
Abstract
Background:
Imbalance between the production and clearance of amyloid-β (Aβ) promotes the development of Alzheimer’s disease (AD). Presenilin-1 (PS1) is the catalytic subunit of γ-secretase, which is involved in the process of Aβ production. The profiles of autoantibodies are dysregulated in AD patients.
Objective:
This study aims to investigate the relative levels and clinical relevance of naturally occurring antibodies to PS1 (NAbs-PS1) in AD.
Methods:
A total of 55 subjects with AD (including both dementia and mild cognitive impairment due to AD), 28 subjects with cognitive impairment (including both dementia and mild cognitive impairment) not due to AD (non-AD CI), and 70 cognitively normal (CN) subjects were recruited. One-site ELISA was utilized to determine the relative levels of NAbs-PS1 in plasma.
Results:
AD subjects had lower plasma levels of NAbs-PS1 than CN and non-AD CI subjects. Plasma NAbs-PS1 were negatively associated with the brain Aβ load, as reflected by PET-PiB SUVR, and were positively associated with cognitive functions of participants. Plasma NAbs-PS1 discriminated AD patients from CN with an area under the curve (AUC) of 0.730, a sensitivity of 69.09%, and a specificity of 67.14%, and they discriminated AD patients from non-AD CI subjects with an AUC of 0.750, a specificity of 70.91%, and a sensitivity of 71.43%.
Conclusion:
This study found an aberrant immunological phenotype in AD patients. Further investigations are needed to determine the pathophysiological functions of NAbs-PS1 in AD.
INTRODUCTION
Alzheimer’s disease (AD) is characterized by the extracellular deposition of senile plaques formed by aggregated amyloid-β (Aβ) peptides and the intracellular neurofilament tangles formed by hyperphosphorylated tau proteins [1]. AD is currently incurable, partly due to the largely unrevealed pathogenic networks. However, the imbalance between the production and clearance of Aβ is a well-recognized mechanism of AD.
Aβ peptide is generated through sequential cleavage of amyloid-β protein precursor (AβPP) by β- and γ-secretases [2]. It is suggested that the activities of these secretases are increased during aging and the development of AD [2]. Therefore, these secretases have become potential therapeutic targets of AD. Presenilin-1 (PS1) is the major catalytic subunit and a key regulator of the activity of the γ-secretase complex [3]. The genetic mutation in PS1 genes causes early-onset familial AD through promoting the production of Aβ. Besides, changes in the conformation of endogenous wild-type PS1, similar to those found with mutant PS1, occur in sporadic AD brain and during normal aging [4].
Dysregulation of the innate and adaptative immunity appears to be a significant feature of AD, and a mounting body of evidence suggests that pathological alterations in humoral immune responses promote the development of AD [5, 6]. Previous studies have identified a panel of dysregulated autoantibodies in AD [7], including those targeting Aβ [8], microtube-associated proteins [9], and neuronal surface antigens [10]. Some of these autoantibodies are also brain-reactive and may have important pathophysiological functions [11]. In the present study, we investigated the relative levels and clinical relevance of naturally occurring antibodies to PS1 (NAbs-PS1) in AD patients.
METHODS
Participants
A total of 55 AD subjects, including both dementia and mild cognitive impairment (MCI) due to AD, 28 subjects with cognitive impairment not due to AD (non-AD CI), including both dementia and MCI not due to AD, and 70 cognitively normal (CN) subjects were consecutively recruited from Department of Neurology, Daping Hospital, Third Military Medical University. Subjects were excluded if they fulfiled one or more of the following conditions: 1) severe cardiac, pulmonary, hepatic, renal, or neoplastic disorders; 2) autoimmune diseases which could potentially have dysregulated autoantibody profiles; and 3) refused to participate in this study. Written consents for participation and blood and cerebrospinal fluid sampling were obtained from all participants or their legal representatives.
As to the clinical assessment procedures, the general cognitive status was first examined with the Mini-Mental State Examination (MMSE). Subjects with an MMSE score lower than 27 were further subjected to a battery of neuropsychological tests, including the Clinical Dementia Rating (CDR), Montreal Cognitive Assessment (MoCA), Activities of Daily Living (ADL), Clock Drawing Task (CDT), Trail Making Test (TMT), Auditory Verbal Learning Test (AVLT), Digital Span Test (DST), Boston Naming Test (BNT), Pfeffer’s Outpatient Disability Questionnaire (POD), Hachinski Ischemic Score (HIS), Neuropsychiatric Inventory (NPI), the Geriatric Depression Scale (GDS), and Self-rating Anxiety Scale (SAS) [12]. CDR was used to judge cognitively normal (CDR = 0), MCI (CDR = 0.5), and dementia (CDR> =1). Participants with CDR of 0.5 or higher further received an MRI scan, a PET scan using 11 C Pittsburgh Compound B (PiB) tracer (Aβ PET), and a Fluorodeoxyglucose (FDG)-PET scan to confirm the etiology of cognitive impairment. All participants received APOE genotyping and blood tests including blood cell count, fast glucose, cholesterol, hepatic and renal functions, vitamin B12, folic acid, and thyroxin. The diagnosis of AD was made according to the criteria of the National Institute on Aging and Alzheimer’s Association (NIA-AA) [13, 14]. The differential diagnosis between AD and non-AD CI was determined by the Aβ PET scan. The term “non-AD CI” was used to describe subjects with cognitive impairment but with no or sparse Aβ deposition with a standardized uptake value ratio (SUVR) below 1.42 [15]. The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Institutional Review Board of Daping Hospital.
Determination of plasma NAbs-PS1
Relative levels of NAbs-PS1 in human plasma were determined by one-site enzyme-linked immunosorbent assay (ELISA). Recombinant PS1 protein (Abcam, USA) was dissolved in carbonate-bicarbonate buffer at a concentration of 50 μg/mL. Nunc 96-well ELISA plates (Corning, Millipore, Germany) were coated with 120 μl recombinant PS1 protein at 4°C overnight. After washing with PBST for 5 times, nonspecific binding sites were blocked with 3% BSA in PBS at 37°C for 1 h. After two washes with PBST, human plasma samples diluted at 1 : 100 with PBS were applied into the coated wells (100 μL/well) and incubated overnight at 4°C, with PBS as a negative control. A monoclonal antibody to PS1 (Abcam, USA) was used to validate peptide coating. The washing procedure was repeated three times, and horseradish peroxidase-conjugated goat anti-human IgG (H + L) antibody (Abcam, USA) (diluted at 1 : 5000 with PBS) was used to probe autoantibodies to PS1 in plasma. After three washes, substrate solution (TMB, Sigma, USA) (100 μL/well) was applied. After a 30-min incubation, the reaction was stopped using 1 M H2SO4 (100 μL/well) and the absorbance was read at 450 nm using an ELISA processor (ELISA Processor II; Dade Behring, Marburg, Germany). To ensure the consistency of the data generated from different Elisa plates, an internal reference plasma from a confirmed AD patient which showed relatively high optical absorbance in the preliminary test was used in each plate. If the OD value of a plasma sample in a plate was beyond the mean OD±2 SD of all plates, the sample would be re-measured. Samples were measured in duplicates, and the mean OD values of the duplicates were used for statistical analyses.
Statistical analysis
Demographic and clinical characteristics of participants were summarized using descriptive statistics, with continuous variables described as median/mean where appropriate and categorical data summarized as absolute frequencies and percentages. Comparative group p-values were determined via independent samples t-test and one-way analysis of variance (ANOVA). For three group analyses, post-hoc p-values were determined using Tukey’s adjustment for multiple comparisons. Spearman rank correlation analyses were conducted to examine the correlation of NAbs-PS1 with PET-Aβ SUVR, and with cognitive functions. Receiver operating curve (ROC) analyses provided the area under the curve (AUC) for differentiating subjects with different diagnoses.
RESULTS
Demographic information of participants
No intergroup difference was found in the mean age or median education level among groups. The non-AD CI group (71.43%) had a higher proportion of females than the CN group (42.85%). The AD group (45.45%) had more APOE ɛ4 carriers than the non-AD CI group (14.28%) and the CN group (11.42%). Both AD patients [median (IQR): 15 (10, 19)] and non-AD CI patients [median (IQR): 22 (18, 28.5)] had lower MMSE scores than CN [median (IQR): 27 (25, 29)]. AD patients [median (IQR): 2 (1, 2)] and non-AD CI patients [median (IQR): 1 (0.5, 1)] had higher CDR scores than CN [median (IQR): 0 (0, 0)]. Non-AD CI patients also had lower MMSE scores and higher CDR scores than CN. AD patients [median (IQR): 1.98 (1.79, 2.25)] had higher PET-PiB SUVR than non-AD subjects [median (IQR): 1.10 (1.05, 1.20)] (Table 1).
Characteristics of the Chongqing cohort
Measurement data is represented by n (%); numeration data is represented by mean±SEM or median (IQR) according to the normality. p-value, one-way ANOVA, two-tailed t-test, or Kruskal Wallis test as appropriate. APOE ɛ4, apolipoprotein E ɛ4 allele; MMSE, Mini-Mental State Examination; CDR, Clinical Dementia Rating; OD, optical density; SUVR, standard uptake value ratio. *p < 0.05 in comparison with CN. †p < 0.05 in comparison with AD.
Plasma NAbs-PS1 in subjects with different diagnoses
The relative levels NAbs-PS1 were determined using one-site ELISA. Amyloid-PET + subjects (AD patients) (mean±SD: 0.529±0.242) had significantly lower plasma NAbs-PS1 than Amyloid PET – subjects (including CN and non-AD CI subjects) (mean±SD: 0.741±0.223) (Fig. 1A). AD patients (mean±SD: 0.529±0.242) had significantly lower plasma levels of NAbs-PS1 than non-AD CI subjects (mean±SD: 0.753±0.209) and CN (mean±SD: 0.736±0.230). However, no difference was found in plasma NAbs-PS1 levels between non-AD CI patients and CN (Fig. 1B), suggesting a relatively specific decrease of NAbs-PS1 in AD.
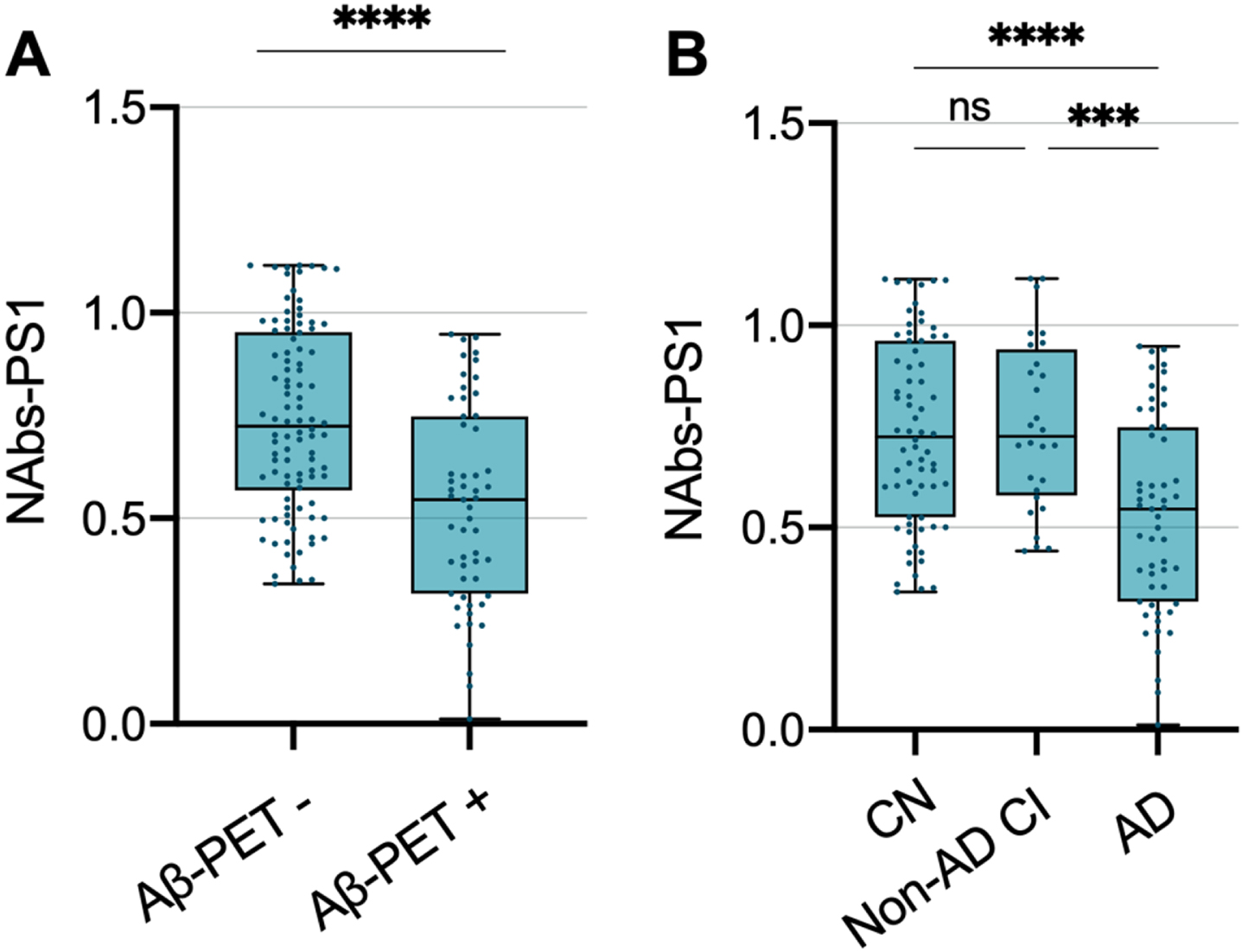
Plasma naturally occurring antibodies to presenilin-1 are decreased in AD patients. A) Plasma naturally occurring antibodies (NAbs-PS1) are decreased in Aβ-PET+ subjects, in comparison with Aβ-PET– subjects. Independent t-test. B) Plasma NAbs-PS1 are decreased in AD patients, in comparison with non-AD CI subjects and CN. Plasma NAbs-PS1 are not different between CN and non-AD CI subjects. One-way ANOVA. ns, no significant difference; CI, cognitive impairment. ***p < 0.001,****p < 0.0001.
Associations of plasma NAbs-PS1 with brain Aβ burden and cognitive functions
We found in this study that plasma NAbs-PS1 were negatively associated with brain Aβ burden, as reflected by PET-Amyloid SUVR (coefficient=– 0.533, p < 0.0001) (Fig. 2A). However, plasma NAbs-PS1 were positively associated with the cognitive functions of participants, as reflected by MMSE scores (coefficient = 0.307, p = 0.0002) (Fig. 2B). These findings suggested that plasma NAbs-PS1 were associated with the severity of AD.
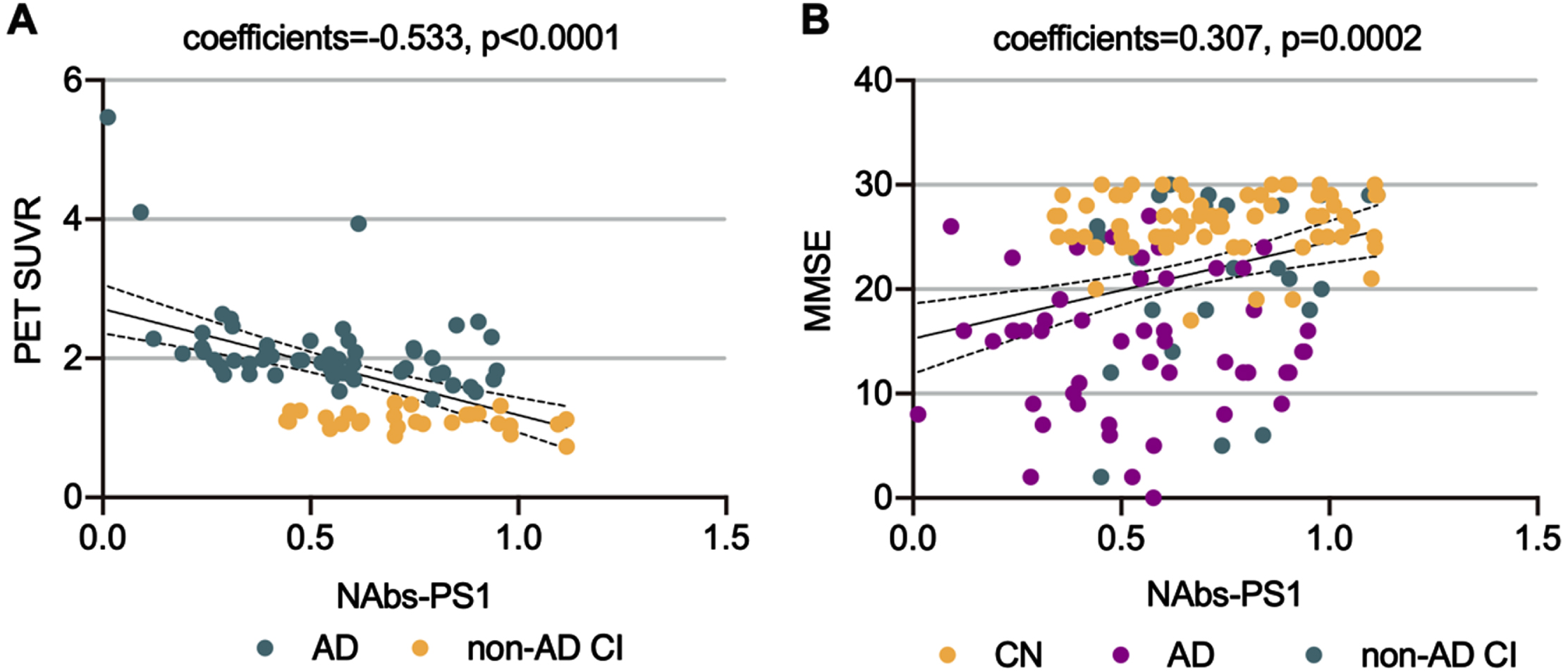
Associations of plasma NAbs-PS1 with brain Aβ burden and cognitive function. A) The association between plasma NAbs-PS1 and brain Aβ burden, as reflected by PET-Amyloid SUVR. Spearman correlation analysis. B) The association between plasma NAbs-PS1 and cognitive function, as reflected by MMSE scores. Spearman correlation analysis. CI, cognitive impairment.
Associations between plasma NAbs-PS1 with plasma AD-associated biomarkers
We next investigated the associations between plasma NAbs-PS1 with AD-associated biomarkers, including Aβ40, Aβ42, and total tau (tTau). No significant association was found of plasma NAbs-PS1 with either Aβ40 (coefficients = 0.003, p = 0.981), Aβ42 (coefficients = 0.068, p = 0.578), or tTau (coefficients=– 0.134, p = 0.271). However, a subgroup analysis suggested that plasma NAbs-PS1 were associated with tTau levels in the AD group (coefficients=– 0.289, p = 0.046) (Fig. 3A–C). These findings also imply that plasma NAbs-PS1 may be associated with the severity of AD.
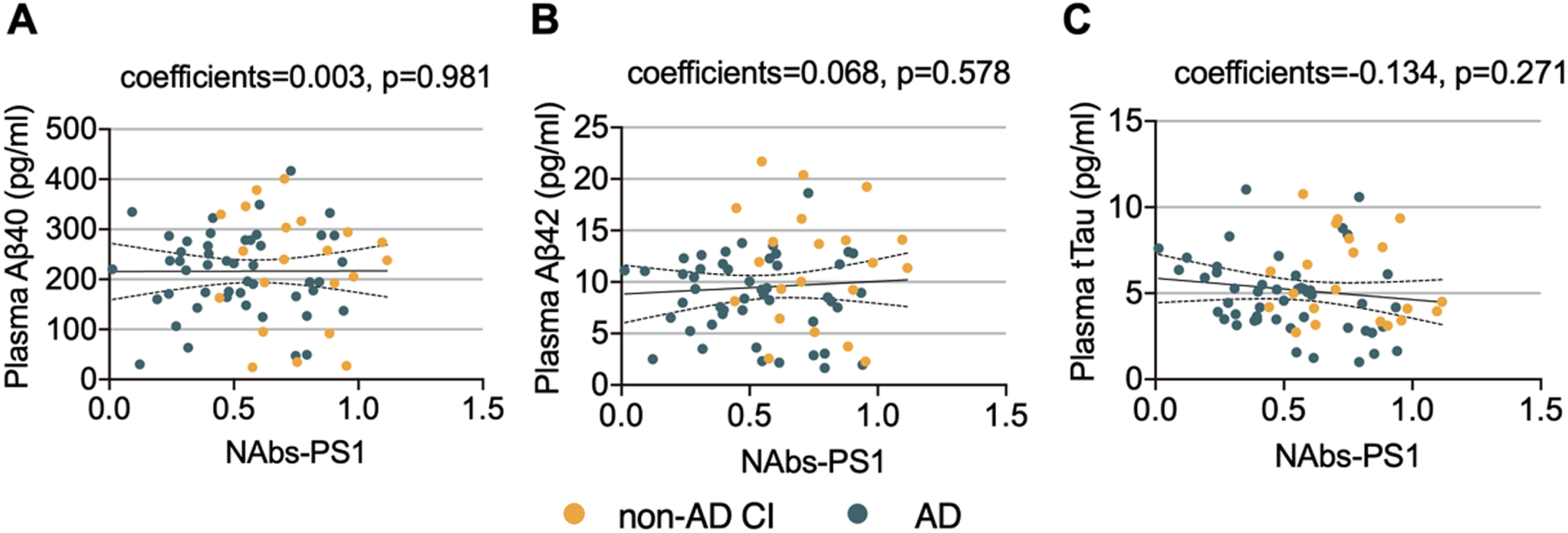
Associations between plasma NAbs-PS1 and AD-associated biomarkers. A) The association between plasma NAbs-PS1 and Aβ40 levels. Spearman correlation analysis. B) The association between plasma NAbs-PS1 and Aβ42 levels. C) The association between plasma NAbs-PS1 and tTau levels. Spearman correlation analysis. CI, cognitive impairment.
Discriminating value of NAbs-PS1 for subjects with different diagnosis
We then investigated how NAbs-PS1 discriminate subjects with different diagnoses. We found that plasma NAbs-PS1 discriminated AD patients from CN with an AUC of 0.730, a sensitivity of 69.09%, and a specificity of 67.14% (Fig. 4A). Plasma NAbs-PS1 discriminated AD patients from non-AD CI subjects with an AUC of 0.750, a specificity of 70.91%, and a sensitivity of 71.43% (Fig. 4B).
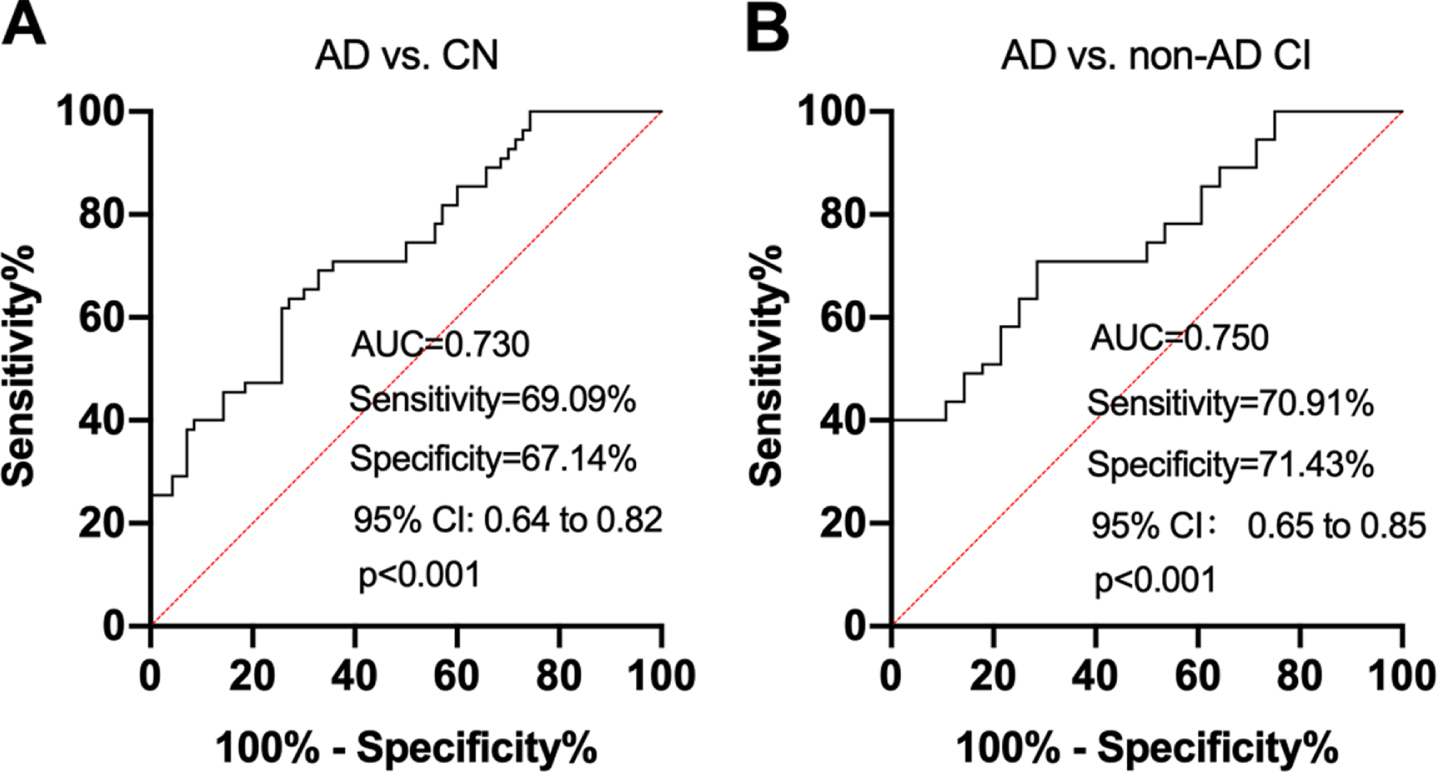
ROC analyses to test the ability of plasma NAbs-PS1 in differentiating subjects with different diagnoses. A) ROC analysis to test the ability of NAbs-PS1 in differentiating AD and CN. B) ROC analysis to test the ability of NAbs-PS1 in differentiating AD and non-AD CI subjects. ROC, receiver operating characteristic; AUC, area under the curve; CI, cognitive impairment.
DISCUSSION
In the present study, we found that plasma NAbs-PS1 were decreased in AD dementia patients, in comparison with CN and non-AD CI subjects. Furthermore, plasma NAbs-PS1 were associated with the severity of AD, as reflected by brain Aβ burden and cognitive functions. Plasma NAbs-PS1 discriminated AD patients from non-AD CI subjects and CN with acceptable accuracy.
A growing body of evidence has suggested that dysregulation of humoral immunity is a significant feature of AD. For incidence, biopsy studies identified an increased level of membrane-bound autoantibodies in the brain of AD patients, and the levels of autoantibodies are associated with the severity of brain amyloid pathology [16]. Profiles of circulating autoantibodies in AD patients are dramatically different from those of cognitively normal controls, and autoantibodies are potent in diagnosing AD [17]. Our group also identified several autoantibodies that are altered in AD and suggested that the alterations of these autoantibodies may contribute to the pathogenesis of AD [8, 19]. In recent years, clinical trials of AD immunotherapies have found that Aβ antibodies could induce AD-type pathologies, such as cerebral amyloid angiopathy, brain atrophy, and neuronal hyperactivity [20–26]. Taken together, these findings suggest an autoimmune element in the pathogenic network of AD.
The increase in the permeability of the blood-brain barrier during aging and the development of AD facilitates the exposure of brain antigens to peripheral immune cells and the infiltration of peripheral brain-reactive autoantibodies into the brain, and these pathological events may further promote the pathogenesis of AD [27–29]. Based on the present findings, it is hard to interpret whether the decrease of NAbs-PS1 is a subsequent or an upstream event of AD. During aging and the development of AD, the activities of BACE1 and γ-secretase are increased [2, 31]. Therefore, it is reasonable to speculate that the decrease in the levels of NAbs-PS1 might be due to the increased consumption of NAbs-PS1 by γ-secretase. The relative specificity of such an alteration in plasma NAbs-PS1 also supports this notion, as reflected by the unchanged NAbs-PS1 levels in non-AD CI subjects. In this study, we found that NAbs-PS1 discriminated AD from CN and non-AD CI with acceptable accuracy. As the sample size of this study is relatively low, this conclusion would be reinforced by further studies with larger sample sizes. The diagnostic accuracy of NAbs-PS1 for AD is not as high as recently discovered biomarkers, such as neurofilament light [32], pTau181 [33], and pTau 217 [34]. These biomarkers more reflect the neurodegeneration in the AT(N) framework [35], but NAbs-PS1 are involved in the etiological process of AD. Therefore, the association between NAbs-PS1 and longitudinal brain amyloidosis is worth investigating.
Recent advances in the development of β- and γ-secretase inhibitors yielded novel therapeutic strategies for AD [36, 37]. However, due to the off-target inhibition of multiple physiological functions of these secretases, the clinical trials of these inhibitors have not been successful [36]. Selective or partial inhibition of β-secretases by antibodies showed potential in the treatment of AD [38]. However, it is not clear yet whether the inhibition of γ-secretase by antibodies would also have therapeutic potential. If that were the case, we could speculate that NAbs-PS1 might be physiologically protective against AD through inhibiting the amyloidogenic processing of AβPP by γ-secretase, and thus the decrease of NAbs-PS1 would promote brain amyloidosis and the development of AD. However, inhibitors of β- and γ- secretases are not selective for AβPP and Notch, and thus the feasibility of NAbs-PS1 as a therapeutic target remains to be analyzed, perhaps better in an early intervention time window.
In conclusion, this study found an aberrant autoimmune phenotype in AD patients. This study has several limitations. First, this is a cross-sectional study based on comparative and correlative analyses. Further longitudinal investigations are of significance to demonstrate the association between NAbs-PS1 and the trajectories of AD from a clinical perspective. Second, this study lacks mechanistic investigations to further demonstrate the role of NAbs-PS1 in the pathogenesis of AD. Furthermore, as mentioned above, the diagnostic accuracy of NAbs-PS1 is not high enough to be a potential biomarker of AD, and this may need validation by larger sample-size studies. However, this study for the first time found altered levels and clinical relevance of NAbs-PS1 in AD and may provide some clues for future directions to investigate the immunological elements of AD pathogenesis.