
Abstract
Background:
Neuroinflammation is one of the cardinal mechanisms of Alzheimer’s disease (AD). with amyloid-β (Aβ) playing a critical role by activating microglia to produce soluble inflammatory mediators, including several chemokines. Peripheral monocytes are, therefore, attracted into the central nervous system (CNS), where they change into blood-born microglia and participate in the attempt of removing toxic Aβ species. The translocator protein-18 kDa (TSPO) is a transmembrane protein overexpressed in response to neuroinflammation and known to regulate human monocyte chemotaxis.
Objective:
We aimed to evaluate the role of the oligomeric Aβ1-42 isoform at inducing peripheral monocyte chemotaxis, and the possible involvement of TSPO in this process.
Methods:
In vitro cell lines, and ex vivo monocytes from consecutive AD patients (n = 60), and comparable cognitively intact controls (n = 30) were used. Chemotaxis analyses were carried out through both μ-slide chambers and Boyden assays, using 125 pM oligomeric Aβ1-42 as chemoattractant. TSPO agonists and antagonists were tested (Ro5-4864, Emapunil, PK11195).
Results:
Oligomeric Aβ directly promoted chemotaxis in all our models. Interestingly, AD monocytes displayed a stronger response (about twofold) with respect to controls. Aβ-induced chemotaxis was prevented by the TSPO antagonist PK11195; the expression of the TSPO and of the C-C chemokine receptor type 2 (CCR2) was unchanged by drug exposure.
Conclusion:
Oligomeric Aβ1-42 is able to recruit peripheral monocytes, and we provide initial evidence sustaining a role for TSPO in modulating this process. This data may be of value for future therapeutic interventions aimed at modulating monocytes motility toward the CNS.
INTRODUCTION
Alzheimer’s disease (AD), widely recognized as the most common cause of dementia, is an age-related neurodegenerative disorder, characterized by progressive cognitive deterioration, affecting both memory and other aspects of cognitive functioning. Despite being well established that neurofibrillary tangles and amyloid-β (Aβ) plaques and oligomers [1] play a central role in the development of the disease, it is now recognized that neuroinflammation—the inflammatory response originated within the central nervous system (CNS)—contributes to AD pathogenesis just as much as its pathological hallmarks [2].
A crucial player of neuroinflammation is microglia, the immune cell of the brain and Aβ is known to be able to bind to microglia and activate these cells that in brain are tightly associated with senile plaques. Microglia then reacts by releasing inflammatory cytokines and chemokines [3]. Within the core of amyloid plaques, resident microglia, coexists with blood-derived macrophages, which are also able to develop a specific immune reaction against Aβ [4–6].
Curiously, the peripheral immune system has long been considered completely separated from the CNS by means of the blood-brain barrier (BBB); however, several studies have now reported evidence of the influence of aging [7], increased cytokines and chemokines concentration [8, 9], or the presence of noxious stimuli [10], in modulating BBB integrity and functions.
Indeed, peripheral blood monocytes enter the CNS through the BBB when attracted by chemokines, small chemotactic molecules strongly upregulated in the context of AD and specifically produced by activated microglia in plaque-rich sites [11–13]. The C-C chemokine receptor type 2 (CCR2) is known to play an important role in monocyte recruitment: in particular, CCL2 (also known as MCP-1), one of the most effective chemotactic soluble factors for monocytes, is upregulated near Aβ deposits as a result of its release from plaque-associated microglia [14–16].
Interestingly, the receptor systems modulating peripheral monocyte activation are also able to recognize protein aggregates, such as Aβ, which is known to belong to the group of “danger-associated molecular patterns”, i.e., potent inflammation perpetuators [17].
This led us to hypothesize that the presence of Aβ aggregates alone could be sufficient to influence the movement of peripheral immune cells into the brain, a process that we called ‘neuroinvasion’. This information could be of high interest in the era of passive anti-Aβ immunization, potentially defining a peripheral (and accessible) surrogate marker for disease progression and drug response. Indeed, data already available are showing that soluble Aβ may be a powerful chemotactic stimulus in both in vitro blood-brain barrier models, and AD transgenic mice [4, 18]. However, the characterization of Aβ-induced monocyte chemotaxis is still scanty and multiple signaling systems are postulated to modulate this process.
The translocator protein 18 kDa (TSPO), originally discovered as a peripheral binding site for benzodiazepines [19], is a transmembrane protein widely expressed throughout the body. Although TSPO is primarily located on the outer mitochondrial membrane, it has also been identified on the plasma membrane of various cell types, including lymphocytes and monocytes [20, 21]. In the CNS, under physiological conditions, TSPO expression is limited to glial cells and its levels are very low; however, they experience a dramatic increase in response to the rise of neuroinflammation [22]. Moreover, microglial cells strongly upregulate the expression of TSPO in their activated state in different neurodegenerative diseases, including AD, and this can be revealed in vivo through PET imaging with TSPO specific tracers, such as PK11195 [23, 24]. Peripheral monocytes’ recruitment through the BBB may give an additional contribution to the increase of TSPO levels in the CNS: indeed, previous ex vivo human studies demonstrated both the presence of functional TSPO receptors on monocytes’ surface [25] and the ability of various TSPO ligands to influence their chemotaxis. In particular, RO5-4864 has emerged as an agonist able to induce chemotaxis in human monocytes, while the process is blocked by the antagonist PK11195 [26, 27]. Of special interest, an alanine-to-threonine polymorphism (rs6971) in a region of TSPO that is highly conserved results in a conformational change affecting its interaction with a variety of ligands: the presence of the predominant form (alanine) is associated with high-affinity binding, while the polymorphism (threonine) shows reduced binding affinity [28, 29].
The potential role of TSPO in controlling Aβ-induced monocyte chemotaxis is still to be elucidated. Therefore, the present study aimed at investigating the ability of Aβ to promote peripheral monocytes chemotaxis, and the putative involvement of the TSPO receptor in modulating this event. We used both in vitro human cell lines (THP-1 monocyte leukemia cells and U-937 monocytes/pre-macrophage line), and ex vivo human monocytes obtained from AD patients and cognitively intact controls, challenging them with TSPO selective ligands characterized by different modulatory properties. Finally, in order to speculate on a possible correlation between TSPO chemotactic activity and its established rs6971 polymorphism, we included the evaluation of monocytes from AD patients characterized by different haplotypes.
METHODS
Subject recruitment
Following Ethical Committee approval (Prot. n. 20220500U - “Bando Progetti di rete - II Edizione” – Project CP2_16/2018 NEVERMIND) and informed consent, n = 60 consecutive AD outpatients were recruited at the memory services of the IRCCS “San Gerardo dei Tintori”, Monza (Italy) and “Manzoni” Hospital, Lecco (Italy). Dementia specialists diagnosed probable AD according to the NINCDS-ADRDA criteria [30] and alternative diagnoses were excluded by MR brain imaging and a routine extensive neuropsychological test battery. Patients were recruited only if the main caregiver was available for the interview. For each patient, current scores at the Mini-Mental State Examination (MMSE) was obtained, and the eventual use of the acetylcholinesterase inhibitors was noted. Thirty age- and sex-comparable healthy controls (CTRL) were additionally recruited among patients’ spouses, without personal or family history of neurological or psychiatric disorders; lack of cognitive impairment was established by a clinical interview, including a MMSE score > 26. Either patients or controls, with recent infections or surgery (6 months) or under anti-inflammatory, corticosteroid, or immunosuppressive drug treatments, or affected by kidney or liver failure were excluded. Clinical and demographic data are shown in Table 1. The full population was used for genotyping and expression studies, while chemotaxis studies were performed in a subgroup of n = 8 AD patients (M/F 5/3, age 80.5±4.2 years, MMSE score 18.0±5.6) and n = 8 sex- and age-matched (±3 years) cognitively spared controls.
Demographic and clinical characteristic of the recruited subjects. AD, Alzheimer’s disease; CTRL, cognitively intact controls; where appropriate, data are shown as mean
Monocyte sample preparation and APOE genotyping
Blood samples (15 ml) were collected in dipotassium ethylene-diamine-tetra-acetic acid (4.08 mM final concentration) from the antecubital vein after over-night fasting, always between 08.00 and 09.00 A.M., to obtain peripheral blood mononuclear cells (PBMC), isolating them by density-gradient centrifugation by Lympholyte (Cedarlane, Burlington, Canada) [31]). Briefly, whole blood samples were diluted with the same amount of saline solution layered on Lympholyte and centrifuged (490 g, 30 min, room temperature). PBMCs were collected from the interface between plasma and Lympholyte, washed with saline solution, aliquoted and stored at –80°C for further protein or RNA extraction. For monocyte isolation PBMC were immediately transferred to a six well plates (see below).
To analyze the apolipoprotein E (APOE) genotype, total DNA was extracted from peripheral blood using a commercial DNA extraction kit (Qiagen, Venlo, Netherlands), and DNA amplification was performed using specific primers, as previously reported [32].
Cell cultures
THP-1 (derived from an acute monocytic leukemia, ATCC, USA) and U-937 (derived from a histiocytic lymphoma, ATCC) cell lines were used. The culture conditions were: RPMI 1640 plus 10% FBS, 1% penicillin streptomycin, 37°C, 5% CO2.
Cells were exposed with different compounds: oligomeric Aβ42 125 pM, Ro5-4864 10/25/50μM (Merck KGaA, Darmstadt, Germany), Emapunil 1/50μM (Merck), and PK11195 20/120μM (Merck) in combination or not with oligomeric Aβ42 125 pM. Oligomerization of Aβ42 was performed according to Stine and colleagues [33]. Briefly, 200μg of lyophilized Aβ42 (Phoenix Pharmaceuticals, Inc., USA) was dissolved in 200μl of 1,1,1,3,3,3-Hexafluoroisopropanol (HFIP, Merck), divided in 10μl (corresponding to 10μg of Aβ1-42) aliquots and dried. To obtain Aβ42 oligomers, a 10μg aliquot was first resuspended in 1μl of DMSO (Merck), then 43μl of cold PBS was added and the vial was sonicated for 10 min on ice and left at 4°C overnight. After the incubation, the vial was centrifuged (14,000 g, 10 min, 4°C) and the supernatant containing Aβ42 oligomers was stored at –20°C. Immunocomplexes of oligomeric Aβ42 and anti-Aβ42 monoclonal antibodies (clone G211, Merck) were prepared according to Xiang and colleagues [34] by incubating oligomeric 125 pM Aβ1-42 with anti-Aβ42 monoclonal antibodies (25 ng/ml) for 1 h at 37°C.
Chemotaxis of THP-1 by μ-slide chambers
THP-1 cells were suspended in gel matrix (CorningTM MatrigelTM) to reach the final concentration of 3×106 cells/ml, then 6μl of this suspension were loaded in the observation area of the μ-slide chamber, each slide has three different chambers for parallel assays (Ibidi®, Gräfelfing, Germany) and set aside until the gel was solid. Later, one lateral chamber was filled with cellular medium alone and the other one with cellular medium and 15μl of chemoattractant (MCP-1 10 ng/ml or oligomeric Aβ42 125 pM). Cellular migration was recorded by time-lapse microscopy and the migratory trajectories of 20 cells/chamber were designed using the Chemotaxis and Migration Tool (Ibidi®). The Forward Migration Index (FMI II, parallel component of the chemotactic gradient) was calculated to measure cellular motility: higher migration rates correspond to higher FMI II values. Results were obtained from at least n = 5 separated experiments, as reported in each Figure legend.
Chemotaxis of THP-1 or U-937 and human monocytes by Boyden chambers
For these experiments, Boyden chambers (CorningTM TranswellTM) were used. For monocyte isolation, plastic adherence method was used. Briefly, 6×106 PBMC per well were seeded into 6 wells plates and allowed to adhere in a 5% CO2 incubator at 37°C for 2 h in 3 ml of culture medium. Non-adherent cells were removed and adherent cells were carefully washed twice with PBS, than adherent monocytes were collected and suspended in cellular medium with serum at 2% to reach the final concentration of 5×105 cells/ml. 600μl of each chemoattractant solution containing Aβ1-42 in combination or not with TSPO ligands, were loaded in the lower chamber of each well and an insert was added to separate this part from the upper one by means of a filter (pores Ø: 5μm), 100μl of monocyte suspension (5×104 cells) were loaded in the upper chamber. Analogously, 100μl of the cellular suspension of THP-1 or U-937 cells at a concentration of 8×105 in complete medium with serum at 2% were loaded in the insert with pore diameter of 8μm. For all cellular type the plates were incubated at 37°C for 3 h. After the incubation, the filters were fixed, colored with a modified-Giemsa staining, removed from the inserts, mounted on glass slides, and observed at the optical microscope (magnification 10x). Nucleated cells in 20 optical fields/filter were counted and their number was used as a chemotactic index, considering the ratio between the number of cells migrated toward the stimulus and the number of cells in absence of any stimuli. Results were obtained from at least n = 5 separated experiments, as reported in each figure legend.
mRNA expression by real-time polymerase chain reaction
mRNA expression levels were quantified on stimulated THP-1 and on PBMC obtained from 60 AD patients and 30 CTRL. Total RNA was extracted from THP-1 or PBMC using the RNeasy Mini kit (Qiagen), according to the manufacturer instructions. cDNA was synthesized from 500 ng of RNA using the SuperScriptVILO cDNA Synthesis Kit (Invitrogen, by Thermo Fisher Scientific, Waltham, MA, US) at the following conditions: 10 min at 25°C and 60 min at 42°C. The reaction was terminated at 85°C for 5 min and tubes chilled on ice. cDNAs were stored at –20°C until quantitative polymerase chain reaction (qPCR) assay. Primers sequences were: TaqMan probes (Applied Biosystems, Thermo Fisher Scientific) for TSPO (Hs00559362_m1) and β-actin (Hs01060665_g1) and KiCqStart™ Primers pre designed by Sigma Aldrich (Merck) for CCR2 and β-actin. For the quantification of TSPO mRNA TaqMan Gene expression kit (Thermo Scientific) was used under these conditions 1 cycle at 50°C for 2 min, 1 cycle at 95°C for 10 min, 40 cycles at 95°C for 15 s, 60°C for 20 s. For the quantification of the other targets cDNA was amplified in triplicate using the Titan HotTaq EvaGreen™ qPCR Mix (ROX) (Bioatlas, Tartu, Estonia), at the following conditions: 1 cycle at 50°C for 2 min, 1 cycle at 95°C for 15 min, 40 cycles at 95°C for 15 s, 58 °C for 20 s. Samples were assayed in a relative qPCR using the ABI Prism 7500 HTSequence Detection System (Applied Biosystems) for the analysis of mRNA levels of protein of interest as compared with mRNA encoding beta actin, as internal standard. For relative quantification of mRNA of interest versus housekeeping gene mRNA the comparative CT (cycle threshold) method was used.
Protein expression by western blotting
Cell pellets were lysed in cell extraction buffer (Invitrogen) supplemented with 1 mM PMSF and protease inhibitor cocktail (Merck) and protein concentrations determined by Bradford’s method. Following denaturation, 40μg proteins were separated by electrophoresis in 4–12% SDS-PAGE and transferred to nitrocellulose. Blots were blocked for 1 h, incubated overnight at 4°C with specific primary antibodies: TSPO (ThermoFisher Scientific) 1:500 dilution, or CCR2 (Cell Signaling Technology) 1:1,000 and then with the HRP-linked anti-goat or anti-rabbit IgG for 1 h. Beta-tubulin (Merck, 1:30,000 dilution) was used as internal standard. Signals were revealed by chemiluminescence, visualized on ImageQuant™ LAS 4000 and quantified by Image J software.
TSPO rs6971 genotyping
Genomic DNA was extracted from venous blood samples using Qiagen QIAamp DNA Blood Midi Kit (Qiagen). Primers amplifying TSPO sequence containing rs6971 SNP were designed using Primer3.0 (http://bioinfo.ut.ee/primer3-0.4.0) Web-based server and their specificity was checked by Primer blast on NCBI genome browser (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). PCRs were performed directly on 50-100 ng of genomic DNA in standard conditions using the forward 5’-CCTTGGTGGATCTCCTGCTGGT-3’ and reverse 5’-ACATCACAAGCGTGATGGCACC-3’ primers. Sanger sequencing was carried out directly on both strands of purified PCR products by using the BigDye Terminator Cycle Sequencing kit v3.1 and an automated ABI-3130 DNA sequencer (Applied Biosystems). ChromasPro v1.34 (Technelysium Pty Ltd.) software was used for mutation detection.
Statistical analysis
Data are shown as mean±standard deviation (SD). Statistical analysis was performed with GraphPad Prism, version 7.00. Two-tailed Student’s t test was used for computing differences between two groups, while analysis of variance (ANOVA) was used for three or more groups, followed by Tukey multiple comparison post hoc test.
RESULTS
Oligomeric Aβ42 induces chemotaxis both in THP-1 or U-937 cells and in human monocytes
Oligomeric Aβ42 125 pM was able to attract THP-1 cells when initially analyzed by μ-slide chambers. Under oligomeric Aβ42 induction, the direction of cell motility, calculated as forward migration index (FMI II), resulted two-fold increased with respect to vehicle. These data were in line with the results obtained after MCP-1 stimulation (MCP-1 10 ng/ml FMI II 0.35±0.04 and Aβ 125 pM 0.36±0.02 versus vehicle 0.18±0.04, (p < 0.01), see Fig. 1A, B).
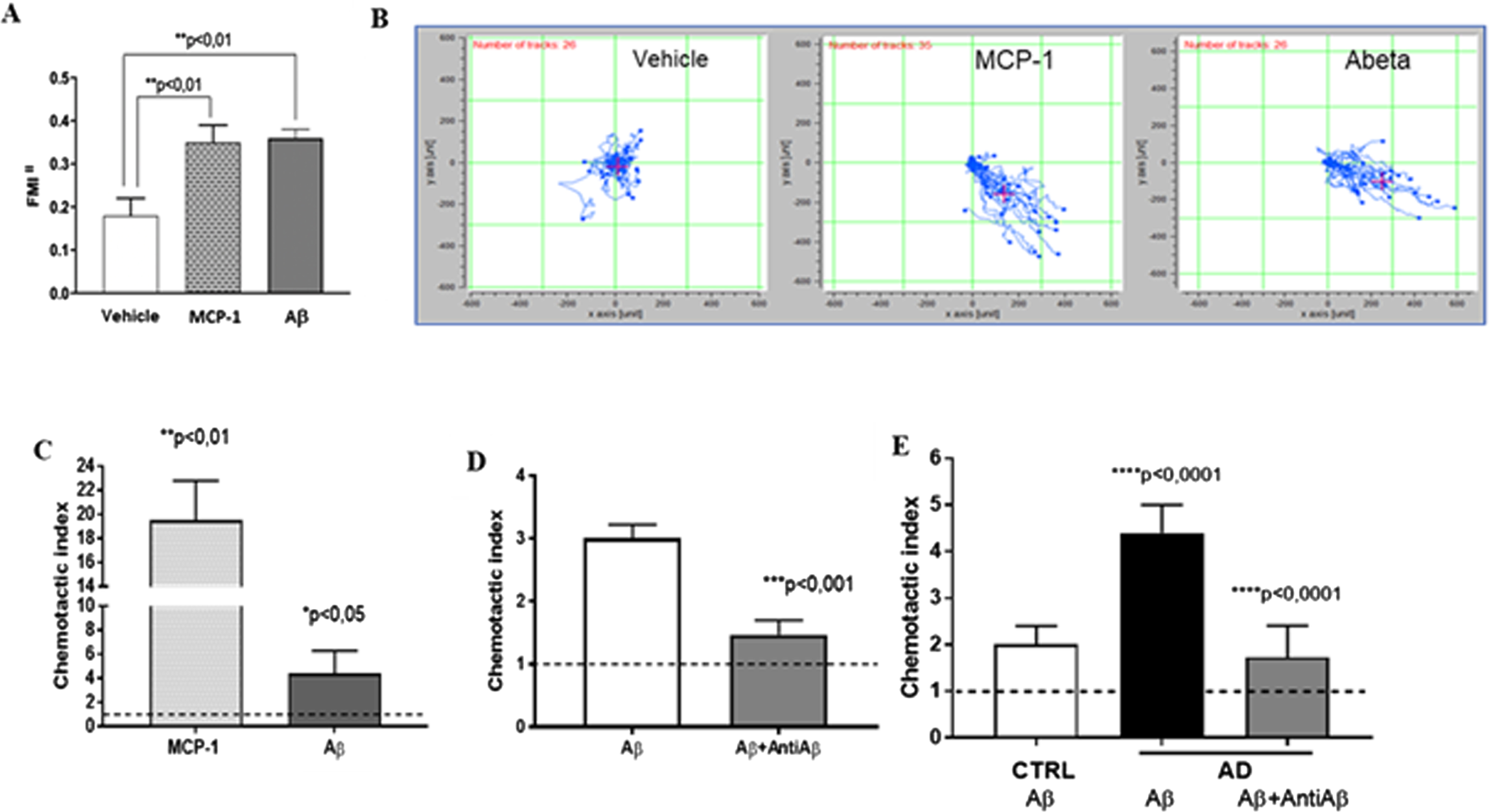
Oligomeric Aβ42 125 pM is an efficient monocyte chemotactic stimulus. A) Chemotaxis of THP-1 cells measured by μ-slide chambers. p < 0.01 at ANOVA, followed by Tukey’s multiple comparisons test, p < 0.01 versus Vehicle, n = 5. B) Typical trajectories toward chemoattractant solutions of THP-1 cells in μ-slide chambers are represented. C) Chemotaxis of monocytes from control subjects measured by Boyden chambers. p < 0.01 at ANOVA, followed by Tukey’s multiple comparisons test, p < 0.01 MCP-1 versus Vehicle, p < 0.05 Aβ42 versus Vehicle, n = 6. D) Chemotaxis of U-937 cells measured by Boyden chambers. p < 0.0005 at ANOVA, followed by Tukey’s multiple comparisons test, p < 0.001 Aβ + AntiAβ versus Aβ (n = 5). E) Chemotaxis of monocytes from AD patients with respect to heathy matched Controls measured by Boyden chambers. p < 0.01 at ANOVA, followed by Tukey’s multiple comparisons test, p < 0.0001 AD Aβ versus CTRL, p < 0.0001 AD Aβ+AntiAβ versus AD Aβ, n = 8).
Similar results were obtained using Boyden chambers and other cell types (U937 and ex-vivo human monocytes). In fact, monocytes from selected healthy controls (n = 6) stimulated with oligomeric Aβ42 125 pM in Boyden chambers showed a significant increase in chemotaxis with respect to vehicle (horizontal line), albeit, in this case, of lesser magnitude with respect to MCP-1 10 ng/ml stimulation (Aβ42 125 pM: 4.4±1.9, p < 0.05; MCP-1:19.5±3.3; p < 0.01, Fig. 1 C).
We confirmed the chemotactic properties of oligomeric Aβ42 by demonstrating that the sequestration of Aβ by anti-Aβ42 monoclonal antibodies (clone G211, 25 ng/ml) significantly halved Aβ-induced cell migration both in U937 and in monocytes from AD patients (U937: Aβ42 125 pM: 3.01±0.21 Aβ+AntiAβ: 1.47±0.23, p < 0.001 versus Aβ; AD patient monocytes: Aβ42 125 pM 4.4±0.6 versus Aβ+AntiAβ: 1.73±0.68, p < 0.0001, n = 8; Fig. 1D, E).
We finally evaluated the chemotactic properties of monocytes from selected AD patients with respect to matched elderly CTRL subjects. Monocytes from AD patients showed a more than two-fold increment in Aβ42-induced chemotaxis with respect to cells from elderly controls (AD 4.4±0.6 versus CTRL 2.0±0.4, p < 0.0001, n = 8; Fig. 1E).
Monocyte chemotaxis is mediated by TSPO
TSPO agonists were tested in chemotaxis assays on monocytes from control subjects and AD patients. The selective TSPO agonist, Emapunil 1μM alone induced an increase of 75% of chemotaxis in monocytes from AD patients (p < 0.05). Analogously, the TSPO agonist Ro5-4864 10μM was able to increase chemotaxis in monocytes obtained from both, AD patients (76%, p < 0.05), and comparable controls (about two-fold, p < 0.01; see Fig. 2A).
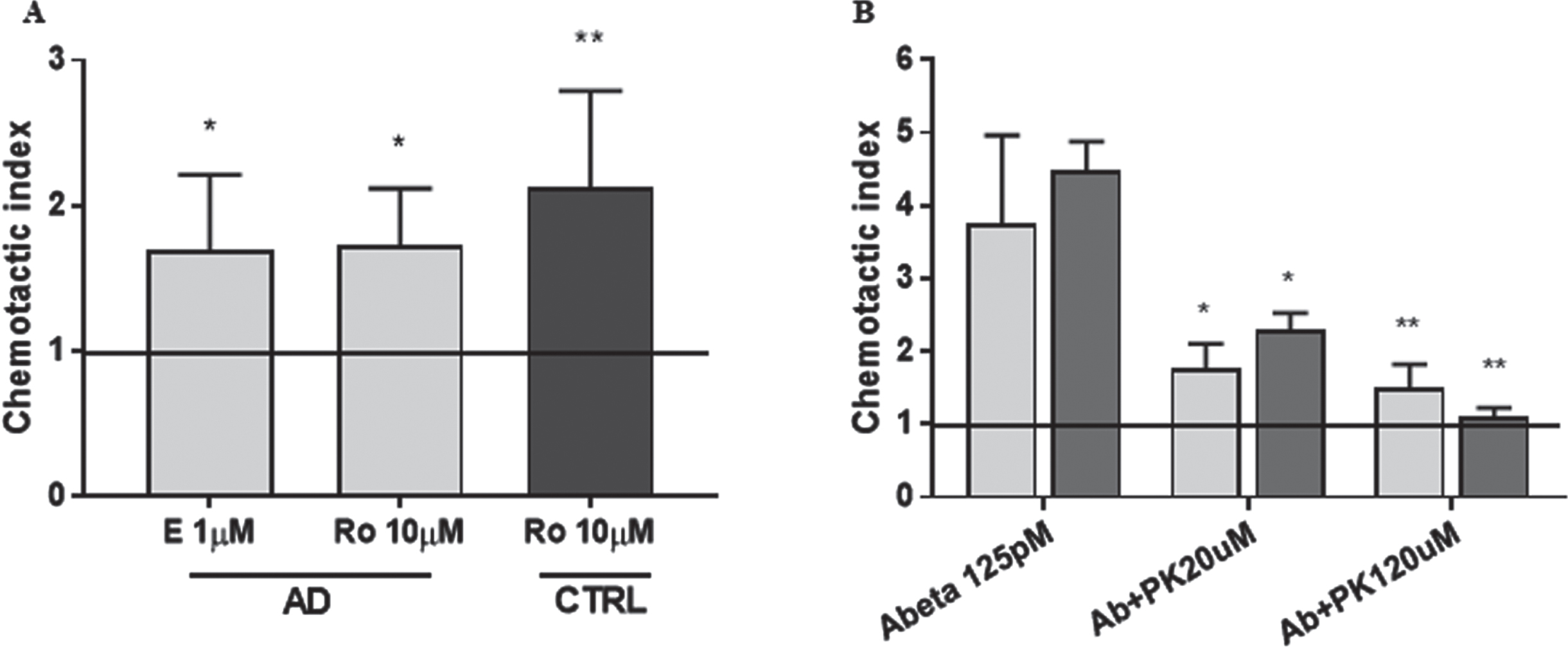
TSPO is a relevant modulator of monocyte chemotaxis. A) TSPO agonists Emapunil (E 1μM) and Ro5-4864 (Ro 10μM) induced chemotaxis in monocytes obtained from both AD patients and healthy controls (CTRL). p < 0.01 at ANOVA followed by Tukey’s multiple comparisons test, *p < 0.05 and **p < 0.01 versus Vehicle (horizontal line), n = 5; B) TSPO antagonist PK11195 prevents oligomeric Aβ42-induced chemotaxis in both, THP-1 cells (light grey bars, n = 6 replicates), and monocytes from AD patients (dark grey bars, n = 5). p < 0.05 at ANOVA, followed by Tukey’s multiple comparisons test, *p < 0.05 and **p < 0.001 versus Aβ42.
The TSPO inhibitor PK11195 was then tested in Boyden chambers assays. PK11195 prevented oligomeric Aβ42-induced chemotaxis in a significant way both, in THP-1 cells (∼60% decrease, p < 0.05 Aβ + PK11195 20μM and p < 0.001Aβ + PK11195 120μM versus Aβ, Fig. 2B, light grey bars), and in monocytes from AD patients where the effect was concentration-dependent (40% and 80% decrease for PK11195 20μM, p < 0.05 and 120μM p < 0.001, respectively; Fig. 2B, dark grey bars). The administration of PK11195 alone at both concentrations did not affect the chemotactic behavior (data not shown).
Considering the potential role of the TSPO rs6971 polymorphism in modulating PK11195 binding, we tested also for the presence of a possible correlation with the chemotactic activity. We initially characterized the whole cohort of AD patients (n = 60), showing a distribution (GG 51%, GA 35%, AA 14%) not dissimilar to what was previously reported [29]. Subsequently, oligomeric Aβ42-induced chemotaxis was evaluated by Boyden assay in monocytes from AD patients divided according to the TSPO polymorphism. No changes were shown between the different tested haplotypes (n = 5 for each haplotype, data not shown). Also the APOE ɛ4 allele was represented in the AD population according to literature expectation [35], and we did not find any change comparing APOE ɛ4 carriers with respect to non-carriers (n = 4, data not shown).
TSPO and CCR2 expression is not modified by Aβ42 exposure and is unaltered in AD monocytes
TSPO and CCR2 mRNA and the protein expression levels were quantified following oligomeric Aβ42 treatment (125 pM/24 h). In this condition, Aβ42 failed to modify in THP-1 cells the expression levels of target mRNAs (Aβ42: 105% and 104% of vehicle, for TSPO and CCR2, respectively, not significant, n = 5) and proteins (Aβ42: 104% and 98% of vehicle, for TSPO and CCR2, respectively, not significant, n = 5). Moreover, TSPO mRNA and protein levels were assessed following exposure to Emapunil, Ro5-4864, and PK11195, alone or in combination with Aβ42, without showing any change (data not shown).
Finally, both target mRNAs were ex vivo assessed in AD patients and compared to comparable controls, again without finding any difference (TSPO mRNA: 1.45±1.23 versus 1.54±1.03, respectively n = 60 AD versus n = 30 CTRL; CCR2 mRNA: 1.08±0.78 versus 1.13±0.55, respectively n = 20 AD versus n = 10 CTRL; not significant) (Fig. 3). Finally, mRNA data failed to segregate with clinical and demographic data available.
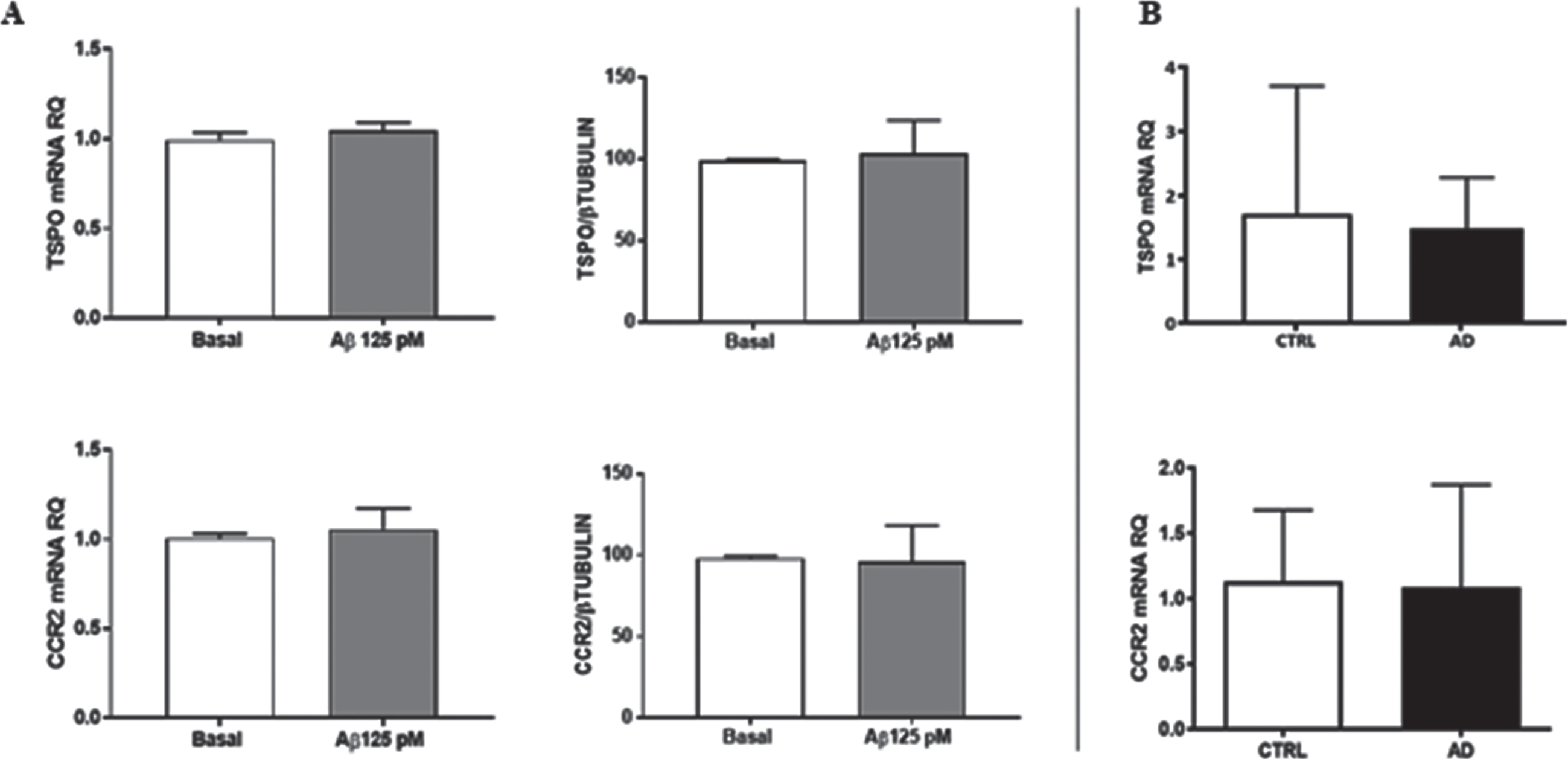
TSPO and CCR2 expression levels. A) mRNA and protein levels in THP-1 cells exposed to oligomeric Aβ42 125 pM; n = 5. B) TSPO and CCR2 mRNA levels in lymphomonocytes obtained from AD patients with respect to matched controls; (n = 20 AD versus n = 10 CTRL for CCR2 mRNA; n = 60 AD versus n = 30 CTRL for TSPO mRNA).
DISCUSSION
The aim of the present study was to evaluate the ability of Aβ, the key protein accumulated in the AD brain, to promote the peripheral monocytes chemotaxis, and to assess the putative involvement of the TSPO receptor in this circumstance.
Despite the peripheral immune system being long considered completely separated from the CNS, it is now widely accepted that, in AD, blood-derived monocytes are recruited in the brain through a dysfunctional BBB and they settle within the core of the amyloid plaques, shifting to blood-borne macrophages [4]. Furthermore, hyperactivation of peripheral monocytes plays a pivotal role during the progression from the prodromal stages to full-blown AD dementia [12].
We hypothesized that in AD brains the presence of increasingly complex aggregated states of Aβ, could be enough by themselves to influence the movement of peripheral immune cells into the brain, a process that we have named ‘neuroinvasion’. What we showed through time-lapse microscopy was a clear tendency of THP-1 cells to migrate towards oligomeric Aβ42 to the same extent as when they are exposed to MCP-1, the most effective chemotactic factor for monocytes. Boyden chambers experiments confirmed these results also on human monocytes from elderly controls, showing an increased migration rate following exposure to Aβ, even though the effect was of reduced magnitude with respect to the classical chemoattractant. In any case, it should be pointed out that Aβ42 was efficient in inducing monocyte chemotaxis at a concentration of the same order of magnitude as the one detected in human CSF (125 pM). Finally, our experiments suggested that an alteration of peripheral immune cell migration is operative in AD, since monocytes obtained from AD patients resulted significantly more prone to Aβ-induced chemotaxis compared to those from healthy controls. We may hypothesize that cells from AD patients could have already been primed by the chronically exposure to Aβ, or that more vigorous inflammatory responses may characterize the inflammaging milieux predisposing to the disease [36, 37]. These results are in line with the data obtained by Zhang and colleagues [38], describing an increased ability of monocytes isolated from AD patients to cross a model of endothelial barrier upon Aβ stimulation.
Our subsequent investigations focused on the putative role of TSPO, a transmembrane receptor previously recognized to be involved in monocyte chemotactic responses [26], by testing it in the specific context of AD pathology.
We initially confirmed the influence of different selective ligands, known as chemotactic modulators for peripheral monocytes [26, 27]: the TSPO agonists Emapunil and Ro5-4864 were both able to induce an increase of chemotactic properties in either monocyte-related cell lines and human monocytes from control subjects and AD patients. On the other hand, TSPO selective inhibitor PK11195 alone did not alter monocyte chemotactic behavior respect to baseline, but rather partially reverted Aβ-induced chemotaxis in a concentration dependent manner, perhaps suggesting that oligomeric Aβ42 and PK11195 could be competing for the very same binding sites. Taken together, these findings point towards a clear involvement of TSPO in the chemotaxis of peripheral monocytes in the context of AD, where the TSPO is classically upregulated [39].
Our first hypothesis was that differences in monocyte chemotactic behavior could be dependent on the modulation of TSPO expression levels; however, neither exposure to Aβ, nor to TSPO selective ligands were able to affect the TSPO mRNA or the protein abundance. Surely, the time of exposure is a variable that deserves further attention, since in vivo exposures to Aβ are very long lasting, and we cannot exclude a potential modulatory effect when changing the experimental conditions. Furthermore, considering these findings, additional investigations would be required to determine if also TSPO cellular distribution, rather than its expression, may be affected by Aβ exposure. As a matter of fact, the biological function of the receptor is intimately linked to its membrane localization, and an interdependence between chemotactic activity and TSPO targeting to the plasma membrane of the monocytes might exist. According to this premise, LPS was shown to increase in TSPO cell surface expression in human monocytes [40, 41].
On the other hand, we also explored a possible correlation between monocyte chemotactic modulation and the rs6971 TSPO polymorphism. As a matter of fact, this alanine-to-threonine polymorphism results in a conformational change, potentially affecting TSPO binding affinity for its selective ligands [28]; the general hypothesis was that a reduced affinity binding could be associated with a decreased migration rate. Contrarily to this expectation, the evaluation of Aβ-induced chemotaxis in monocytes from AD patients characterized by different haplotypes failed to show any meaningful difference.
The TSPO system, in any case, is not alone in controlling monocyte chemotaxis and the CCR2/CCL2 signaling is well known to play a major role in the recruitment and accumulation of immune cells at the level of senile plaques [14, 15]. CCR2, the surface receptor for MCP-1 (also known as CCL2), is mainly expressed on the surface of monocytes [42], and recent evidence supports the hypothesis that a downregulation in monocytes from AD patients [2, 16] can contribute to Aβ accumulation and cognitive decline [43]. MCP-1 expression is linked to Aβ pathology, as Aβ-induced upregulation of MCP-1 has been demonstrated in microglia, astrocytes, and human monocytes [44, 45], and we considered the possibility that the same mechanism could contribute to CCR2 modulation. However, no difference was observed in mRNA and protein levels of CCR2 in monocyte-related cell lines and monocytes from AD patients upon exposure to Aβ.
In conclusion, we confirm a robust monocyte chemotactic role for Aβ and the involvement of TSPO, no longer a simple marker of neuroinflammation but a real full player. The major question, however, i.e., whether or not blocking neuroinvasion could be beneficial or harmful for neuronal survival in AD, remains yet to be fully elucidated and we may merely postulate that future therapeutic agents able to counteract neuroinflammation may be focusing on Aβ-induced monocyte chemotaxis, by modulating it directly in the peripheral compartment [46].
Footnotes
ACKNOWLEDGMENTS
We thank Dr. Luca Mologni (University of Milano-Bioccoca, Italy) and Dr. Erica Dander (Tettamanti center, Fondazione IRCCS San Gerardo dei Tintori, Monza, Italy) for their valuable help.
FUNDING
This work was supported by Grant # 2020-ATE-0018 UNIMIB (LT).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The data supporting the findings of this study are available on request from the corresponding author.