
Abstract
Background:
The longevity gene Klotho (KL) was recently associated with neurodegenerative diseases including Alzheimer’s disease (AD). Its role in the brain has not been completely elucidated, although evidence suggests that KL-VS heterozygosity is associated with a reduced risk of AD in Apolipoprotein E ɛ4 carriers. Conversely, no data about genetic association with frontotemporal dementia (FTD) are available so far.
Objective:
To investigate the involvement of KL in AD and FTD by the determination of the genetic frequency of KL-VS variant and the expression analysis of KL gene.
Methods:
A population consisting of 438 patients and 240 age-matched controls was enrolled for the study. KL-VS and APOE genotypes were assessed by allelic discrimination through a QuantStudio 12K system. KL gene expression analysis was performed in a restricted cohort of patients consisting of 43 AD patients, 41 FTD patients and 19 controls. KL gene expression was assessed in peripheral blood mononuclear cells with specific TaqMan assay. Statistical analysis was performed using GraphPad 9 Prims software.
Results:
KL-VS frequency was comparable to the ones found in literature and no differences were found in both allelic and genotypic frequencies between patients and controls were found. Conversely, KL expression levels were significantly lower in AD and FTD patients compared with controls (mean fold regulation – 4.286 and – 6.561 versus controls in AD and FTD, respectively, p = 0.0037).
Conclusion:
This is the first study investigating KL in FTD. We showed a decreased expression of the gene in AD and FTD, independent of the genotype, suggesting a role of Klotho in common steps during neurodegeneration.
INTRODUCTION
α-klotho gene, hereafter referred to as klotho (KL), is a longevity factor, serendipitously discovered in 1997, when its accidental disruption resulted in a phenotype resembling aging in a mouse model [1]. KL gene is located on Chromosome 13 and encodes for a single-pass transmembrane protein, mainly expressed in kidney and brain. Klotho is known to be involved in phosphate and calcium homeostasis [2, 3], but its role in the brain has not been fully elucidated. KL deficient mice displayed cognitive decline when learning and memory were tested [4], while overexpression of klotho enhances such performances [5]. Although the klotho deficient mouse does not recapitulate a clear neurodegenerative disease, several studies have been addressed at finding a link between Klotho and the cognitive decline typical of neurodegeneration in humans. Higher plasma Klotho levels have been found to be associated to lower risk of cognitive decline [6]. In older humans, cerebrospinal fluid (CSF) Klotho appeared to be decreased, and even lower levels were found in patients with Alzheimer’s disease (AD) [7]. An intriguing link has been proposed with the frontotemporal dementia (FTD) spectrum as well, since Ho and colleagues found deregulation of Klotho levels in Chromosome 9 Open Reading Frame 72 gene (C9ORF72) knockout mice [8]. Moreover, functional genetic variants of KL have been characterized, finding further association with brain health. KL with Valine and Serine substitutions (KL-VS) is the most common KL variant. It encompasses six single nucleotide polymorphisms (SNPs) in perfect linkage disequilibrium [9]. Three SNPs are intronic and do not affect splicing; one is silent; two are missense substitutions leading to amino acid changes which give the name to the variant. KL-VS variant in heterozygous status is associated to healthy aging and appeared to mitigate amyloid and tau burden in AD [10, 11]. Interestingly, KL-VS homozygosity is associated with an opposite phenotype: reduced life expectancy and cognitive decline [12, 13]. Moreover, Belloy and colleagues demonstrated that KL-VS genotype was associated with reduced AD risk in Apolipoprotein (APO)E ɛ4 carriers [14]. Recent findings showed that Klotho protein levels in CSF and plasma correlated with KL-VS heterozygosity status and AD CSF biomarker levels [15].
To further investigate the involvement of KL in neurodegenerative diseases, we assessed the frequency of KL-VS variant and KL expression levels in AD and FTD patients. Healthy geriatric subjects were analyzed as the control group.
METHODS
Population
In order to assess the frequency of KL-VS genotype, 678 subjects were retrospectively recruited for this study. 438 patients were referred to Alzheimer Unit of the IRCCS Fondazione Ca’ Granda, Ospedale Maggiore Policlinico, University of Milan (Milan, Italy) in suspicion of dementia. They underwent the standard clinical workup, comprising detailed medical history, physical and neurological examination, screening laboratory tests, Mini-Mental State Examination (MMSE); qualitative brain magnetic resonance imaging (MRI) or computed tomography (CT). For a better characterization, 225 patients underwent also fluorodeoxiglucose positron emission computed tomography (FDG-PET). The presence of significant vascular brain damage was excluded (Hachinski Ischemic Score < 4). All patients underwent lumbar puncture for the analysis of CSF biomarkers Aβ, total tau (tau), and tau phosphorylated at position 181 (Ptau). The diagnosis of AD (n = 221) or FTD spectrum (n = 217) was made in accordance with current research criteria [16, 17].
A cohort of 240 healthy geriatric subjects, provided by the Geriatric Unit of the same Institution, was enrolled as a control group.
Controls were tested for frailty, according to Canevelli’s Frailty Index (FI), which defines frailty as a product of a cumulative deficits model that could be considered as an objective marker of “biological aging” [18].
Demographic and clinical data are reported in Table 1.
Demographic information of total cohort
Demographic information of total cohort
AD, Alzheimer’s disease, FTD, frontotemporal dementia; SD, standard deviation; y, years; n.p., not performed; n.a., not applicable; n.s., not significant.
Informed consent was obtained from all participants or their caregivers, and the study was approved by the local Institutional Review Board (Parere 532_2019bis del 13-6-2019-Comitato Etico Milano Area 2).
Cerebrospinal fluid analysis
CSF samples were collected into 15 mL polypropylene tubes by lumbar puncture in the L3/L4 or L4/L5 interspace. Samples were centrifuged at 2000 r/min for 10 min at 4°C. The supernatants were stored at – 80°C until use. CSF Aβ, tau, and Ptau were measured with ELISA kits (Fujirebio, Ghent, Belgium). Normal values of biomarkers were: Aβ>600 pg/mL; tau < 400 pg/mL and Ptau < 61 pg/mL [19].
Allelic discrimination
Allelic discrimination assay has been performed in patients and controls in order to assess the frequency of the three KL-VS genotypes (wild-type, heterozygote, and homozygote). Genomic DNA was extracted from whole blood. Using a TaqMan allelic discrimination assay (Thermo Fisher Scientific), performed on QuantStudio 12K Real-time system (Applied Biosystem), two tagging SNPs identifying the variant were screened, namely rs9536314 (T/G) for F352 V and rs9527025 (G/C) for C370 S. Patients were also characterized for APOE genotype (rs429358 and rs7412).
Gene expression study
The gene expression study was performed in a subgroup of patients for whom whole blood was available for RNA extraction. Total RNA was extracted from whole blood collected in PAXgene tubes, containing reagents that lyse blood cells and stabilize RNA for downstream analysis. RNA was purified using PAXgene blood RNA kit (PreAnalytix) following manufacturer’s instructions. RNA was analyzed with 2100 Bioanalyzer and RNA 6000 Nano kit (Agilent Genomics) for qualitative and quantitative characterization.
Retrotranscription of total RNA was performed with SuperScriptTM ViloTM cDNA Synthesis Kit (Thermo Fisher Scientific) according to manufacturer’s protocol.
For gene expression analysis, specific TaqMan probes were used for KL gene, and for β-Actin (ACTB) and Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as house-keeping genes. Real Time PCR was run on QuantStudio 12K Flex system (Thermo Fisher Scientific).
Statistical analysis
Allelic and genotypic frequencies were obtained by direct counting. Chi Square test was used to test differences in allele distributions between the groups. Odds ratio (OR) was calculated along with its 95% confidence interval (CI).
For KL gene expression analysis, 2-ΔΔCt method was used to calculate the relative quantification (RQ) [20]. RQ was then converted into fold-regulation [21].
Normality test was performed to assess data distribution; accordingly, appropriate tests (t-test or ANOVA and relative non parametric tests) have been applied to compare gene expression levels among the three groups, and to correlate them with clinical data (Spearman test).
RESULTS
Genotype distribution study
Allele and genotype frequencies of KL-VS variant are shown in Table 2 and are comparable to the ones found in literature [5, 12]. For the statistical analysis, taking into account low frequency and deleterious effect of KL-VS homozygosity [12, 13], we grouped subjects into KL-VS carriers (KL-VShet +) and non-carriers (KL-VShet–). No significant differences were found in both allelic and genotypic frequencies between patients and controls as well (p > 0.05).
KL-VS genotype frequency
Statistical analysis was performed using GraphPad 9 Prism (San Diego, CA). Significant threshold was set at p < 0.05.
Allele and genotype frequencies expressed as n (%) of KL rs9536314 (T/G) and rs9527025 (G/C) SNP in patients and controls. p > 0.05. n.s., not significant
We stratified for sex and age at onset (early, i.e.,<65 years versus late), finding no significant differences in KL-VShet + versus KL-VShet– group, both for AD and FTD patients.
Moreover, for each group, we stratified KL-VS status for the presence of APOE ɛ4 allele. In AD group, we found a comparable distribution among the KL-VShet– group, while KL-VShet + group showed a higher percentage of APOE ɛ4 allele carriers [p = 0.0405; OR(95% CI)=2(1.05– 3.83)] (Table 3).
KL-VShet– and KL-VShet + groups stratified for APOE status
Genotype distribution expressed as n (%). n.s., not significant.
In light of this finding, in AD group, we compared CSF biomarkers (namely Aβ, tau, and Ptau), age at onset and male: female ratio in APOE ɛ4 carriers stratified for KL-VS status, in order to verify differences driven by the presence of KL-VS heterozygosis. Nevertheless, no significant differences were found (p > 0.05).
In a subgroup of subjects (Table 4), we performed a gene expression study in order to evaluate KL mRNA levels in patients and healthy controls.
Demographics of gene expression cohort
AD, Alzheimer’s disease; FTD, frontotemporal dementia; SD, standard deviation; n.p., not performed; n.a., not applicable.
As shown in Fig. 1, KL expression levels were significantly lower in AD and FTD patients compared with control group (AD mean Fold-regulation=– 4.286; FTD mean Fold-regulation=– 6.561; Ctrls mean Fold-regulation=0.8419; p = 0.0037) whereas no differences have been found between AD and FTD KL expression levels (p > 0.05)
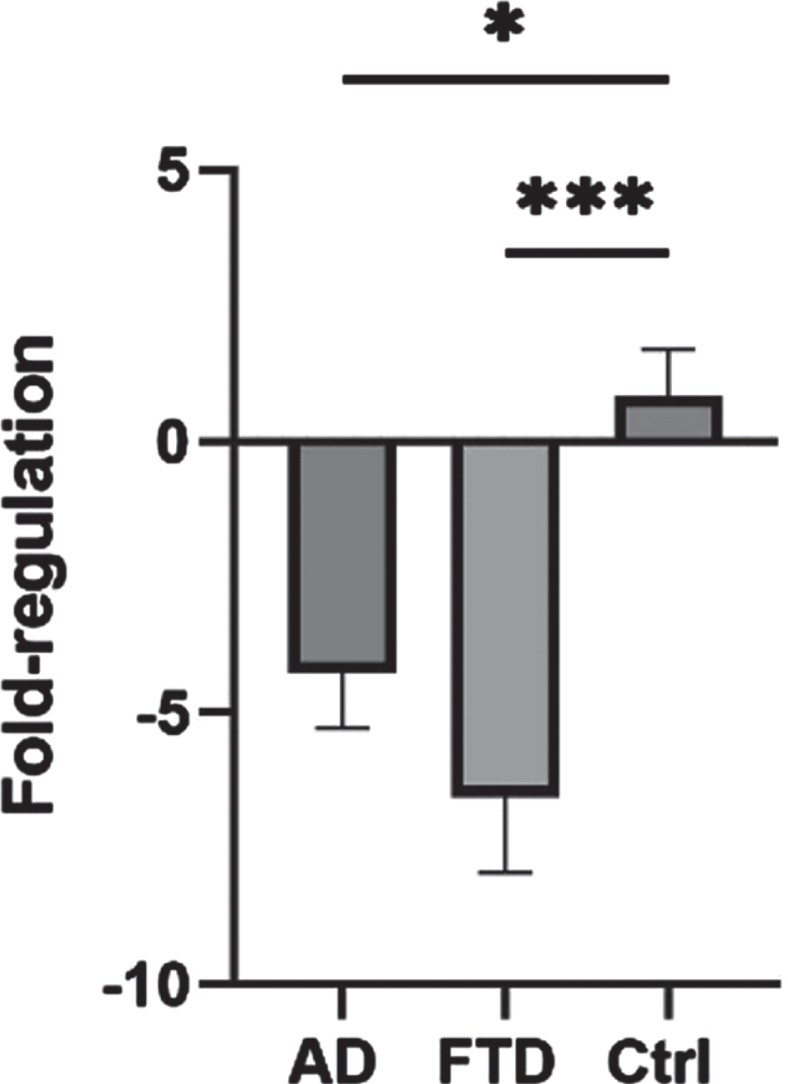
KL gene expression.
Mean fold-regulation±SEM of patients and controls.
In order to identify a possible effect of KL-VS status on gene expression level, we compared fold regulation in KL-VShet + and KL-VS het– . No significant differences were found in all the three groups. No significant differences were found stratifying for sex as well.
Correlation study
Moreover, we correlated KL expression levels with CSF biomarkers and clinical variables such as age at onset, gender, and disease duration. There was a positive correlation (p = 0.0181; 95%) between KL expression and CSF Aβ levels in AD patients: the latter were lower in patients with lower KL expression levels (Fig. 2), whereas no correlation has been found for CFS biomarkers and KL expression levels in FTD group.
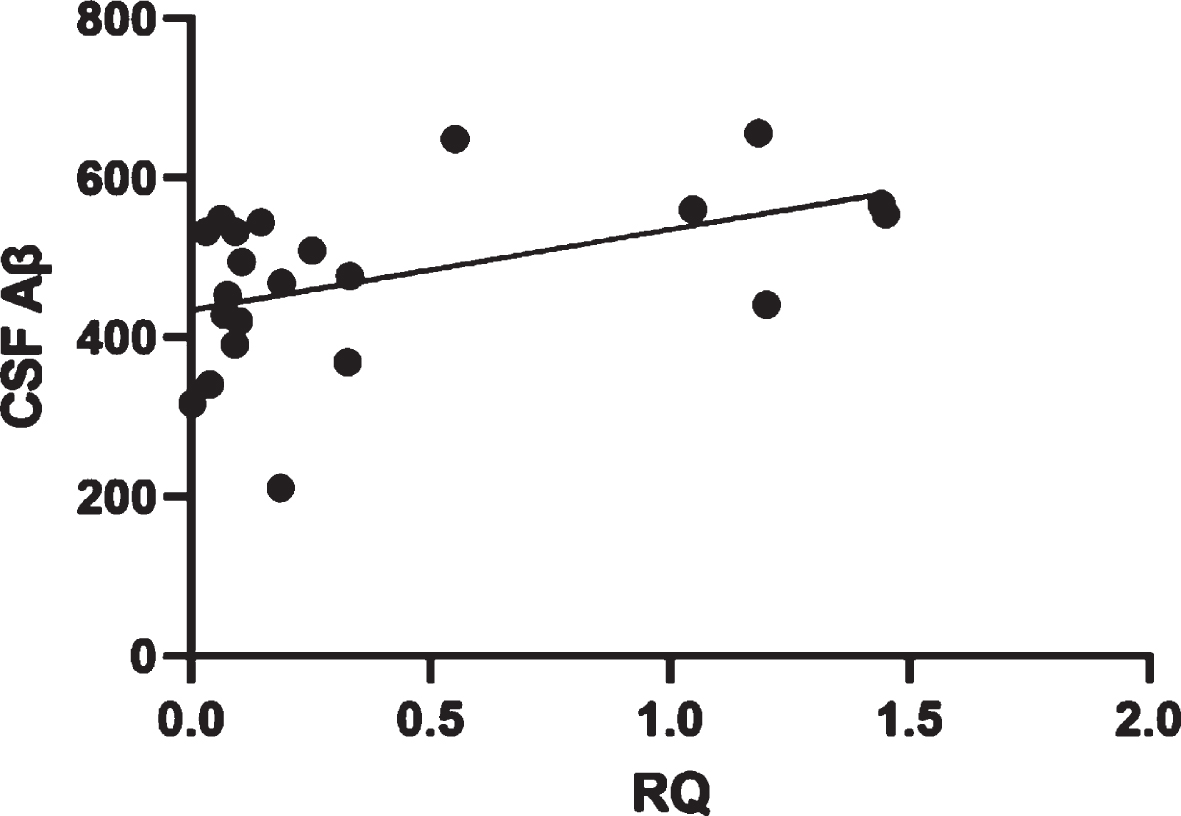
Correlation between CSF Aβ levels and KL expression levels in AD patients.
No significant differences were found when correlating RQ with the other clinical variables.
CSF Aβ levels expressed as pg/mL.
DISCUSSION
Since KL serendipitously discovery, rising evidence suggest its involvement in brain health. As reviewed by Hanson and colleagues [22], animal studies suggested multiple mechanisms for Klotho in the brain, including reducing inflammation, enhancing autophagy and amyloid clearance, all processes known to be involved in neurodegeneration.
Here, we reported low levels of KL gene expression in AD and FTD patients compared to control subjects, indicating a possible non-specific association with neurodegeneration.
Nevertheless, a limitation of the study is that measurements in peripheral blood cells may reflect several systemic conditions associated with neurodegenerative diseases (e.g., inflammation). In this regard, there is no definite evidence of blood-brain barrier permeability for KL and it has been shown, in Parkinson’s disease at least, that blood and CNS represent two separated compartments, showing different trends of KL expression and distinct relationship with neurodegeneration markers [23]
When evaluating the impact of KL-VS status on gene expression, we did not find any statistically significant difference between KL-VS carriers and non-carriers. Findings from literature report higher circulating levels of KL protein in KL-VShet +, whereas more recent data suggest that Klotho protein levels are associated with cognitive decline directly rather than depending on the KL-VS heterozygosity. In this scenario, it has to be acknowledged that so far none of the studies on Klotho consider the role of epigenetics, which may influence the level of gene expression. However, the inconsistency of our results with the literature could be due to the small size of the gene expression cohort.
We found a correlation between KL gene expression and CSF Aβ in AD patients: high expression levels were associated to higher Aβ levels. In this regard, studies in animal models revealed that overexpression of KL in the brain reduces Aβ burden [24, 25]. KL could therefore play a role in AD pathology and this corroborate its potential as therapeutic target.
To the best of our knowledge, this is the first study investigating KL in FTD patients. As already stated, we found low KL expression levels in FTD compared to control group. Its involvement in both AD and FTD could indicate a pleotropic effect of KL. Regarding controls, subjects were very well characterized in terms of frailty index and cognition, but we acknowledge that a few of them may have comorbidities such as main internal diseases (i.e., diabetes) which can influence KL expression [26, 27]. As proof of its pleiotropic nature, KL seems to be associated to several conditions. For instance, reduced KL gene expression levels or protein levels have been found in relapsing-remitting multiple sclerosis (RR-MS) [28], Parkinson’s disease [29], and in schizophrenia, possibly acting together with other detrimental factors. Therefore, the broad-spectrum neuroprotection of KL is intriguing since it expands the landscape of its therapeutic applicability. To date, several approaches are indeed being studied to increase KL expression (reviewed by Hanson and colleagues [22]). In this scenario, further investigating KL mechanisms would help to sharpen its therapeutic potential.
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
This work was supported by grants from the Italian Ministry of Health (Ricerca Corrente and RF-2018-12365333), Dino Ferrari Center and Fondazione Gigi & Pupa Ferrari Onlus. MS is supported by the Italian Ministry of Health, grant GR-2019-12369100. FS is supported by the Italian Ministry of University and Research (Joint Program on Neurodegenerative Diseases 2019 – project “DIPPA-FTD”).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The data supporting the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.