter assay system (Promega, E1910). The relative luciferase activities were calculated by normalizing the fluorescence luciferase with Renilla luciferase as an internal standard.
Nuclear extraction and EMSA
Sp8- or Sp9-enriched nuclear extracts (NEs) were obtained from HEK293T cells transfected with 24μg expression plasmids pcDNA-Sp8 or pcDNA-Sp9 containing Myc tags for 36 h according to the NE-PERtrademark Nuclear and Cytoplasmic Extraction Protocol (Thermo Scientific, 78833, USA). NEs were then quantified with the BCA protein assay kit (Thermo, 23227) and aliquoted into 1μg/μl to store at –80°C. Sp consensus probes (Sp consensus*), Sp-A probes (IL-10RB Sp-A*), and Sp-B probes (IL-10RB Sp-B*) were synthesized and labeled with Alexa Fluor 700 in the 5′-flanking end (Thermo Scientific). In addition, other unlabeled oligonucleotides were also synthesized and hybridized into double-strands (see Table 2 for sequences). For binding reactions, 0.5μl of labeled oligomers (5 fmol) were incubated with 2μl of NEs (2μg) in binding buffer (1-μl 10X binding buffer, 25-nmol DTT, 50-nmol MgCl2, and 12.5-ng poly (dI-dC)) in a total volume of 10μl at room temperature (RT). For competition analysis, 200-fold (1 pmol) excess of unlabeled double oligonucleotides were incubated with 2μl of NEs and 5 fmol of labeled probes for 20 min at RT. For supershift analysis, 3μl of antibodies against Myc (Cell Signaling Technology, 2276, USA) were incubated with 2μl of NEs for 20 min at RT, followed by an additional 20 min incubation with 5 fmol of labeled probes. The protein–DNA complexes were analyzed on a 4% nondenaturing polyacrylamide gel at 80 V for 60 min in 1× TGE buffer in darkness. The gel was scanned using the LI-COR Odyssey (LI-COR Biosciences, USA) at a wavelength of 700 nm.
Sequences of oligonucleotides for EMSA, ChIP, and qRT-PCR
*indicates the probe is labeled with an Alexa Fluor 700 at the 5′ flanking end. The italic boldface represents the binding sites of Sp8 and Sp9.
ChIP
ChIP was performed using the SimpleChIP Enzymatic Chromatin IP kit (Cell Signaling Technology, 9003) following the supplier’s protocol. First, HEK293T cells in 10-cm dishes were transfected with the expression plasmids Myc-tagged Sp8 and Sp9. After 36 h, the cells were treated with formaldehyde to cross-link the chromatin-associated proteins to DNA. The samples were then digested with micrococcal nuclease and ultrasound to shear the DNA into 150–900 bp fragments (verified by performing agarose gel electrophoresis). Equal aliquots of chromatin supernatants were immunoprecipitated with a mouse monoclonal antibody against Myc (Cell Signaling Technology, 2276) or normal mouse IgG (Cell Signaling Technology, 61656) at 4°C overnight. DNA was extracted and used as a template to amplify the 212 bp targeted fragment in the IL-10RB promoter. Meanwhile, the targeted sequences were also amplified using a DNA template without Sp8 and Sp9. In addition, a 73 bp fragment of the IL-10RB exon 3 lacking Sp8 and Sp9 binding sites was also amplified using the DNA template with Sp8 and Sp9. All ChIP assays were performed at least three times, and representative results are presented. The primers used for PCR are shown in Table 2.
RNA extraction and quantitative real-time PCR
Total RNA was isolated from HEK293T and U251 cells using RNA extraction kits (BioTeke, RP1201, USA), and first-strand cDNAs were synthesized from 1μg of total RNA using the PrimeScript RT Kit (Takara, RR047A, Japan). The mRNA levels of Sp8 and Sp9 were determined with SYBR green real-time RT-PCR (Takara, RR820A) on the CFX96 Real-Time PCR Detection System (Bio-Rad CFX96, USA). GAPDH was used as an internal control to normalize the variability in expression levels. The primer sequences are shown in Table 2. The ΔΔCt method was used to transform Ct values into relative quantities.
Western blotting
Nine-month-old APP23 mice carrying human Swedish mutant APP751 were sacrificed by rapid decapitation, and the hippocampus tissue was homogenized and lysed in RIPA lysis buffer containing protease inhibitor (Roche, Switzerland) for protein assays [43]. For cultured cells, whole-cell samples were lysed in RIPA lysis buffer with a protease inhibitor (Roche, Switzerland) for 20 min on ice and were then centrifuged at 12,000× g for 15 min at 4°C. Protein concentrations were determined using the BCA protein assay kit (Thermo, 23227). Equal amounts of proteins were diluted in 5× SDS sample buffer, resolved on 10% Tris-glycine SDS PAGE gels, and transferred onto polyvinylidene difluoride (PVDF) membranes (Immobilon-P, Millipore, USA). Nonspecific binding was blocked with 5% skim milk in Tris-Buffered Saline-Tween (TBST) for 60–90 min at RT. The samples were incubated with a primary anti-IL-10RB (Proteintech, 15102-1-AP), anti-Sp8 (Abcam, ab73494, British), anti-Sp9 (Invitrogen, PA5-70643), or GAPDH (Proteintech, 60004-1-lg) antibody at 4°C overnight, followed by 60–90 min of incubation with a secondary HRP-conjugated goat anti-rabbit or goat anti-mouse (1:3000 dilution) (PerkinElmer, USA) antibody. Finally, the immunoreactivity was determined by ECL detection according to the manufacturer’s instructions (Pierce, USA). The data were analyzed using the Image J software.
Statistical analysis
All experiments were repeated independently three or more times. All results are presented as means±SEM. Statistical comparisons were conducted using one-way analysis of variance (ANOVA) or Student’s t-test. In all cases, p≤0.05 was considered to be statistically significant.
RESULTS
The expression of IL-10RB gene is elevated in AD brains
To investigate whether there is a differential expression for IL-10RB in brain tissues from AD patients and normal controls, we analyzed the expression data from AlzData database and found that the human IL-10RB gene was expressed at a significantly higher level in the cerebral cortex and hippocampus from AD patients compared with healthy controls (Fig. 1A). Furthermore, the protein levels of IL-10RB in the hippocampus of wild-type and AD model mice APP23 were analyzed using western blot (Fig. 1B). The results showed that the protein levels of IL-10RB were higher in the hippocampus of APP23 mice than that in wild-type mice (Fig. 1C).
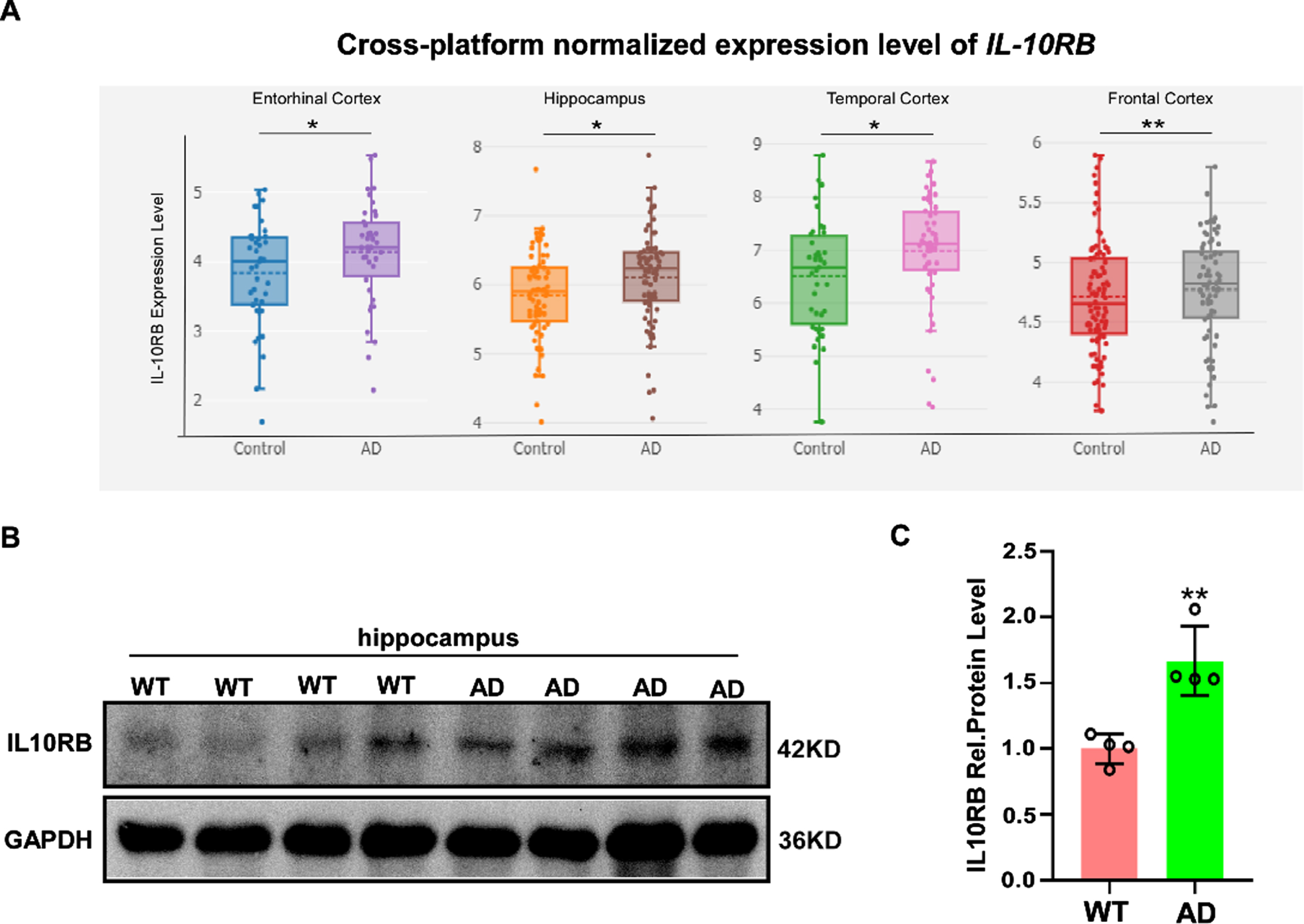
The expression of the IL-10RB gene is high in AD brain. A) The expression data of IL-10RB gene was obtained from AlzData database (https://www.AlzData.org). The data shows the difference of expression for the IL-10RB gene in healthy- and AD- derived brains. Each dot represents one sample. B) WT and AD mice (APP23 mice of the 9-month age) were sacrificed and hippocampus were isolated for the immunoblotting analysis of IL-10RB and GAPDH. C) Quantitative analysis of the IL-10RB protein level. Values indicate means±SEM (n = 4 per group) by student’s t-test (**p < 0.01).
Structural characterization of the human IL-10RB gene
The encoding human IL-10RB gene is located within a gene cluster on human chromosome 21 (Fig. 2A) that consists of the IL-10RB, IFN αR1, and IFNγR2 genes [44]. The human IL-10RB gene has seven exons that generate only one transcript variant (NM000628.5) and encode for a 37-kD protein of 326 amino acids (Fig. 2B). The human IL-10RB gene is a member of the class II cytokine receptor family (CRF2) and is shared with the expanded families of class II cytokines IL-10, IL-22, IL-26, IL-28, and IL-29 [45]. Signals from these cytokines are transduced via the JAK-STAT signaling pathway [10, 46] (Fig. 2C). Furthermore, analyzing the 801 bp proximal promoter fragment from –700 to +101 bp of the IL-10RB gene using the online databases UCSC and JASPAR, we found that the proximal promoter of the IL-10RB gene lacks typical TATA boxes and contains several putative regulatory elements, including SNAI1, Sp1, Sp3, Sp4, Sp8, Sp9, and MAZ (Fig. 2D).
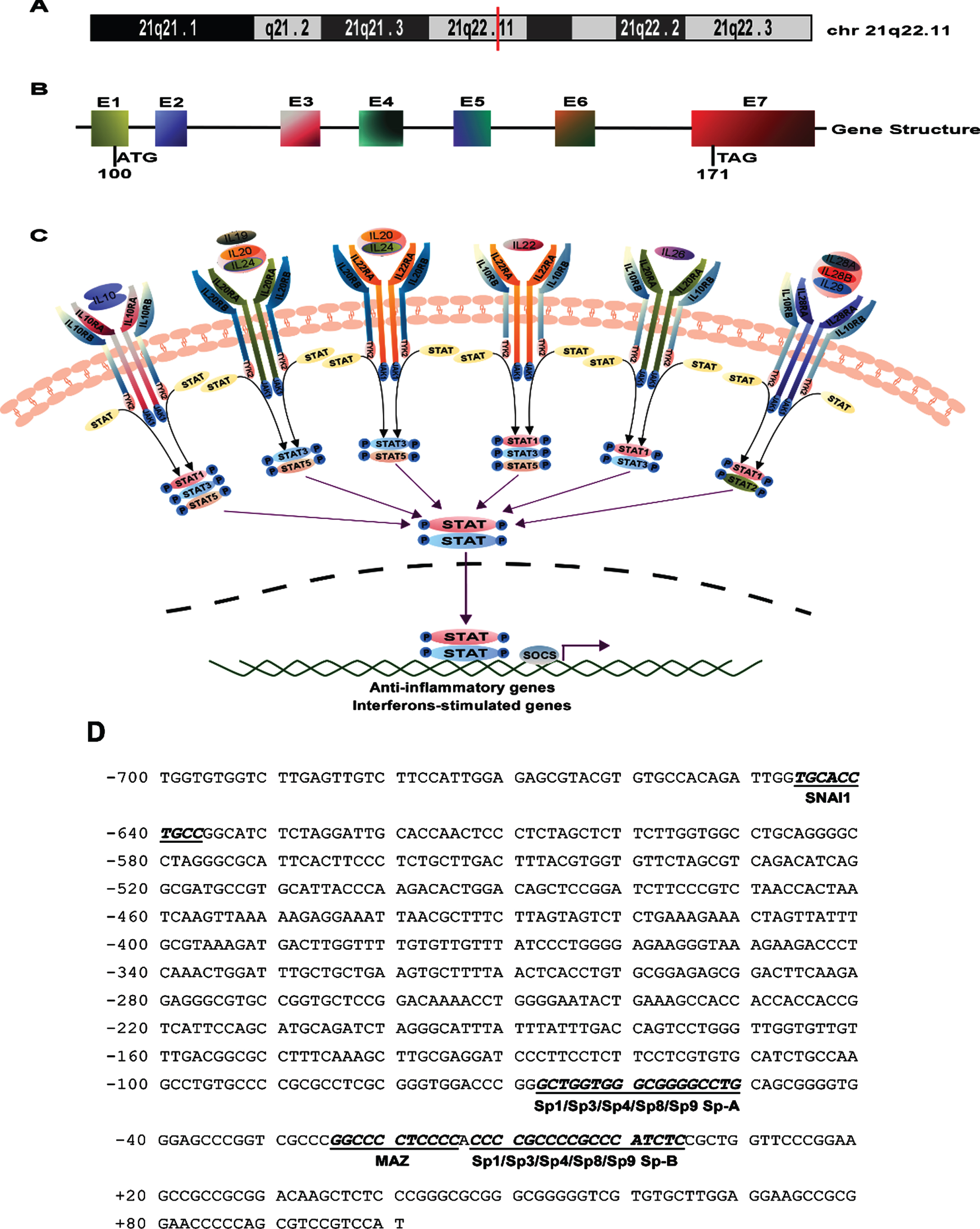
The structure and pathway related to the IL-10RB gene. A) The location of the human IL-10RB gene was marked by a red vertical line on chromosome 21. B) The gene structure diagram of the IL-10RB gene. ATG is the translation start codon and TAA is the stop codon. E represents exon. C) The expanded family of the class II cytokines share the IL-10RB chain and JAK-STAT signaling pathway. D) The promoter sequence of the IL-10RB gene from –700 to +101 bp. The adenine +1 represents the first base of the TSS in the first exon. The putative transcription factor binding sites are displayed in italic boldface and the names of the transcription factors are annotated below the line.
Functional characterization of the human IL-10RB gene promoter
To identify the minimal promoter of the IL-10RB gene, a series of deletion fragments of the 5′-flanking region of the IL-10RB gene were cloned into a promoterless luciferase reporter plasmid pGL4.10 vector. Luciferase activity in cells transfected with the pGL4.10 plasmid depends on a functional promoter inserted upstream of the luciferase coding sequence in the pGL4.10 vector. Four plasmids, containing various lengths from –3075 to +101 bp (relative to the TSS) of the 5′-flanking region of the IL-10RB gene, were constructed and transfected into HEK293T and U251 cells along with the Renilla luciferase vector pRluc. These plasmids were named pIL-10RB-3075+101, pIL-10RB-2055+101, pIL-10RB-884+101, and pIL-10RB-246+101 (Fig. 3A). Compared with cells transfected with pGL4.10, HEK293T and U251 cells transfected with pIL-10RB-3075+101 showed a significantly higher luciferase activity (Fig. 3C, D). The data indicated that the –3075 to +101 bp region of the IL-10RB gene contains the functional promoter. The deletion plasmids pIL-10RB-2055+101 and pIL-10RB-884+101 had little impact on promoter activity compared with pIL-10RB-3075+101. However, a further deletion at the 5′ end from –884 to –246 bp (pIL-10RB-246+101) strongly increased the luciferase activity (Fig. 3C, D). This result suggested that the 638 bp fragment from –884 to –246 bp contains important negative regulatory elements required for IL-10RB gene transcription.

Deletion analysis of the human IL-10RB promoter in HEK293T and U251 cells. A) Schematic diagram of the IL-10RB promoter deletions. The numbers represent the endpoints of the IL-10RB inserted into the pGL4.10 vector relative to the TSS. The arrows show the direction of transcription. B) Luciferase reporter plasmids were confirmed by restriction enzyme Kpn I and Age I digestion and were analyzed on a 1.0% agarose gel. C, D) The deletion plasmids were cotransfected with the pRLuc into HEK293T and U251 cells, respectively. Luciferase activity was measured and showed in relative luciferase units (RLU). The values represent means±standard error of the mean (SEM) of no less than three independent experiments by ordinary one-way ANOVA multiple comparisons (**p < 0.01, ***p < 0.001, ****p < 0.0001).
To further narrow down this important region, more deletion plasmids were generated and transfected into HEK293T and U251 cells. Deletions from +101 to +51 bp at the 3′ end and from –246 to –90 bp at the 5′ end led to little change of promoter activity in HEK293T cells (Fig. 3C), whereas a deletion from +101 to +51 bp at the 3′ end resulted in a robust decrease of promoter activity in U251 cells (Fig. 3D). This result indicated that the region from +101 to +51 bp plays a crucial regulatory function for the IL-10RB gene in U251 cells, while it does not have such an effect in HEK293T cells. Further deleting a 69 bp fragment from +51 to –18 bp at the 3′ end of IL-10RB based on the pIL-10RB-90+51 plasmid drastically reduced luciferase activity to a low level, which suggested that the 69 bp fragment is essential for IL-10RB promoter activity. Deleting a 40 bp fragment from –90 to –50 bp at the 5′ end of IL-10RB based on the pIL-10RB-90+51 sharply reduced luciferase activity, indicating that the potential positive regulatory elements are contained in the –90 to –50 bp region in HEK293T and U251 cells (Fig. 3C, D). Our data demonstrated that the 141 bp fragment from –90 to +51 bp at the 5′-flanking end functions as the minimal promoter region and is sufficient for high-level expression of the IL-10RB gene.
Sp8 and Sp9 enhanced human IL-10RB promoter activity
As shown in Fig. 2D, the binding sites of transcription factors of the Sp family are located in the key regions from –90 to +50 bp of the IL-10RB promoter. Therefore, we focused on whether Sp proteins play a key role in regulating IL-10RB promoter activity. First, the expression plasmids of Sp proteins were constructed and named pcDNA-Sp1, pcDNA-Sp3, pcDNA-Sp4, pcDNA-Sp8, and pcDNA-Sp9. These plasmids were then transfected into HEK293T and U251 cells along with pIL-10RB-90+51. pRLuc was also transfected for transfection efficiency normalization. After 36 h, the luciferase activity was measured. The results indicated that Sp proteins increase the luciferase activity of pIL-10RB-90+51 in HEK293T and U251 cells (Fig. 4A, B). In view of the significant impact of Sp8 and Sp9 on IL-10RB promoter activity, we wanted to further explore the regulatory function of Sp8 and Sp9 in the transcription of the IL-10RB gene. Different concentrations of Sp8 and Sp9 expression plasmids were cotransfected with pIL-10RB-90+51 into HEK293T cells or U251 cells. The luciferase activity showed that IL-10RB promoter activity is positively correlated with the concentration of Sp8 (Fig. 4C, E) and Sp9 (Fig. 4D, F). In addition, knockdown of the endogenous expression of Sp8 and Sp9 by siRNA decreased IL-10RB promoter activity by 15% and 20% in HEK293T cells (Fig. 4G) and by 35% and 31% in U251 cells (Fig. 4H), respectively. These results indicated that IL-10RB promoter activity is significantly enhanced by Sp8 and Sp9.

Sp8 and Sp9 upregulate the human IL-10RB promoter activity. A, B) The overexpression of exogenous Sp8 and Sp9 increased the IL-10RB promoter activity. Sp plasmids were cotransfected with pIL-10RB-90+51 into HEK293T (A) or U251 (B) cells, respectively. After 36 h, the luciferase activity was measured and expressed in Relative Luciferase Units (RLU). C-F) Sp8 and Sp9 enhanced promoter activity of the IL-10RB gene in a dose-dependent manner. Sp8 and Sp9 at different concentrations were cotransfected with pIL-10RB-90+51 into HEK293T cells (C, D) or U251 cells (E, F), respectively. G, H) Knockdown of the endogenous Sp8 and Sp9 decreased the IL-10RB promoter activity. Sp8 siRNA and Sp9 siRNA (50 nM) were cotransfected with pIL-10RB-90+51 into HEK293T (G) or U251 (H) cells, respectively. After 36 h, the luciferase activity was measured. Each bar represents means±SEM of no less than three independent experiments by ordinary one-way ANOVA multiple comparisons (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
To further explore whether Sp8 and Sp9 regulate IL-10RB promoter activity by binding the Sp-A or/and Sp-B in the –90 to +50 bp region of the IL-10RB promoter, we carried out mutant analyses of Sp-A and Sp-B sites one by one or together (Figs. 2D, 5A). The mutant plasmids pIL-10RB-Mut a, pIL-10RB-Mut b, and pIL-10RB-Mut a+b based on pIL-10RB-90+51 were constructed and cotransfected with the expression plasmids pcDNA-Sp8 or pcDNA-Sp9 into HEK293T cells and U251 cells. The luciferase activity showed that IL-10RB promoter activity was increased when Sp8 and Sp9 were cotransfected into HEK293T or U251 cells compared with when the empty vector pcDNA4 was transfected (Fig. 5B–E lane 2). However, IL-10RB promoter activity was sharply reduced when the binding site Sp-A of Sp8/Sp9 was mutated (Fig. 5B–E lane 3). Interestingly, a mutation of the binding site Sp-B had little effect on the promoter activity of the IL-10RB gene in HEK293T cells (Fig. 5B, C lane 4), whereas the mutation had a significant impact on IL-10RB promoter activity in U251 cells (Fig. 5D, E lane 4). Mutations of both Sp-A and Sp-B sites led to a stronger inhibitory effect on IL-10RB promoter activity than a sole mutation of the Sp-A or Sp-B site (Fig. 5B–E lane 5). The results indicated that Sp8 and Sp9 mainly regulate the promoter activity of the IL-10RB gene by the Sp-A binding site rather than the Sp-B binding site in HEK293T cells. However, Sp8 and Sp9 regulated IL-10RB promoter activity by both the Sp-A and Sp-B sites in U251 cells.

The analysis of the binding sites of Sp8 and Sp9 in the IL-10RB promoter. A) The binding sites Sp-A and Sp-B were mutated one by one or simultaneously. The mutant base sequences were annotated with italic bold letters. B-E) the mutant plasmids pIL-10RB-Mut a, pIL-10RB-Mut b, and pIL-10RB-Mut a+b were cotransfected with pcDNA-Sp8 or pcDNA-Sp9 into HEK293T cells (B, C) or U251 cells (D, E), respectively. After 36 h, luciferase assays were performed. Each bar represents means±SEM of no less than three independent experiments by ordinary one-way ANOVA multiple comparisons (**p < 0.01, ***p < 0.001, ****p < 0.0001).
The IL-10RB promoter contains cis-acting response elements of Sp8 and Sp9
To investigate whether Sp8 and Sp9 directly interact with the IL-10RB promoter via the putative Sp binding sites, an electrophoretic mobility shift assay (EMSA) was performed. First, Sp8 or Sp9 plasmids containing Myc tags were transfected into HEK293T cells to extract Sp8- or Sp9-enriched nuclear proteins, respectively. The Sp probes were then synthesized and labeled with Alexa Fluor 700 to acquire three labeled probes (Sp consensus*, IL-10RB Sp-A*, and IL-10RB Sp-B*) and their mutant and competitive unlabeled probes. Sp consensus* served as a positive control. As shown in Fig. 6A, D, the shifted protein-DNA complex bands were detected after incubating the Sp consensus* probes with Sp8- or Sp9-enriched nuclear extract (NE) (Fig. 6A, D lane 2). The shifted bands were significantly inhibited when adding a 200-fold excess of unlabeled Sp consensus competition oligonucleotides (Sp consensus) (Fig. 6A, D lane 3). Applying 200-fold excess of mutant Sp consensus oligonucleotides (Sp consensus Mut) led to the loss of the competing effect on Sp protein-DNA complex bands (Fig. 6A, D lane 4). More importantly, 200-fold excess of the putative cis-acting elements of Sp8 and Sp9 (IL-10RB Sp-A and IL-10RB Sp-B) could markedly reduce the intensity of the shifted bands (Fig. 6A, D lanes 5 and 7), while the mutations failed to compete for the complex bands (Fig. 6A, D lanes 6 and 8). Adding an anti-Myc antibody led to the formation of slower-migrating supershifted complexes (Fig. 6A, D lane 9), which confirmed the specific binding between Sp8 or Sp9 and Sp consensus*.
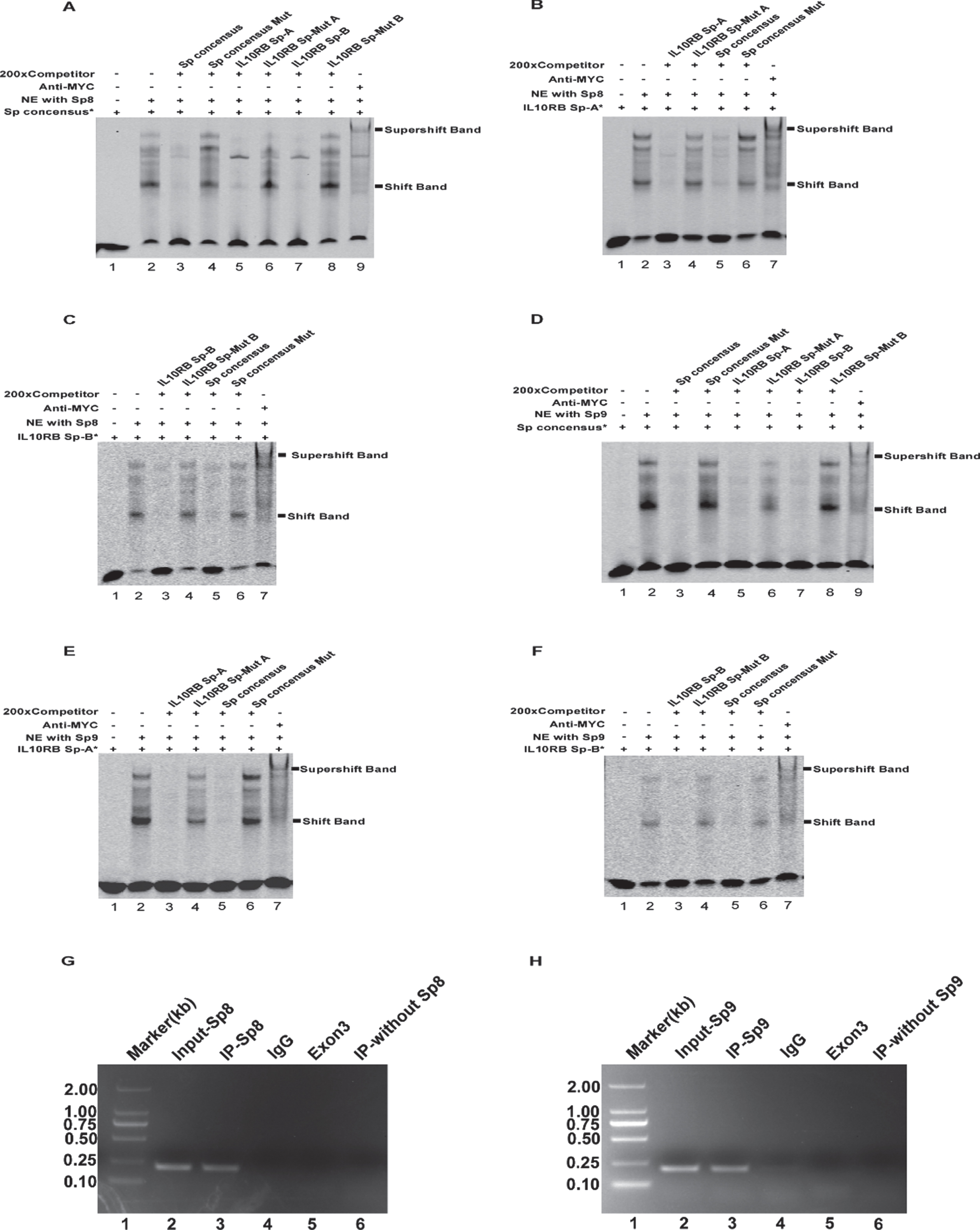
EMSA and ChIP assays for the IL-10RB promoter. A, D) EMSA for Sp consensus probes. Lane 1 was added Sp consensus* probes alone without NE used as a negative control. When labeled Sp consensus* probes were incubated with NE enriched in Sp8 or Sp9, lane 2 formed shifted DNA-protein complex bands. Competition assays were performed by adding 200-fold excess of unlabeled Sp consensus oligos (lane 3), Sp consensus Mut (lane 4), IL-10RB Sp-A/B (lanes 5, 7), and IL-10RB Sp-A/B Mut (lanes 6, 8). Lane 9 showed a supershifted band with the anti-Myc antibody. B, C, E, F) EMSA for putative Sp-A and Sp-B probes. Similarly, incubation NE with IL-10RB Sp-A or IL-10RB Sp-B generated shifted complex bands (lane 2). Competition assays were performed from lane 3 to lane 6 by adding 200-fold excess of unlabeled IL-10RB Sp-A/B, IL-10RB Sp-A/B Mut, Sp consensus, or Sp consensus Mut. Supershifted bands formed in lane 7 by adding an anti-Myc antibody. G, H) ChIP analysis for the IL-10RB promoter in HEK293T cells. The precipitated ChIP DNA was amplified by PCR and analyzed on a 1.5% agarose gel using primer pairs containing the putative Sp8 and Sp9 binding sites (the target locus) or exon 3 (a non-target locus) in the IL-10RB gene. The size of the DNA marker is 2 kb (lane 1). The input chromatin DNA samples without immunoprecipitation was amplified by PCR (lane 2). The targeted IL-10RB promoter fragment (212 bp) was amplified by PCR using DNA template immunoprecipitated by anti-Myc antibody (lane 3). ChIP DNA immunoprecipitated by mouse IgG2a was amplified by PCR used as a negative control (lane 4). A 73bp band in the exon3 of the IL-10RB gene was amplified using the DNA template immunoprecipitated by anti-Myc antibody (lane 5). The targeted IL-10RB promoter fragment was amplified using IP samples without Sp8 and Sp9 proteins as the template (lane 6).
To further determine the binding specificity between Sp8 or Sp9 and IL-10RB Sp-A or IL-10RB Sp-B in the IL-10RB promoter, we performed EMSA with labeled IL-10RB Sp-A* and IL-10RB Sp-B*. Consistently, the interaction between Sp8 or Sp9 and IL-10RB Sp-A* or IL-10RB Sp-B* formed shifted complex bands (Fig. 6B, C, E, F, lane 2). Meanwhile, 200× competition probes IL-10RB Sp-A/B and Sp consensus weakened the shifted bands (Fig. 6B, C, E, F, lanes 3 and 5), whereas the mutant probes IL-10RB Sp-Mut A/B and Sp consensus Mut had no effect (Fig. 6B, C, E, F, lanes 4 and 6). Supershifted bands were also detected (Fig. 6B, C, E, F, lane 7). Taken together, these data demonstrated that the IL-10RB promoter contains two functional Sp binding sites Sp-A and Sp-B, which could directly interact with Sp8 and Sp9 in vitro.

Sp8 and Sp9 regulate the IL-10RB gene transcription. A-D) The mRNA levels of IL-10RB gene were increased after Sp8 and Sp9 overexpression in HEK293T cells (A, B), whereas were decreased in U251 cells (C, D). E-H) Knockdown of Sp8 or Sp9 by siRNA (50 nM) significantly reduced the IL-10RB mRNA levels in HEK293T cells (E, F), while increased it in U251 cells (G, H). Sp8 and Sp9 plasmids or siRNA were transfected into HEK293T and U251 cells for 36 h, respectively. Total RNA was extracted for qRT-PCR to detect the mRNA levels of Sp8, Sp9, IL-10RB, and GAPDH using primers specific to the human Sp8, Sp9, IL-10RB, or GAPDH genes. GAPDH was used as the internal control. Each bar represents means±SEM of no less than three independent experiments by student’s t test (**p < 0.01, ***p < 0.001, ****p < 0.0001).
To further investigate whether Sp8 and Sp9 are bound to the Sp-A and Sp-B sites of the endogenous IL-10RB promoter in vivo, we utilized chromatin immunoprecipitation (ChIP) analysis. First, pcDNA-Sp8 and pcDNA-Sp9 plasmids containing Myc tags were transfected into HEK293T cells to acquire Sp8- and Sp9-enriched samples. The samples were then immunoprecipitated using an anti-Myc antibody that specifically discerns Sp8-Myc or Sp9-Myc fusion proteins and normal mouse IgG as a negative control. The DNA fragments were extracted from the precipitated samples and used as a template to amplify the targeted DNA sequence spanning the Sp-A (–68 to –50) and Sp-B sites (–13 to +5) of the IL-10RB promoter. The results suggested successful amplification of the targeted IL-10RB promoter fragments using a DNA template from Sp8 or Sp9-enriched samples (Fig. 6G, H lane 3), whereas the amplification failed when using DNA templates from samples immunoprecipitated by the IgG antibody (Fig. 6G, H lane 4) or samples without Sp8 or Sp9 (Fig. 6G, H lane 6). Meanwhile, the IL-10RB exon 3 without Sp8 and Sp9 binding sites was also not amplified using the DNA template from Sp8 or Sp9-enriched samples (Fig. 6G, H lane 5). The ChIP analysis demonstrated that the transcription factors Sp8 and Sp9 bind to the endogenous IL-10RB promoter in vivo.

Sp8 and Sp9 regulate the IL-10RB gene expression. A-H) The overexpression of exogenous Sp8 and Sp9 increased the IL-10RB protein levels in HEK293T cells (A-D), whereas the situation was totally contrary in U251 cells (E-H). I-P) The protein levels of IL-10RB gene were decreased by knockdown of Sp8 and Sp9 in HEK293T cells (I-L), whereas were increased in U251 cells (M-P). Western blot was performed after Sp8 and Sp9 plasmid or siRNA (50 nM) transiently transfected into HEK293T or U251 cells. GAPDH was used as an internal protein control. Each bar represents means±SEM of no less than three independent experiments by student’s t test (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Regulation of IL-10RB expression by Sp8 and Sp9 activity
To further explore whether Sp8 and Sp9 affect the endogenous transcription of the IL-10RB gene, qRT-PCR was performed. The expression plasmids and siRNAs of Sp8 and Sp9 were transfected into HEK293T and U251 cells. Total RNA was then extracted 36 h later to perform qRT-PCR. GAPDH was used as an internal control. qRT-PCR showed that Sp8 and Sp9 overexpression leads to increased IL-10RB mRNA levels in HEK293T cells (Fig. 7A, B) and decreased IL-10RB mRNA levels in U251 cells (Fig. 7C, D). Furthermore, siRNAs against Sp8 and Sp9 caused a 45% and 48% reduction of IL-10RB mRNA, respectively, compared to a negative control in HEK293T cells (Fig. 7E, F). However, these siRNAs increased IL-10RB mRNA by 3.2-fold and 4.0-fold, respectively, in U251 cells (Fig. 7G, H). These results demonstrated that the transcription factors Sp8 and Sp9 positively regulate IL-10RB transcription in HEK293T cells but negatively regulate IL-10RB transcription in U251 cells.
To investigate whether overexpression or knockdown of Sp8 and Sp9 affected IL-10RB gene expression, the protein levels of the IL-10RB gene were determined by western blot. GAPDH served as an internal control. The results showed that Sp8 and Sp9 overexpression significantly increases IL-10RB protein levels in HEK293T cells (Fig. 8A–D) but decreases them in U251 cells (Fig. 8E–H). In addition, Sp8 and Sp9 knockdown caused a marked reduction of IL-10RB protein levels in HEK293T cells (Fig. 8I–L) but an increase of IL-10RB protein levels in U251 cells (Fig. 8M–P). In sum, these results suggested that Sp8 and Sp9 positively regulate IL-10RB expression in HEK293T cells, while they negatively regulate it in U251 cells.
DISCUSSION
IL-10RB mRNA and protein have been detected in microglia, astrocytes, and oligodendroglia. IL-10 suppresses the secretion of pro-inflammatory cytokines via IL-10RB signaling transduction [11]. IL-10RB activation in neurons enhances Bcl-2 and Bcl-xL transcription for neuronal survival [47]. Restoring the normal level of IL-10 and IL-10 receptors in AD could be an effective treatment strategy for AD. Here, we detected for the first time the differential expression of IL-10RB in normal mice and AD model mice, characterized the promoter of the IL-10RB gene, and demonstrated that Sp8 and Sp9 regulate the promoter activity and expression level of the IL-10RB gene in HEK293T and U251 cells.
Combining AlzData database and western blot analysis, we found that there was a higher expression of the IL-10RB gene in AD patients or mice than that in normal controls (Fig. 1). Given the intricate role of IL-10 in AD, it is difficult to speculate whether increased IL-10RB plays a protective or destructive role in AD. Therefore, it is necessary to further study the effect of IL-10RB in AD. To determine the promoter character of the IL-10RB gene, we cloned a 3176 bp fragment of the 5′-flanking region of the IL-10RB gene. We found that the promoter sequence does not have typical TATA boxes but contains high GC content and several putative regulatory elements, including SNAI1, Sp1, Sp3, Sp4, Sp8, Sp9, and MAZ (Fig. 2D). We determined that the –90 to +51 bp sequence is the minimal promoter region of the IL-10RB gene using a luciferase assay. In addition, the –246 to +101 bp region had a stronger promoter activity than the –246 to +51 bp and –90 to +51 bp sequences in U251 cells, whereas the opposite was true in HEK293T cells (Fig. 3C, D). This result suggests that the region from +101 to +51 bp contains important regulatory elements in U251 cells, while it does not in HEK293T cells. Sp family members are frequently involved in the basal expression of many TATA-less promoters [48], and two predicted binding elements for Sp8 and SP9 are located in the IL-10RB proximal promoter region (no TATA boxes). Sp8 and Sp9 have similar structures and bind to the cognate Sp protein binding sites, although their DNA-binding properties and regulatory functions are different [38]. Sp8 and Sp9 play an essential role in the area patterning of the cerebral cortex; molecular specification of subsets of cortical layer neurons; and differentiation of cortical progenitors through regulating the transcription of corresponding genes in neuronal cells [49–51]. We postulated that Sp8 and Sp9 regulate IL-10RB gene transcription. Using a luciferase assay, we demonstrated that IL-10RB promoter activity was largely upregulated with the overexpression of exogenous Sp8 and Sp9 (Fig. 4A–F) and decreased with the knockdown of Sp8 and Sp9 (Fig. 4G, H). Site-directed mutations of the Sp8 and Sp9 binding site Sp-A significantly reduced IL-10RB promoter activity, while mutations of the binding site Sp-B had little effect in HEK293T cells (Fig. 5B, C). Interestingly, mutations of both the binding sites Sp-A and Sp-B had significant effects on the promoter activity of the IL-10RB gene in U251 cells (Fig. 5D, E). The results indicate that Sp8/Sp9 regulates the promoter activity of the IL-10RB gene via different mechanisms in HEK293T and U251 cells. Meanwhile, the results also prove that the binding site Sp-A is more important than Sp-B for Sp8 and Sp9 in mediating IL-10RB promoter activity. Furthermore, IL-10RB promoter activity was not entirely inhibited by the mutation of Sp-A or Sp-B (Fig. 5B–E), indicating that there may be other undiscovered transcription factor(s) crucial for the transcriptional regulation of the IL-10RB gene. Using EMSA and ChIP assays, we confirmed that Sp8 and Sp9 bind to the IL-10RB promoter in vivo and in vitro (Fig. 6).
More importantly, we demonstrated that the overexpression or knockdown of Sp8 and Sp9 affects the expression of the endogenous IL-10RB gene using qRT-PCR and western blot (Figs. 78,). It should also be noted that Sp8 and Sp9 positively regulate the expression of the IL-10RB gene in HEK293T cells (Figs. 7A, B, E, F and 8A–D, I–L), while they negatively regulate its expression in U251 cells (Figs. 7C, D, G, H and 8E–H, M–P). The results suggest that the same transcription factor may have different regulatory functions in different cell types. Moreover, transcriptional regulation of genes is often extremely complex and is achieved by subtle interplay between RNA polymerase, various transcription factors and promoter sequences. Luciferase assays only showed the regulation function of Sp8/9 on the minimal promoter of IL-10RB gene, which could not completely simulate the regulation effect of Sp8/9 on endogenous IL-10RB gene. The qPCR and western blot assays showed the regulation effect of Sp8/9 on endogenous IL-10RB gene. In HEK293T cells, luciferase assay, qPCR and western blot showed consistent effects of Sp8/9 to increase transcription and expression of IL-10RB gene. But in U251 cells, luciferase assay, qPCR and western blot showed inconsistent effects. A possible explanation is that in U251 cells, there may be other unknown transcription factors. These transcription factors are regulated by Sp8/9 or interact with them, and work together at the promoter-distal sites outside the minimal IL-10RB promoter to play a transcriptional regulatory role on endogenous IL-10RB gene.
In conclusion, our study found that the IL-10RB gene is elevated in AD patient brains and AD model mice. It will be interesting to further investigate the role of IL-10RB gene in AD. We characterized the IL-10RB promoter and identified the IL-10RB minimal promoter. In addition, our experiments revealed that both Sp8 and Sp9 regulate the promoter activity of the IL-10RB gene and affect its expression, possibly through cooperative regulation. It would be interesting to further explore whether Sp8 and Sp9 interact in regulating the expression of the IL-10RB gene. Furthermore, we found it essential to explore the regulatory function of other Sp family’s transcription factors predicted in the IL-10RB gene (Fig. 2D). To our knowledge, this is the first report on the transcriptional regulation of the IL-10RB gene. Our results could contribute to refining and extending the understanding of the modulatory mechanism of IL-10RB gene expression.