
Abstract
Background:
Chronic disruption of the circadian timing system, often reflected as a loss of restful sleep, also includes myriad other pathophysiological effects.
Objective:
The current study examined how chronic circadian disruption (CD) could contribute to pathology and rate of progression in the AβPP/PS1 mouse model of Alzheimer’s disease (AD).
Methods:
A chronic CD was imposed until animals reached 6 or 12 months of age in AβPP/PS1 and C57BL/6J control mice. Home cage activity was monitored for a period of 3–4 weeks prior to the endpoint along with a single timepoint measure of glucose sensitivity. To assess long term effects of CD on the AD phenotype, animals were re-entrained to a no disruption (ND) schedule just prior to the endpoint, after which a Morris water maze (MWM) was used to assess spatial learning and memory.
Results:
Dampening of nighttime activity levels occurred in disrupted animals, and female animals demonstrated a greater adaptability to CD. Diminished arginine vasopressin (AVP) and vasoactive intestinal peptide (VIP) levels in the suprachiasmatic nucleus (SCN) of 12-month male AβPP/PS1 exposed to the CD paradigm were observed, potentially accounting for the diminished re-entrainment response. Similarly, CD worsened performance in the MWM in 12-month male AβPP/PS1 animals, whereas no effect was seen in females.
Conclusions:
Collectively, these findings show that exposure to chronic CD impairs circadian behavioral patterns and cognitive phenotypes of AβPP/PS1 mouse model in a sex-dependent manner.
Keywords
INTRODUCTION
The available evidence suggests a bidirectional relationship between dementia, including Alzheimer’s disease (AD), and circadian dysfunction [1, 2]. Historically, circadian dysfunction was considered a symptom of AD, with complaints of “sundowning”, increased daytime sleepiness, and nighttime restlessness. However, current literature suggests that circadian dysfunction could be an early marker of AD, even preceding notable cognitive deficits [3, 4]. Studies also suggest that the suprachiasmatic nucleus (SCN), the primary circadian oscillator, is a crucial mediator of circadian dysfunction resulting from dementia [5, 6]. Under conditions of normal aging, the SCN is comparatively resistant to neurodegeneration as compared to other brain regions, with SCN volume and neuronal number remaining relatively unaffected [7, 8]. In dementia however, arginine vasopressin (AVP), vasoactive intestinal peptide (VIP), and neurotensin containing neurons are first among those to be affected by neurodegeneration in the SCN [5, 9]. Receiving glutamatergic input from the retina and cholinergic input from sleep centers, this subset of SCN neurons is vital for synchronizing physiological rhythmicity to the external light cycle and in coordinating this information to regulate sleep timing.
In the brain, functional circadian clocks are present in astrocytes both inside and outside the SCN and considerable evidence suggests that disruption of clock-controlled genes impacts astrocyte function in AD and other neurodegenerative diseases [10–12]. Genetic models of circadian disruption (CD), such as Bmal1 null animals, present with severe astrogliosis, oxidative damage, and synaptic degradation [13, 14]. Similarly, non-genetic approaches that induce CD in animals lead to learning and memory deficits [15]. Animal models of jet-lag present learning and memory deficits, and human studies suggest that exposure to chronic CD is associated with significant hippocampal atrophy [16, 17]. Therefore, this study sought to examine whether chronic exposure to CD throughout disease development could potentially exacerbate or increase the rate of progression in an AβPP/PS1 mouse model of AD. Animals of both sexes were used in this study to determine if there were any sex differences in response to chronic CD.
The CD paradigm used in this study was designed to mimic human social behavior. The protocol involves a 7-day schedule, repeated weekly up to the 6- or 12-months of age endpoints that were assigned based on plaque initiation in the AβPP/PS1 model (6 months) and later stage AD development (12 months). We previously observed sex-dependent increases in nighttime home cage activity in AβPP/PS1 mice as compared to C57BL/6J mice, with significant increases seen at 6 months in males and 12 months in females [18]. The observed circadian differences in AβPP/PS1 mice prompted the question as to how disruption of the circadian system might differentially impact the AβPP/PS1 AD model as compared to C57BL/6J mice or how sex might be a factor in any observed differences. The CD protocol involves alteration of the lighting schedule such that the disrupted animals are phase delayed for days 1–5, then shifted to the same schedule as control animals on days 6–7; these alterations in the light cycle are repeated throughout the experiment. For simplicity, the paradigm can be compared to a person working night shifts during the week and attempting to rebound to a typical daytime schedule on the weekend. The circadian system is plastic to slight changes in the light-dark cycle; however, as is shown in this study and others, adaptation to drastic/sudden changes in the lighting schedule occurs gradually over consecutive circadian periods [19]. Experimentally, short-term CD paradigms are commonly used; however, this study for the first time exposes animals to chronic CD throughout AD progression, examining both the photoentrainment response and SCN physiology, as well as potential long-term effects on the AD phenotype.
METHODS
Animals
The Institutional Animal Care and Use Committee (IACUC) at Southern Illinois University School of Medicine approved animal use protocols. Eight-week-old male and female AβPP/PS1 (RRID: MMRRC 034832-JAX; Mo/HuAPP695swe/PS1-dE9) mice, as well as controls (C57BL6/J) (RRIF:IMSR JAX:000664), were bred locally, group housed and entrained to a no disruption (ND) 12:12-h light:dark schedule with food and water available ad libitum. Following entrainment to the ND schedule, half of the animals were introduced to the chronic CD paradigm. CD consists of a single 8-h phase delay of the time of lights on, followed by 5 days on that schedule, then a single 8-h phase advance to return them to the control schedule. After 2 days on the control schedule, the cycle was repeated for the duration of the experiment. Animals were left on either the ND or CD schedule until 6- or 12-month endpoints. One month prior to the endpoint, animals underwent Glucose Tolerance Test (GTT), then were individually housed and activity was monitored for 3 weeks using infrared-activity detectors (Minimitter, Bend, OR). Activity data were collected in 6-min bins and analyzed using Clocklab software (Actimetrics, Evanston, IL). Following activity monitoring, mice were entrained to the ND light/dark cycle and finally underwent a Morris water maze (MWM) cognitive assessment. Following MWM, all animals were sacrificed at Zeitgeber time (ZT) 12, defined as the time of lights off. Brain tissue was fixed in 4% paraformaldehyde for 24 h, then transferred to 30% sucrose for storage in 4°C.
Chemicals
All chemicals were prepared and stored according to manufacturer recommendations unless otherwise noted. Sodium chloride (Cat: 7647-14-5), Sucrose (Cat: 57-50-1), Sodium phosphate Dibasic, Anhydrous (Cat: 7558-79-4), Potassium chloride (Cat: 7447-40-7), Dextrose (Cat: 50-99-7), and Bovine serum albumin (Fraction V) (Cat: 9048-46-8) were obtained from Thermo Fisher Scientific (Waltham, MA). Potassium phosphate (Cat: 7778-77-01) was obtained from Sigma-Aldrich Co. (St. Louis, MO). Triton X-100 (Cat: 807426) and Sodium borohydride (Cat: 102894) was obtained from MP Biomedicals (Santa Ana, CA). Rabbit polyclonal VIP antibody (Cat: 20077) and rabbit polyclonal AVP antibody (Cat: 20069) were obtained from Immunostar (Hudson, WI). Chicken polyclonal glial fibrillary acidic protein (GFAP) antibody (Cat: C-1373-50) and Amylo-Glo®RTDtrademark (Cat: TR-300-AG) were obtained from Biosensis (Temecula, CA). Normal goat serum (Cat: S26-LITER) was obtained from Millipore (Burlington, MA). Goat anti-chicken IgY H&L (Alexa Fluor® 594) (Cat: ab150172) and Goat anti-rabbit IgG H&L (Alexa Fluor® 594) (Cat: A11012) was obtained from Invitrogen.
Glucose tolerance test
Following a 15-h fast, initial blood glucose measurements were performed at ∼ZT 12 (time = 0). A bolus of 20% glucose was subsequently administered intraperitoneally (IP) at 2 g of glucose/kg body weight (b.w.). Blood glucose levels were tested at 15, 30, 45, 60, and 120 min using a Contour Next EZ Glucometer (Parsippany, NJ).
Morris water maze
Mice underwent cognitive assessment of spatial learning and memory recall using MWM at either the 6- or 12-month endpoint. The MWM test requires the animals to utilize distinct visual clues placed around the room and around the maze borders to find the location of a static, hidden platform (submerged 1 cm below an opaque water surface) across repeated trials [20, 21]. The MWM paradigm consists of 5 consecutive training days (3×90 s trials/day) with a minimum inter-trial-interval of 20 min. Each trial’s starting quadrant was randomized among three starting locations, with no location repeated in a single day. Two days post-training, mice underwent a 60-s probe trial from a singular starting location to determine memory-recall. Annulus 40 was defined as a region 40 cm in diameter centered around the platform, which is different from the annulus zone used to evaluate search strategies in annular analysis of MWM. Animal navigation during the challenge and probe trials was recorded using ANY-maze video tracking system (Stoelting Co., Wood Dale, IL; RRID:SCR 014289). The three trials for each training day were averaged for each animal in all analytical parameters.
Immunofluorescence (IF) and analysis
Brain samples were fixed in 4% paraformaldehyde for 48 h and then transferred into 30% sucrose in 0.1M phosphate buffer for 24 h prior to sectioning. 20μ SCN and hippocampal sections were collected serially using a cryostat (Model HM525 NX, ThermoFisher Scientific). Every fourth section in the SCN sections underwent IF using rabbit polyclonal VIP antibody or rabbit polyclonal AVP antibody. PBST (0.1M PBS with 0.25% TritonX-100) was used to permeabilize the sections followed by 3×10-min washes in sodium borohydride in PBS (1 mg/ml) for antigen retrieval. Sections were washed 3×10 min with PBST and blocking was done using 10%, normal goat serum/1% BSA for 1 h. Sections were then incubated overnight in primary antibody at 4°C. The next day, sections were washed 4×10 min in PBST and incubated in secondary antibody Goat anti-rabbit IgG H&L (Alexa Fluor® 594) 1:1000 for 2 h, washed 4×10 min in PBST and coverslipped using ProLongtrademark Gold antifade reagent with DAPI. Every 6th section in the hippocampus underwent IF using Chicken polyclonal glial fibrillary acidic protein (GFAP) antibody 1:1000, Amylo-Glo®RTDtrademark 1:100. Prior to GFAP staining, slides were immersed in a 70% ethanol solution for 5 min before being rinsed in PBS for 2 min. Fresh 1:100 dilution of 100X stock Amylo-Glo RTDTM was prepared, and the sections were incubated in the staining solution for 10 min. Slides were then rinsed in 0.9% saline solution for 5 min without shaking and prepared for IHC by rinsing 3×2 min in PBS. The secondary antibody used was Goat anti-chicken IgY H&L (Alexa Fluor® 594) 1:1000. Amylo-Glo/GFAP slides were coverslipped using ProLong Goldtrademark antifade reagent. The section images were captured using Nikon Eclipse E-600 microscope equipped with an Olympus-750 video camera system. Staining intensities for VIP, AVP, and GFAP were measured using National Institute of Health Image J Software 1.48 (RRID:SCR_003070). Staining density was obtained when background staining was subtracted from mean intensities. Amylo-Glo RTDTM stain was analyzed using the same equipment, however measurements were calibrated in Image J software and recorded in mm2.
Data analysis
Prism 10.0 (GraphPad Software, Inc., La Jolla, CA; RRID:SCR_002798) software was used for statistical analyses. Two-way ANOVA was used for locomotor activity analysis, plaque number, MWM corrected integrated path length (CIPL) and MWM cumulative distance. Mann-Whitney test was used to analyze fasting glucose, MWM time spent in annulus, as well as AVP and VIP fluorescence. GraphPad Prism software calculated the area under the curve measurements for the glucose tolerance test. Data are represented as mean±SEM and statistical significance was defined as p < 0.05.
RESULTS
Sex-dependent re-entrainment patterns in AβPP/PS1 mice after chronic CD
Representative actograms are shown for 12-month animals under the CD lighting schedule the ND lighting schedule (Fig. 1). Total activity was determined from infrared detection of activity in home cages. All groups show alignment to the light/dark cycle on the ND schedule. C57BL/6J males and females, and AβPP/PS1 females demonstrate alterations in activity on the CD schedule (Fig. 1). Total activity was not different across genotype or lighting schedule in 6-month females. At 12 months, two-way ANOVA indicated a significant main effect of genotype (F1,63 = 4.244, p = 0.0435), with ND AβPP/PS1 females showing an increase in activity (Fig. 2A) compared to ND c57BL/6J. In males, there was a significant main effect of genotype at 6 months (F1,62 = 6.239, p = 0.0152), with ND and CD AβPP/PS1 males showing increased activity compared to the c57BL/6J groups (Fig. 2B).

CD and ND lighting regimen and representative actograms. Representative, single-plotted actograms for activity in 12-month ND and CD mice. Shaded bars in the actograms indicate the time of lights off. Time of day, on the X axis, is shown as actual clock time.
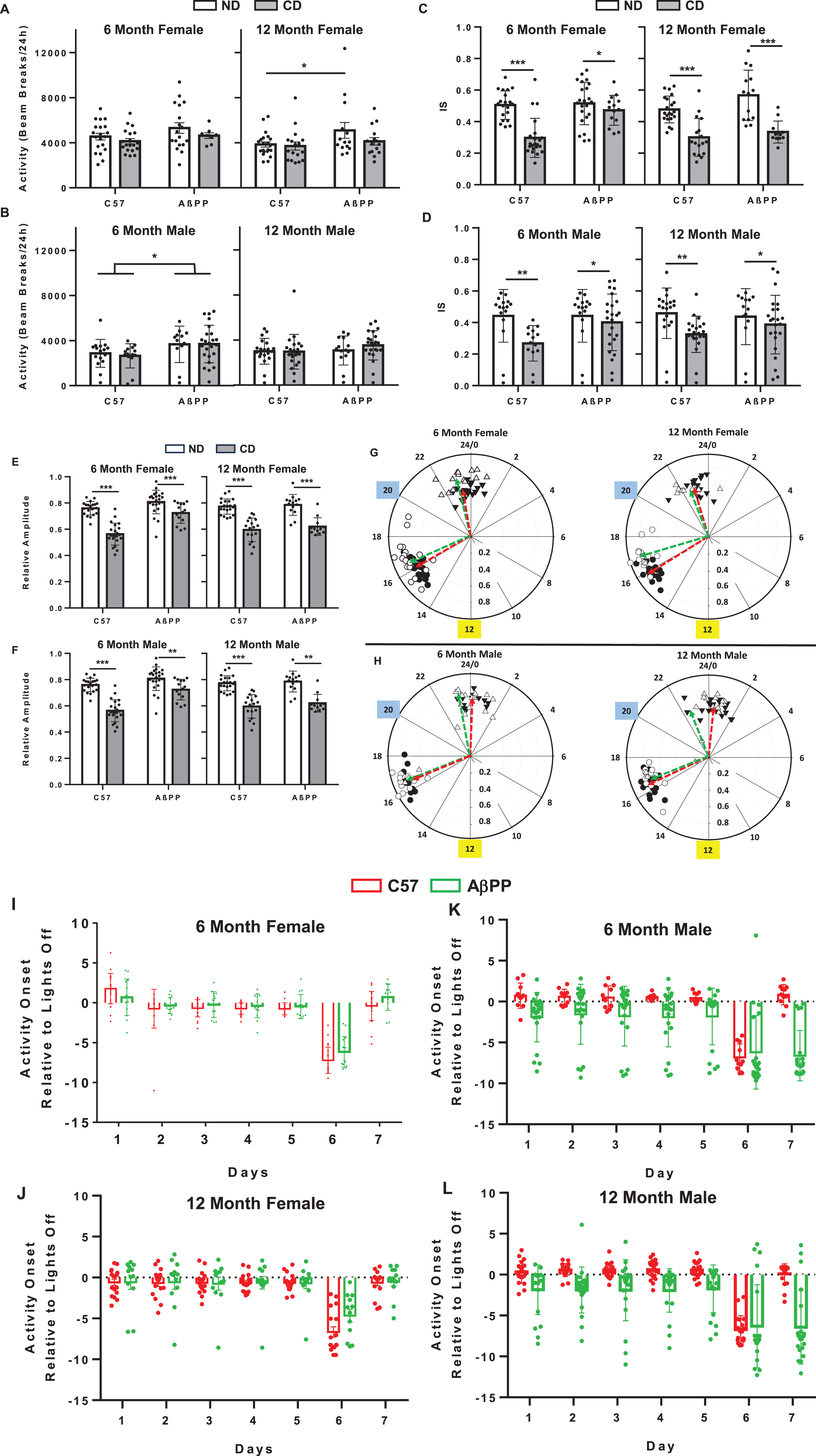
Diurnal locomotor activity levels in C57BL/6J and AβPP/PS1 mice under conditions of circadian disruption. A) Average locomotor activity during the lights on and lights off period in a 6-month and 12-month female C57BL/6J and AβPP/PS1 mice on either the ND or CD lighting schedule as measured via breakage of a cage-mounted infrared beam. B) 6-month and 12-month male C57BL/6J and AβPP/PS1 mice on either ND or CD lighting as measured via breakage of a cage-mounted infrared beam. C) IS of activity for 6-month and 12-month female C57BL/6J and AβPP/PS1 mice. D) IS of activity for 6-month and 12-month male C57BL/6J and AβPP/PS1 mice. E) Relative Amplitude of Activity for 6-month and 12-month female C57BL/6J and AβPP/PS1 mice. F) Relative Amplitude of Activity for 6-month and 12-month male C57BL/6J and AβPP/PS1 mice. G, H) Polar phasor plots of phase of mean peak activity on the angular axis (in 24-h time) and relative amplitude on the radial axis. Yellow box indicates time of lights off in the ND group. Blue box indicates time of lights off during days 1–5 in the CD group. Green vectors indicate AβPP/PS1 mice and red vectors indicate C57BL/6J mice. Black circles are C57BL/6J ND, black triangle are C57BL/6J CD, white circles are AβPP/PS1 ND, and white triangles are AβPP/PS1 CD. I) Time of activity onset in 6-month and 12-month female C57BL/6J and AβPP/PS1 mice housed in CD lighting conditions. J) Time of activity onset in 6-month and 12-month male C57BL/6J and AβPP/PS1 mice housed in CD lighting conditions. Data are represented as mean±SEM. Activity data were analyzed by 2-way ANOVA using GraphPad Prism 10. *p < 0.05; **p < 0.005; ***p < 0.0005.
To explore the effects of CD on diurnal activity characteristics, non-parametric circadian rhythms analysis was used [22, 23]. Interdaily stability (IS) is a measure of the consistency of daily activity patterns across days, with a value of 1 showing high stability, and 0 being no stability. There were no genotype or sex differences in mean IS among groups within the same lighting schedule. At 6 and 12 months, females showed a significant main effect of CD (6 mo, F1,75 = 22.58, p < 0.0001; 12 mo F1,60 = 50.67, p < 0.0001), with CD females of both genotypes showing reduced IS, as expected (Fig. 2C). CD also reduced IS in 6- and 12-month males (6 mo, M1,56 = 7.406, p = 0.0083; 12 mo F1,75 = 6.162, p = 0.0153; Fig. 2D).
Relative amplitude (RA) is a non-parametric method to express the differences between periods of peak activity and rest (rhythm amplitude) based on comparing the periods of highest and lowest activity. RA = (average high activity-average low activity)/total activity. Values range from 0-1 with 1 being a high amplitude and 0 being arrhythmicity. In females, there was a significant main effect of CD (6 mo, F1,75 = 61.89, p < 0.0001; 12 mo F1,60 = 82.32, p < 0.0001, but no effect of genotype or sex (Fig. 2E). Similarly, in males at both time points, there was a significant main effect of CD (6 mo, F1,50 = 16.36, p = 0.0002; 12 mo F1,63 = 21.88, p < 0.0001, but no effect of genotype or sex (Fig. 2F). These data indicate that CD reduced rhythm amplitude.
Phasor plots (Fig. 2G, H) were used to represent the relationship between the average daily time when peak activity occurred and the robustness, as indicated by relative amplitude. Read like a 24-h clock face, the average of the time when peak activity occurred is represented on the angular axis. The length of the vector (arrow), which indicates the amplitude, or robustness of the rhythm, is shown on the radial axis. In all groups, the mean time of peak activity was significantly delayed by CD (occurs later in the 24-h day). In 6- and 12-month females, mean time of peak activity was significantly delayed in ND AβPP/PS1 (6 mo CT16.57±0.68; 12 mo CT16.96±0.68; Fig. 2G green vectors) mice compared to C57BL/6J (6 mo 16.06±0.62; 12 mo 15.9±0.47; 6 mo p = 0.045; 12 mo p < 0.001, Fig. 2G red vectors) ND control mice; there was no difference between genotypes in the CD groups (Fig. 2G green vectors). In males at both ages, there was no significant difference in mean time of mean activity in ND groups (Fig. 2H green vectors); however, mean time of peak activity occurred earlier in AβPP/PS1 males at 12 months (12 mo CT22.94±2.97, Fig. 2H green vectors) compared to C57BL/6J (24.39±1.04; p = 0.042, Fig. 2H red vectors). For AβPP/PS1 males, there was an increase in variability among animals in the CD group, with some animals maintaining timing of peak activity more similar to the ND group (note white triangles in the polar plots).
Time of activity onset relative to the time of lights off in the lighting cycle was explored as a measure of whether animals attempted to adjust their activity to the altered light cycle presented on days 6 and 7 of each week. For 6- and 12-month females, there was main effect of time (6 mo F6,238 = 84.48, p < 0.0001; 12 mo F6,189 = 22.93, p < 0.0001). There was no effect of genotype. On day 6, the day the lights were altered, onset in all female groups was ∼8 h prior to the new lights off, consistent with the lights off time before the change. On the second day of the altered schedule, all female groups had adjusted to the altered schedule (Day 7). Upon return to the day 1–5 schedule, all females quickly adjusted. For males, there was a significant main effect of time (6 mo F6,210 = 16.07, p < 0.0001; 12 mo F6,259 = 26.32, p < 0.0001) and genotype (6 mo F1,210 = 38.88, p < 0.0001; 12 mo F1,259 = 60.25, p < 0.0001) and an interaction of genotype with time in both age groups (6 mo F6,210 = 4.15, p = 0.002; 12 mo F1,259 = 5.347, p < 0.0001). C57BL/6J males were similar to females; they showed misalignment with the light/dark cycle after the lighting switch on day 6 but adjusted to the lighting schedule by day 7. AβPP/PS1 males did not adjust on day 7 of the lighting schedule; activity onset remained consistent with the day 1–5 schedule. There was greater variability in the male AβPP/PS1 groups. Some animals did attempt to adjust to the new light/dark cycle, whereas others did not.
Metabolic changes associated with CD
To determine baseline diurnal patterns in metabolic function, around the clock fasting glucose measurements were taken from C57BL/6J animals (Fig. 3A). Fasting glucose measurements were taken across different days with the same 15-h fast prior to the measurement. Animals for this experiment were 4- to 10-month males and females. In males, fasting glucose levels increased from ZT4 and peaked around ZT12, falling back to baseline by ZT16. In females, fasting glucose peaked earlier, at ZT 10 and returned to baseline by ZT 16.
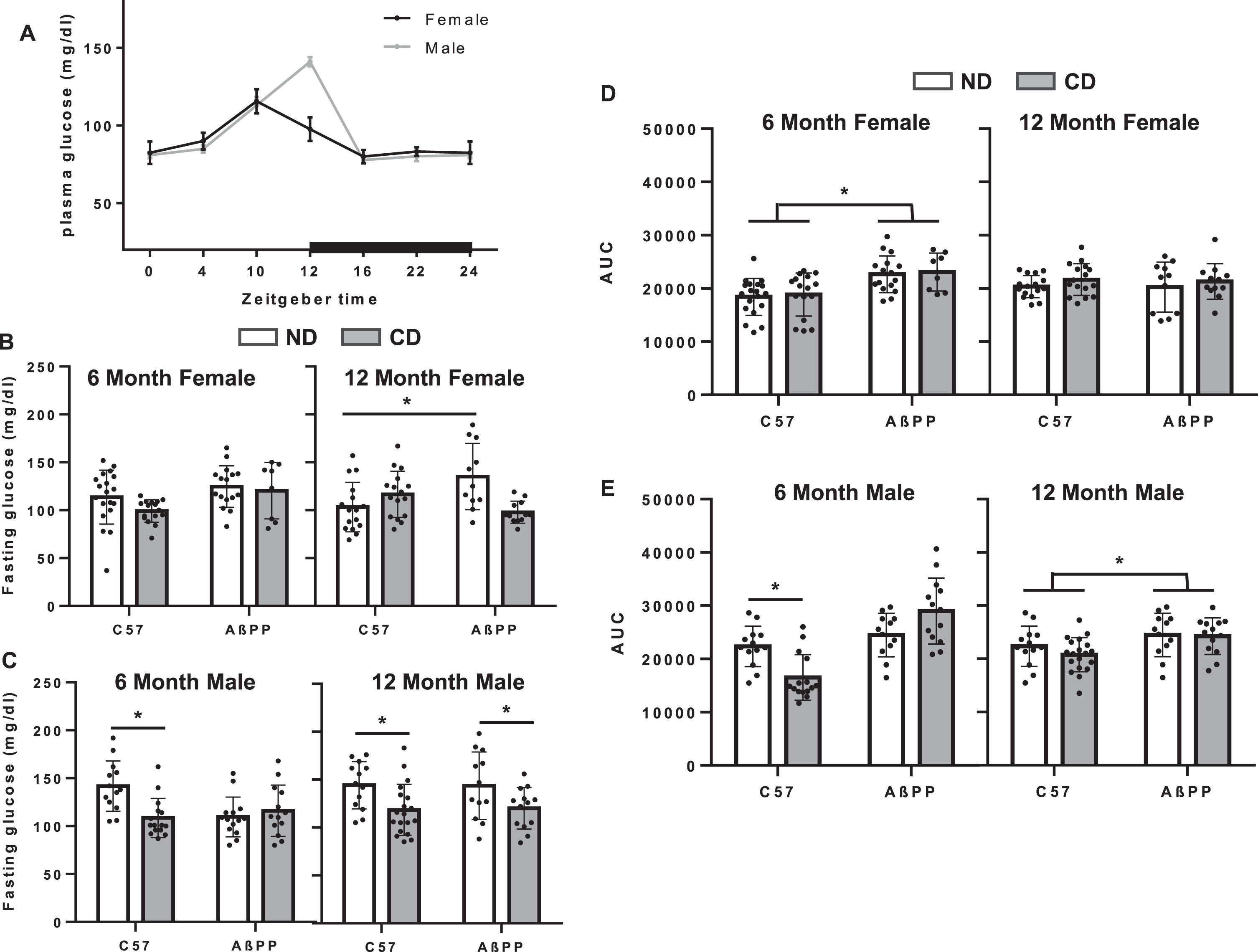
Fasting glucose and GTT AUC in C57BL/6J and AβPP/PS1 mice under conditions of CD. A) Fasting glucose levels following a 15-h fast at ZT 4, 8, 12, 16, and 20 in 4–10-month male and female C57BL/6J mice. B) Fasting glucose in female 6-month and 12-month C57BL/6J and AβPP/PS1 mice on either the ND or the CD lighting schedule. C) Fasting glucose in male 6-month and 12-month C57BL/6J and AβPP/PS1 mice on either the ND or the CD lighting schedule. D) Glucose tolerance as measured via area under the curve of blood glucose across 120 min following a bolus of 20% glucose administered i.p. in 6-month and 12-month female C57BL/6J and AβPP/PS1 mice. E) Glucose tolerance as measured via area under the curve of blood glucose across 120 min following a bolus of 20% glucose administered i.p. in 6-month and 12-month female C57BL/6J and AβPP/PS1 mice. Data are represented as mean±SEM. Fasting glucose and AUC data analyzed by 2-way ANOVA with GraphPad Prism. *p < 0.05, **p < 0.05.
GTTs were performed 3–5 weeks prior to sacrifice. Time = 0 for GTT was 6:45 AM on day 7 of either the control or CD schedule, which for ND animals was at approximately ZT9. Fasting glucose levels were determined from time = 0 measurements and average fasting glucose levels were plotted for each cohort. In females, there was a significant main effect of genotype at 12 months (F1,98 = 6.401, p = 0.0143, Fig. 3B), with ND AβPP/PS1 females showing an increase in fasting glucose, though no significant interaction was observed. In males (Fig. 3C), there was a significant effect of ND on fasting glucose in 6-month c57BL/6J (F1,49 = 32.07, p < 0.001) and in both genotypes at 12 months (F1,51 = 8.398, p = 0.0055), with ND animals showing lower fasting glucose. Six-month female AβPP/PS1 animals presented higher AUC measurements as compared to treatment matched C57BL/6J 6-month female mice (Fig. 4D, F1,55 = 17.92, p < 0.0001). Six-month CD C57BL/6J males presented significantly lower AUC measurements as compared to ND CC57BL/6J male mice at 6 months (Fig. 3E, F1,49 = 8.19, p = 0.02). At 12 months, there was a significant main effect of genotype (F1,53 = 8.398, p = 0.015), but no effect of CD.
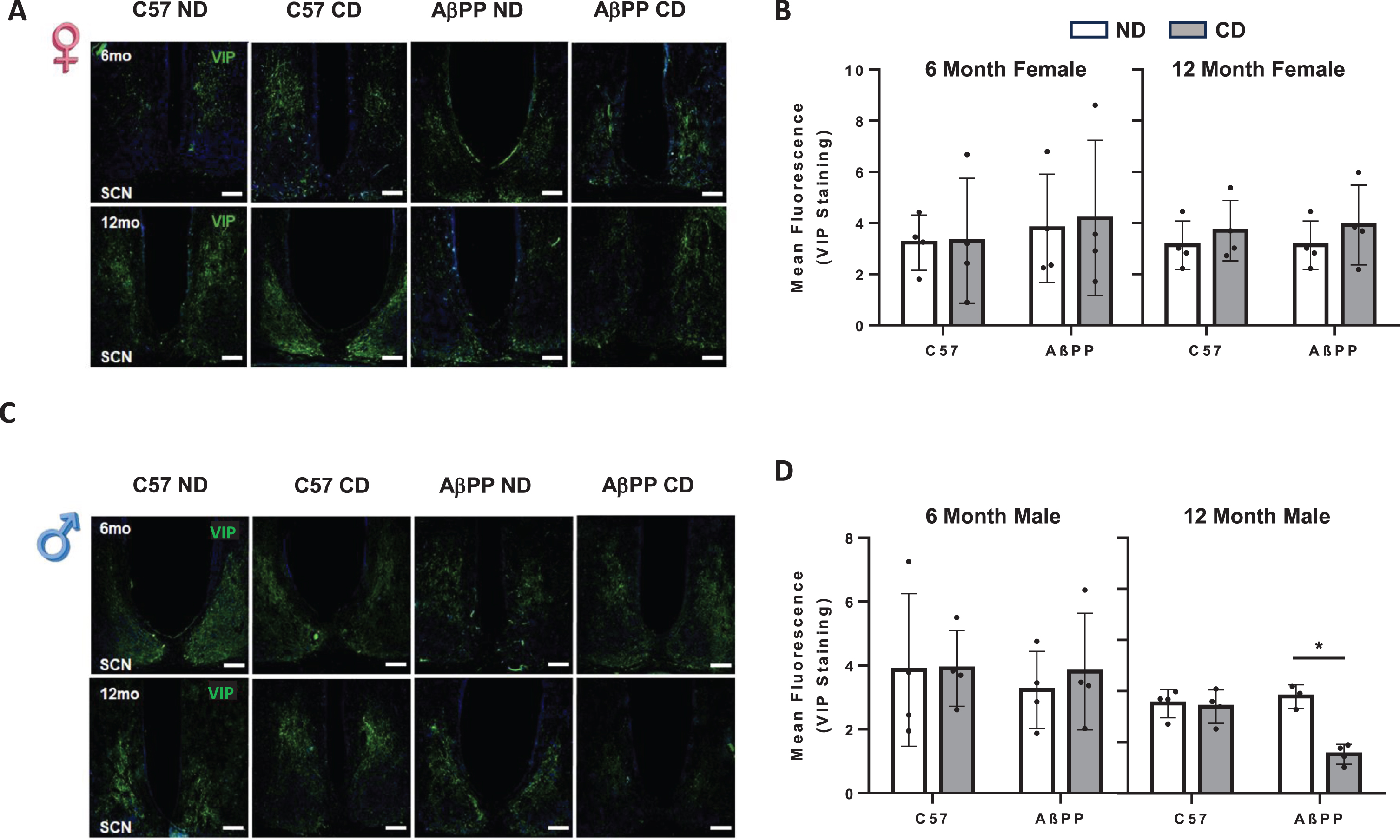
SCN VIP expression is unchanged in response to chronic CD in C57BL/6J and AβPP/PS1 mice. A) Representative VIP images from the SCN of 6-month (top) and 12-month (bottom) female C57BL6J and AβPP/PS1 mice exposed to ND and CD lighting schedules. B) Mean fluorescent intensity for VIP expression in 6-month and 12-month female C57BL6J and AβPP/PS1 exposed to ND lighting schedule CD lighting schedule. C) Representative VIP images from the SCN of 6-month (top) and 12-month (bottom) male C57BL6J and AβPP/PS1 mice exposed to ND and CD lighting schedules. VIP is shown in green, DAPI stain is shown in blue. N = 3-4 for all groups. Data are represented as mean±SEM. Data were analyzed by Two-way ANOVA with Graphpad Prism software. *p < 0.05. Scale Bar represents 100μm. All images were taken at 10x magnification.
Chronic CD resulted in reduced AVP and VIP expression in the SCN of male AβPP/PS1 mice
Differences observed in re-entrainment patterns between male and female AβPP/PS1 animals prompted an examination of AVP and VIP levels in the SCN, which are critical for integrating light information into the SCN and transmitting cues that result in behavioral changes that allow alignment to external lighting cues. Coronal sections were selected to represent the approximate midpoint of the SCN. In females, there was not a significant difference in VIP expression as a result of chronic CD at either 6 months (F1,11 = 0.4117, p = 0.5342) or 12 months (F1,12 = 0.01667, p = 0.8994) in C57BL/6J animals or AβPP/PS1 animals (Fig. 4A, B). In 6-month males, there were no significant changes in VIP expression after CD in either C57BL/6J or AβPP/PS1 mice (F1,12 = 0.0064, p = 0.9375). In 12-month males, there was a significant main effect of treatment groups between genotypes on VIP expression (F3,10 = 8.179, p = 0.0048). In 12-month males, there was no significant difference in VIP expression in CD C57BL/6J mice as compared to ND mice. There was a significant effect of genotype (F1,11 = 8.329, p = 0.02) and CD (F1,11 = 18.73 p = 0.0012) and a significant interaction between genotype and CD (F1,11 = 15.11, p = 0.0025). In AβPP/PS1 12-month males, CD resulted in significantly lower VIP expression in the SCN as compared to ND AβPP/PS1 12-month males (Fig. 4C, D).
Similar results were found in AVP expression. No significant differences were observed as a result of chronic CD in females of either age or genotype (Fig. 5A, B). Six-month males also did not show any significant differences in AVP expression in either C57BL/6J or AβPP/PS1 animals. In 12-month males, however, AVP expression was significantly decreased in CD AβPP/PS1 as compared to ND AβPP/PS1 mice (F1,11 = 6.326, p = 0.0112) (Fig. 5C, D).
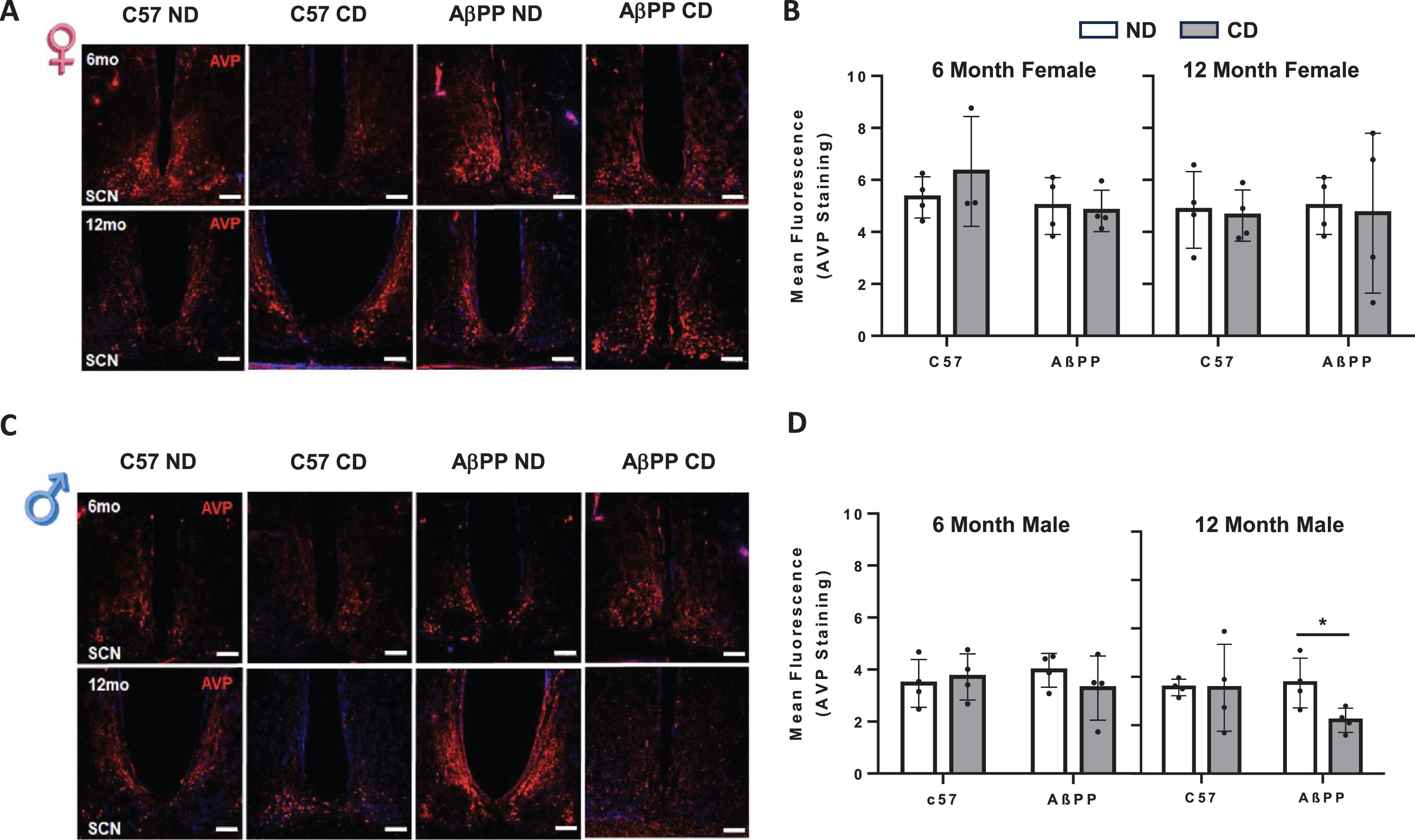
SCN AVP expression is unchanged in response to chronic CD in C57BL/6J and AβPP/PS1 mice. A) Representative VIP images from the SCN of 6-month (top) and 12-month (bottom) female C57BL6J and AβPP/PS1 exposed to ND and CD lighting schedules. B) Mean fluorescent intensity for VIP expression in 6-month and 12-month female mice. C) Representative VIP images from the SCN of 6-month (top) and 12-month (bottom) male C57BL6J and AβPP/PS1 exposed to ND and CD lighting schedules. D) Mean fluorescent intensity for VIP expression in 6-month and 12-month male mice. VIP is shown in green, DAPI stain is shown in blue. N = 3-4 for all groups. Data are represented as mean±SEM. Data were analyzed by Two-way ANOVA with GraphPad Prism. *p < 0.05. Scale Bar represents 100mμm. All images were taken at 10x magnification.
Chronic CD impacted cognition in male AβPP/PS1 mice
Following the activity monitoring period, all animals were re-entrained to the same 12:12-h light/dark schedule for a period of 3–5 weeks prior to the MWM task. Maintaining animals on the same lighting schedule regardless of initial treatment group ensured that any effects on cognition were lasting effects of chronic CD throughout disease progression, as opposed to animals being at a different point in their respective circadian period upon task initiation. An 8-day MWM paradigm was used to assess spatial learning and memory. In 12-month female animals, there was no significant impact of CD on spatial learning in either C57BL/6J or AβPP/PS1 mice. Platform:CIPL and cumulative distance to the platform during the 5-day training phase, were significantly different between genotypes (F3,222 = 6.312, p = 0.0004) and trial day (F5,222 = 33.37, p < 0.0001) with no significant interaction between the two on platform: CIPL measurements. Significantly higher CIPL values were observed for ND 12-month female AβPP/PS1 as compared to ND 12-month female C57BL/6J on days 3 and 5, indicating impaired spatial learning in AβPP/PS1 12-month females; however, there was no detrimental effect of chronic disruption (Fig. 6A, B). Total time spent in the annulus 40 during the probe trial was used to assess long-term spatial memory. In 12-month females, there was no significant effect of CD in either C57BL/6J or AβPP/PS1 mice in performance during the probe trial (Fig. 5C-E). No effect of CD on spatial learning and memory was observed in 6-month female AβPP/PS1.

Spatial learning in AβPP/PS1 12-month mice exposed to chronic CD. CIPL and cumulative distance from the platform were averaged across 3 independent trials per day during the 5-day training period. Probe (day 8) data was taken from a single 60-s trial. A) Platform: CIPL during the 5-day training period is represented in 12-month females. B) Cumulative distance from the platform is represented in 12-month females. C) Platform: CIPL during the probe trial is represented in 12-month females. D) Cumulative distance from the platform during the probe trial is represented in 12-month females. E) Total time spent in the annulus 40 during the probe trial is represented in 12-month females. F) Platform: CIPL during the 5-day training period is represented in 12-month males. G) Cumulative distance from the platform is represented in 12-month males. H) Platform: CIPL during the probe trial is represented in 12-month males. I) Cumulative distance from the platform during the probe trial is represented in 12-month males. J) Total time spent in the annulus 40 during the probe trial is represented in 12-month males. Sample sizes are noted in each respective figure. Data are represented as mean±SEM. Platform: CIPL data were analyzed by 2-way ANOVA with GraphPad Prism. $p < 0.05 C57BL/6J ND versus AβPP/PS1 NS. #p < 0.05 C57BL/6J CD versus AβPP/PS1 PS. **p < 0.01 AβPP/PS1 ND versus AβPP/PS1 PS. *p < 0.05.
Chronic CD had a significant impact on cognition and memory in 12-month AβPP/PS1 males; however, chronic CD had no effect at the 6-month endpoint. There was a significant main effect of treatment groups between genotypes (F3,240 = 16.90, p < 0.0001) and trial day (F5,240 = 40.10, p < 0.0001) with no significant interaction between the two on platform: CIPL measurements. On day 1, 2 and day 4 of the 5-day training phase, CD AβPP/PS1 mice presented significantly higher platform: CIPL values as compared to ND AβPP/PS1 mice. By day 5, there was no significant difference in platform: CIPL values between groups, indicating that though all groups were able to learn the platform location by day 5, CD AβPP/PS1s demonstrated significantly slower learning patterns than ND AβPP/PS1s (Fig. 6F, G). There was no significant impact of chronic disruption on spatial learning in C57BL/6J mice. In the probe trial there was a significant main effect of treatment groups between genotypes on time spent in the annulus 40 (F3,46 = 3.328, p = 0.0276). 12-month male CD AβPP/PS1 animals spent significantly less time in the annulus 40 as compared to age and sex-matched ND AβPP/PS1 and CD C57BL/6J animals, indicating impaired spatial long-term memory (Fig. 6J). Similarly, there was a significant main effect of treatment groups between genotypes (F3,240 = 16.95, p < 0.0001) and trial day (F5,240 = 38.15, p < 0.0001) with no significant interaction between the two on cumulative distance from the platform. Cumulative distance from the platform was significantly higher in CD AβPP/PS1 as compared to ND AβPP/PS1 in 12-month males (Fig. 6I).
Chronic CD does not impact the AD phenotype in AβPP/PS1 mice
In female mice, there was no significant impact of chronic CD on average hippocampal plaque diameter (Supplementary Figure 1), plaque diameter (Supplementary Figure 2) or GFAP fluorescence (Supplementary Figure 3) in AβPP/PS1 mice at either the 6- or 12-month endpoints. Similarly, in male AβPP/PS1 animals, there was no effect of CD on plaque number across all regions of the hippocampus after CD at 6 and 12 months. Average plaque diameter (Supplementary Figure 1) and GFAP expression was also not changed (Supplementary Figure 4).
DISCUSSION
Chronic CD was induced via recurrent alterations in the lighting schedule throughout the entire experimental period, designed to mimic human social behavior or work-related shift work schedules. Chronic CD altered behavioral patterns in activity, glucose metabolism, and performance in the MWM learning and memory task, in a sex-specific manner. As reported previously [18], AβPP/PS1 mice showed as increase in overall locomotor activity that was sex- and time-specific; increase activity occurred in 6-month males and 12-month females. Actogram data for animals exposed to CD clearly indicate altered patterns of circadian locomotor activity. Actograms clearly show that females of both genotypes, as well as c57BL/6J males, adjust their activity in response to the altered lighting schedule on days 6 and 7. AβPP/PS1 males showed a range of responses to the change in light/dark cycle, but generally fail to adjust to the weekend schedule, with some animals not even attempting to change. CD reduced the relative amplitude (Fig. 2E, F) and interdaily rhythm stability (Fig. 2C, D) in all animals, suggesting that the CD paradigm chosen did have disruptive effects on the circadian system of the rhythm in all animals, which is consistent with previous studies using different CD methods [24, 25]. However, genotype effects of CD were different in males and females. The baseline phase (time of average peak activity) of activity occurred later in AβPP/PS1 females in ND groups compared to age-matched c57BL/6J females; differences were increased at 12 months.
Non-parametric measures indicated sex-specific changes in circadian parameters in response to CD. Subsequently, we explored alignment of activity onset to the light/dark cycle with respect to day of the week to determine whether CD animals we attempting to align with the alterations in light timing. Regardless of lighting schedule, ND and CD mice were aligned to their respective light/dark cycle on days 2–4 of the week. Activity onset for all groups on these days occurred slightly before the time of lights off, indicating that all animals were able to anticipate the time of lights off. On day 6, the first day of the subjective weekend, activity onset for all groups of animals was not aligned with the onset of lights off, rather, they became active significantly earlier, at a time that aligned with lights off on the previous day. On day 7, activity onset for all females and c57BL/6J males moved closer to the time of lights off, indicating an attempt to align with the new light/dark cycle. However, AβPP/PS1 males became active significantly earlier, similar to day 6, suggesting an inability to adjust to the altered light/dark cycle. The lack of attempt to adjust to the weekend light schedule may indicate that the male brain is not responding to the change in lighting schedule, which may reflect sex differences in SCN function. A diminished re-entrainment response to multiple insults has been recorded previously in mice, which is suggested to represent a coping mechanism during periods of repeated exposure to changing light schedules [26, 27].
Sex differences in the re-entrainment response to the 8-h phase advance going into the subjective weekend in AβPP/PS1 animals prompted an exploration of the SCN. The SCN receives glutamatergic input onto VIP containing neurons via the retinal hypothalamic tract to synchronize rhythms to the environmental 24-h light/dark cycle [28, 29]. These VIP-containing neurons in turn act to provide inter-neuronal synchronization within the SCN to strengthen overt circadian output [30, 31]. Within the SCN, AVP containing neurons participate in generating robust circadian rhythms and the determination of circadian period, primarily via inter-neuronal coupling [28, 30–32]. A previous study from our laboratory showed that neither AVP or VIP expression was significantly changed from 6–12 months of age in AβPP/PS1 mice of either sex [18]. In postmortem human studies, both AVP and VIP expression was significantly decreased in patients with AD as compared to age-matched controls. Additionally, it was shown that in normal aging, VIP expression was unaffected in females; however, decreased significantly with age in males. In a triple transgenic mouse model of AD, the 3xTg line, decreases in both AVP and VIP were observed prior to the onset of tau pathology, but after the onset of Aβ pathology [33]. Thus, it was surprising that AD progression from 6–12 months had no effect on AVP or VIP expression in the AβPP/PS1 AD model. Future studies looking at neuropeptide expression in more advanced stages in this model could potentially replicate the loss of SCN neuropeptide expression seen in humans.
In the current study, exposure to chronic CD had no effect on AVP or VIP expression at the 6- or 12-month endpoints in female C57BL/6J or AβPP/PS1 animals. Chronic CD also had no effect on AVP or VIP expression in 6-month male C57BL/6J mice or AβPP/PS1 mice. However, both AVP and VIP were significantly reduced in 12-month AβPP/PS1 males exposed to CD. These data suggest that CD may affect SCN function, specifically in AβPP/PS1 males. A study in grass rats showed that chronic circadian desynchronization using a similar CD paradigm induced telomere shortening without changes to SIRT1 expression in liver cells, indicating cellular senescence [34]. It is possible that progressive AD pathology is acting as a second insult to chronic CD to significantly impact neuropeptide expression in the SCN of 12-month AβPP/PS1 males. Considering the important contribution of both AVP and VIP to photoentrainment, the diminished ability of AβPP/PS1 males to adjust to weekly changes in the lighting schedule may be attributed to reduced AVP and VIP expression in the SCN.
Glucose tolerance tests were administered on day 7 to determine whether chronic CD up to the 6- and 12-month endpoints had an impact on metabolic function. Shift workers have higher body mass indices and an increased prevalence of obesity; however, acute mouse models of shift work do not report significant changes in weight [35, 36]. Interestingly, these data indicate significant differences in fasting glucose levels between ND and CD males. At 6 months, CD c57BL/6J males have lower fasting glucose; at 12 months, both C57BL/6J and AβPP/PS1 males exposed to CD had lower fasting glucose levels compared to ND males. A decrease in fasting glucose levels, indicative of increased metabolic function, was counterintuitive and contradictory to our hypothesis that chronic CD would be detrimental to metabolic health. When put into context of the CD paradigm used in this study however, it is important to consider that metabolic function follows diurnal rhythms [37]. Fasting glucose measured in C57BL/6J mice demonstrate a clear diurnal rhythm with males showing a peak at the time of lights off, and females showing a peak 2 h earlier (Fig. 3A). Fasting glucose measurements were taken at 6:45 AM, which for ND animals was at approximately Zeitgeber time (or 3 h prior to lights off). For females, this approaches the time of peak fasting glucose. Because even CD females were aligned to the light/dark cycle on Day 7, it is likely that the circadian system and metabolism among the groups of females were also aligned and are reflected by similar levels of fasting glucose. For males, CD led to reduced fasting glucose, which may reflect the sex differences observed in adjustment to the lighting schedule.
At the end of this study, all animals were re-entrained to the same 12:12-h light/dark schedule for a period of 3–5 weeks prior to the MWM task and tissue collection. It was necessary to make sure that chronically disrupted animals were at the same point in their circadian cycle during tissue collection and MWM, since learning, memory, and soluble protein concentrations are circadian controlled [38, 39]. Thus, we wanted changes in learning and memory or protein expression to reflect a lasting effect of chronic CD as opposed to data collection at different circadian times. Corrected integrated path length (CIPL) was used as the primary parameter for assessing spatial learning through the 5-day training period, as it corrects for variables such as swim speed and the distance of the starting position from the platform location that could factor heavily into parameters such as trial duration. CIPL is a cumulative measure of the animal’s distance from the platform throughout the trial duration. Quickly locating the platform location or spending more time searching in close proximity to the platform location would thus result in lower CIPL measurements, indicating improved spatial learning across trial days.
While female AβPP/PS1 animals did present spatial learning deficits as compared to age- and sex-matched C57BL/6J controls, no effect of chronic CD was seen on spatial learning and memory. In contrast, male AβPP/PS1 animals exposed to chronic CD saw significant learning and memory deficits at 12 months of age as compared to animals on the ND schedule. Data from animals on the ND schedule showed that though insignificant, 12-month female AβPP/PS1 animals presented trending increases in amyloid plaque burden as compared to 12-month male AβPP/PS1 mice. Similarly, on day 1 and day 3 of the 5-day training period, 12-month females presented significantly higher CIPL and cumulative distance measurements than 12-month males, indicating diminished spatial learning. Both MWM and plaque data indicate that at least at the 12-month endpoint, female AβPP/PS1 mice have a worsened AD phenotype as compared to 12-month male AβPP/PS1 mice. Whereas ND AβPP/PS1 and C57BL/6J did not differ in MWM performance at 12 months, 12-month CD AβPP/PS1 males mice presented significantly higher CIPL and cumulative distance values on day 1 and 3 of the 5-day training period as compared to CD C57BL/6J 12-month males. Taken together, the data indicates that male AβPP/PS1 are more susceptible to the effects of CD in terms of ability to re-entrain and diminished neuropeptide expression in the SCN, and the current data indicates that this susceptibility could carry over to the consequential effects of chronic disruption on the AD phenotype, specifically spatial learning.
The findings from the current study indicate that while AD progression up to the 12-month endpoint in AβPP/PS1 mice or chronic CD following the mild protocol as outlined here are not singularly sufficient to produce changes in AVP or VIP neuropeptide expression in the SCN. Decreases in AVP and VIP expression in the SCN, however, appear to present as a sex-dependent, emergent property of both AD progression and chronic CD, displaying specifically in males. These effects also carried over into the cognitive phenotype, with 12-month AβPP/PS1 males presenting a significant impact of chronic CD on the MWM task. The increased susceptibility of males and the apparent resilience of females to the effects of chronic CD and the bidirectional impact that it has on AD progression are significant findings that serve as grounds for future investigations as to what potential mechanisms might be responsible for the resilience seen in females, as well as what biological mechanism might be interacting between the circadian system and AD pathological progression. This is particularly important, as women are disproportionately affected by AD. However, AD is a complex disease process. This study suggests that the mechanisms of disease may be sex-specific, with females protected from insults to the circadian system. Sex differences in exposure to CD in grass rats shows increased effects in the brain [40]. Finally, this mouse model does show seizure activity, especially at early stages, which could affect long-term results; repeating this experiment in other models of AD disease could aid in elucidating what aspects of AD pathogenesis might be contributing to these effects.
Footnotes
ACKNOWLEDGMENTS
The authors are grateful to Kathleen Bottum, MD/PhD for assistance with data analysis and a critical reading the manuscript.
FUNDING
Resources for this work were provided by the Dale and Deborah Smith Center for Alzheimer’s Research and Treatment, the Kenneth Stark Endowment, a team science grant from SIU School of Medicine, and NIH (R01AG057767 and AG061937).
CONFLICTS OF INTEREST
Erin R. Hascup is an Editorial Board Member of this journal but was not involved in the peer-review process nor had access to any information regarding its peer-review.
The remaining authors have no conflicts of interest to report.
DATA AVAILABILITY
The data supporting the findings of this study are available within the article and/or its supplementary material.