
Abstract
Background:
Neurodegeneration is a term describing an irreversible process of neuronal damage. In recent decades, research efforts have been directed towards deepening our knowledge of numerous neurodegenerative disorders, with a particular focus on conditions such as Alzheimer’s disease (AD). Human transferrin (htf) is a key player in maintaining iron homeostasis within brain cells. Any disturbance in this equilibrium gives rise to the emergence of neurodegenerative diseases and associated pathologies, particularly AD. Limonene, a natural compound found in citrus fruits and various plants, has shown potential neuroprotective properties.
Objective:
In this study, our goal was to unravel the binding of limonene with htf, with the intention of comprehending the interaction mechanism of limonene with htf.
Methods:
Binding was scrutinized using fluorescence quenching and UV-Vis spectroscopic analyses. The binding mechanism of limonene was further investigated at the atomic level through molecular docking and extensive 200 ns molecular dynamic simulation (MD) studies.
Results:
Molecular docking uncovered that limonene interacted extensively with the deep cavity located within the htf binding pocket. MD results indicated that binding of limonene to htf did not induce substantial structural alterations, ultimately forming stable complex. The findings from fluorescence binding indicated a pronounced interaction between limonene and htf, limonene binds to htf with a binding constant (K) of 0.1×105 M–1. UV spectroscopy also advocated stable htf-limonene complex formation.
Conclusions:
The study deciphered the binding mechanism of limonene with htf, providing a platform to use limonene in AD therapeutics in context of iron homeostasis.
INTRODUCTION
Several elements abundant in our bodies are responsible for essential physiological functions. Iron is among them, serving a critical role in regulation. It is both a necessary nutrient and a potent neurotoxin. Disrupted iron distribution impacts the iron-dependent biosynthetic pathway, ultimately resulting in an elevated iron pool that leads to oxidative stress. This oxidative stress is a pivotal factor in the progression of numerous neurodegenerative diseases [1]. Unbound ferrous iron is highly hazardous, as it produces reactive oxygen species (ROS) through its interaction with hydrogen peroxide. This interaction results in the generation of an environment abundant in oxidative stress and transforms hydrogen peroxide into a free radical. These ROS go on to assault large molecules such as lipids, proteins, and nucleic acids. It is crucial to diminish the presence of unbound ferrous in an environment, a goal that can be attained by managing iron homeostasis [2]. The pivotal figures in this process include human transferrin (htf) and ferritin. htf is a single-chain polypeptide consisting of 679 amino acids with multiple domains capable of iron binding [3, 4]. Once it forms a complex with iron, htf helps in disseminating iron throughout the bloodstream [5].
Numerous studies have demonstrated a correlation between elevated iron levels in the brain, an abundance of iron-binding proteins, and the onset of neurodegenerative conditions such as Alzheimer’s disease (AD) and Parkinson’s disease [6]. The presence of amyloid plaques is a defining trait of advanced-stage AD. This incurable ailment affects nearly 40 million individuals globally, making it the foremost dementia cause in humans and accounting for an approximate annual healthcare expenditure amounting to 820 billion dollars [7]. A lasting remedy for AD remains elusive, and considerable efforts are underway to develop potent drugs for its treatment [8]. Amyloid-β (Aβ) deposition, resulting in the formation of plaques, is a key characteristic of AD and is responsible for neuronal degeneration and cognitive deterioration [9]. Advancements in drug therapies have made it possible to somewhat manage AD, which was a challenging task in the past [10]. The characterization of Aβ remains the sole supportive biomarker for detecting AD, posing a significant obstacle to advancing AD therapy.
Natural compounds obtained from diverse origins, encompassing plants, animals, and fungi, have garnered significant interest as potential therapeutic agents for AD as they exhibit a range of pharmacological characteristics, including anti-inflammatory, antioxidant, and neuroprotective effects, which hold the potential to influence the pathological mechanisms linked to AD [11].
Limonene, a monoterpene predominantly found in the Rutaceae family, exhibits a range of biological properties, including antioxidant, anti-inflammatory, anticancer, antinociceptive, and gastroprotective attributes. Lately, there has been an increasing curiosity about investigating the pharmacological effects of limonene in the context of various chronic diseases [12]. This is primarily due to its capacity to mitigate oxidative stress and inflammation, as well as its role in regulating apoptotic cell death [13]. Several studies have been carried out that underscore the neuroprotective properties of limonene in various neurodegenerative conditions, viz. AD, multiple sclerosis, epilepsy, anxiety, and stroke [14–17]. There are various properties of limonene that makes it a promising candidate for development as a nutraceutical for preventative purposes, viz. favorable safety profile, and its diverse mechanisms of action position. Additionally, it can serve as an alternative or adjunctive agent to modern therapeutic drugs in the effort to both prevent and mitigate the progression of neurodegenerative diseases [18, 19].
In acidic conditions, iron is dissociated from this complex and translocated to the cytoplasm. Once separated, iron contributes to the assembly of Aβ oligomers, leading to detrimental effects, particularly in later phases when the generation of ROS is involved. htf acts to mitigate the toxicity of Aβ by capturing and retaining iron. htf fulfils a dual role: firstly, by sequestering iron, it impedes the formation of Aβ bonds, and then, by removing free iron, it aids in Aβ formation, thereby influencing the progression of AD [20]. Binding occurs between drugs and proteins, and in most cases, this binding is reversible. Understanding this interaction is crucial for explaining their mode of action in the body. Drugs form complexes with proteins and are primarily transported in the form of a protein-ligand complex. Various factors influence the binding process between proteins and drugs, and the affinity of drugs for proteins plays a significant role in bioavailability and the duration they remain in circulation. Studies that illustrate the binding of drugs with plasma proteins are essential for pharmacological drug profiling. These studies involve gaining insights into the molecular characteristics or fundamental categories of therapeutic drugs that interact with either target tissue proteins or plasma components [21].
MATERIAL AND METHODS
Materials
We procured htf and limonene from Sigma Aldrich, based in St. Louis, MO, USA. All the additional chemicals necessary for buffer preparation were sourced from HiMedia. Initially, the protein was dialyzed in 20 mM sodium phosphate, pH 7.4 buffer overnight to remove the impurities and then we prepared a stock solution of 5 mg/ml (equivalent to 62.5μM) htf. We employed suitable blanks as controls that were analyzed under identical conditions. In all spectroscopic assays, the spectra presented are the result of subtracting the corresponding blanks.
Molecular docking
The structural coordinates of htf were retrieved from the RCSB Protein Data Bank (Identifier: 3V83). Limonene was obtained from the PubChem database (CID: 22311). The structures were subjected to preprocessing using MGL AutoDock tools to ensure its compatibility with the docking process. Molecular docking studies were conducted to analyze the interactions between limonene and htf. AutoDock Vina-based InstaDock was employed for this purpose [22, 23]. The search space for the docking was set within the ‘conf. file’, allowing limonene to explore the entire htf and interact with its preferred binding site(s). During the process, the docking scores provide valuable insights into the affinity and efficiency of limonene with htf. To comprehensively visualize various interfaces between limonene and htf, Discovery Studio Visualizer was utilized [24]. This software facilitated the examination of the molecular interactions, enabling a deeper understanding of the binding mode between limonene and htf.
Molecular dynamic simulations
After conducting molecular docking analysis, we proceeded with all-atom MD simulations on htf in two different conditions: its free state and its bound conformation with limonene. These simulations were conducted under a constant temperature of 300 K. We utilized the GROMACS 2020 beta software package [25] and employed the charmm36-jul2022 force field for the MD simulations [26]. The ligand topology was generated through the CHARMM (CGenFF) program, supplemented with cgenff_charmm2gmx_py3_nx2.py. This approach allowed us to integrate the limonene ligand seamlessly with the htf coordinates. To create a realistic simulation environment, the systems were solvated within a 1 nm cubic box, employing the Simple-point charge 216 solvent model [27]. Subsequently, we performed a steepest descent approach using 1500 steps, ensuring an equilibrium state was reached [28]. Various GROMACS utilities, including gmx energy, gmx rms (root-mean-square deviation), gmx rmsf (root-mean-square fluctuation), gmx gyrate (radius of gyration), gmx sasa (solvent-accessible surface area), gmx sham (secondary structure assignment), and others, were employed to analyze the resulting MD trajectories. Graphical representations and figures derived from the MD simulation analysis were generated using XMGrace [29], facilitating the visualization of key findings and structural changes over the simulation.
UV-visible absorbance spectroscopy
The spectrophotometric measurement of absorbance spectra was carried out using protein sample without a ligand and protein samples with progressively increasing concentrations of the ligand. The spectrophotometer used was the Shimadzu UV-1700 (KYOTO JAPAN), with a 1 cm path length quartz cuvette. For this experiment, 4μM of the protein was utilized, and the ligand concentration was incrementally introduced to the protein solution. The concentration of the ligand was raised from 0 to 15μM.
Fluorescence quenching studies
To explore the binding strength between limonene and htf, a fluorescence-based quenching experiment was conducted using a Jasco spectrofluorometer (FP-6200). The analysis was performed following methodologies established in previous studies [9, 30]. The Inner filter effect was accounted for in all experiments, following established protocols [31]. Protein excitation was set at 280 nm to capture the comprehensive protein fluorescence, and spectra were examined within the range of 300–400 nm [32, 33]. The data were analyzed using the Modified Stern-Volmer (MSV) equation to determine and quantify the binding affinity of the protein for the ligand [33].
RESULTS
Molecular docking analysis
Molecular docking analysis was employed to assess the binding affinity and interactions between limonene and htf. The results revealed a robust binding interaction, signifying limonene’s potential as a promising binding partner of htf. The htf-limonene complex exhibited a significant docking score of –5.8 kcal/mol, coupled with a ligand efficiency of 0.58 kcal/mol/non-H atom. To get deeper insights into the binding mechanisms, we conducted a comprehensive examination of the docking results (Fig. 1). Our analysis highlighted limonene’s preferential binding site and its intricate interactions with htf. Specifically, limonene demonstrated substantial engagement with residues located within the iron and hydrogencarbonate-binding domain (Fig. 1A).

htf-limonene interaction. A) The binding between limonene and htf. B) Surface representation of the htf binding pocket with limonene. C) Two-dimensional view illustrating the interactions of limonene with htf.
Within this binding pocket, limonene formed two crucial Alkyl and Pi-Alkyl bonds with Phe414, Ala472, Pro527, and Tyr534, further setting its binding to htf. The surface representation vividly illustrated limonene’s occupancy within the internal cavity of htf (Fig. 1B). Additionally, we observed several essential interactions with multiple htf residues, contributing to the stability of the complex (Fig. 1C). Our findings align with prior research documenting the binding of iron and hydrogencarbonate to htf through a similar set of residues [34, 35]. This discovery reinforces our proposal that limonene’s binding to htf within the binding pocket obstructs the entry of iron and hydrogencarbonate, consequently disrupting the catalytic function of htf. In summary, the molecular docking analysis and detailed examination of the htf-limonene complex. These results provide valuable insights into the binding mechanism of limonene with htf, offering promise for further exploration in drug development and therapeutic applications.
Structure dynamics analysis
Assessing structural changes in a protein induced by small molecule binding is critical in understanding their interaction [34]. We employed RMSD, a widely-used metric for quantifying structural deviations from the native conformation [36], to evaluate the impact of limonene binding on htf. The RMSD analysis revealed no substantial differences in the RMSD values between the unbound htf and the htf-limonene complex, suggesting that limonene binding did not make significant structural alterations in htf (Fig. 2A). The RMSD plot indicated that the htf-limonene complex reached a stable equilibrium, indicating the overall stability of the complex. These findings suggest that limonene interacts with htf while maintaining the protein’s structural stability. To further investigate the dynamics within htf, we calculated the RMSF for all its residues in htf and htf-limonene bound systems (Fig. 2B). The RMSF analysis revealed some minor fluctuations in multiple parts of htf. While the average RMSF remained consistent upon limonene binding, certain residues in the loop regions, primarily in the N-terminal section, exhibited increased fluctuations. These fluctuations are likely due to ongoing vibrations during the simulation.
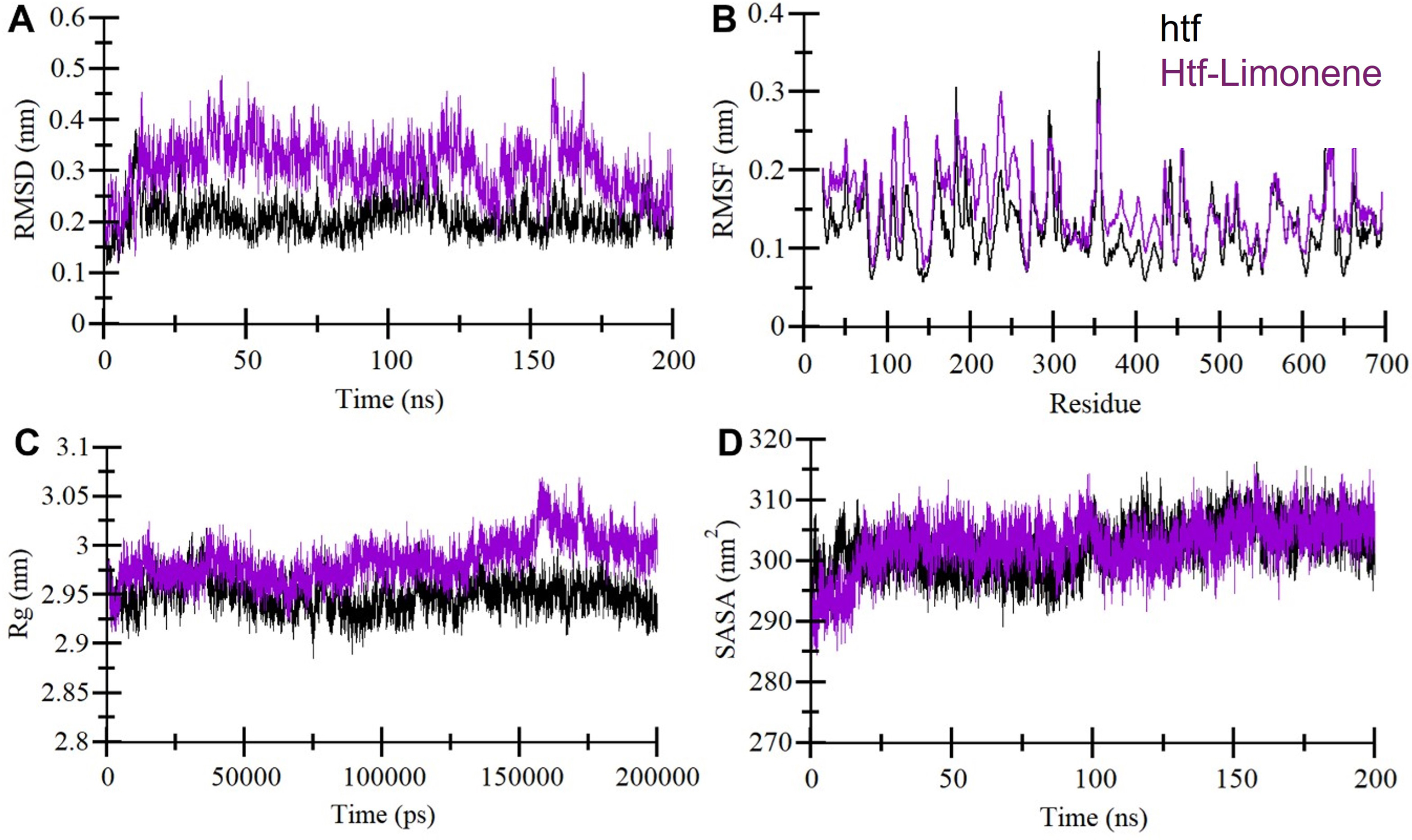
Structural dynamic and compactness in htf upon binding with limonene: A) Trajectory depicting the root mean square deviation (RMSD) of htf over time. B) Chart illustrating residual fluctuations (RMSF) in htf, both in its unbound state and when bound to limonene. C) Evolution of the radius of gyration (Rg) with time. D) Variations in the solvent accessible surface area (SASA) of htf throughout the simulation period. The data was obtained from 200 ns MD simulations, with black representing unbound htf and purple denoting the htf-Limonene complex.
The Rg provides insights into a protein’s conformational shape and folding mechanism [37]. In our analysis, Rg values for htf and the htf-limonene complex showed almost a similar trend during the simulation (Fig. 2C). Notably, the incorporation of limonene resulted in a slightly increased Rg for htf compared to its unbound state, although this change was minimal and consistent throughout the simulation. The overall packing of htf remained structurally stable, indicating that limonene’s influence had a negligible impact on the protein’s structural integrity. Further, we computed average SASA values for htf and the htf-limonene complex during the simulation (Fig. 2D). These values also showed a similar distribution, with a minor increase before 100 ns observed in the SASA for the htf-limonene complex compared to unbound htf. This slight change suggests slight conformational adjustments within htf due to limonene binding. Overall, the structural dynamics analysis indicates that limonene binding to htf does not induce significant structural upheaval. Instead, it maintains the stability of the protein while inducing minor fluctuations in specific residues.
Stabilization of htf-limonene complex
Intramolecular hydrogen bonds are vital in maintaining structural stability in proteins [38]. To evaluate the stability of htf, we quantified the number of hydrogen bonds within htf before and after limonene binding (Fig. 3A). The results indicate a minor increase in the number of hydrogen bonds following limonene interaction, suggesting higher compactness and stability due to limonene binding. Additionally, we evaluated the hydrogen bonds formed between htf and limonene during the simulation. Intriguingly, the analysis revealed the presence of up to three hydrogen bonds between htf and limonene (Fig. 3B). Limonene was observed to occupy htf’s binding pocket, engaging in conventional hydrogen bonds with some degree of fluctuations. Notably, binding through a single hydrogen bond exhibited relatively fewer fluctuations. These findings indicating the formation of intermolecular hydrogen bonds between htf and limonene. The presence of these interactions plays a pivotal role in stabilizing the htf-limonene complex. These interactions not only contribute to the overall stability of the complex but also suggest a specific and directed binding mode between limonene and htf. Overall, the analysis underscores the importance of hydrogen bond interactions in maintaining the stability of the htf-limonene complex.
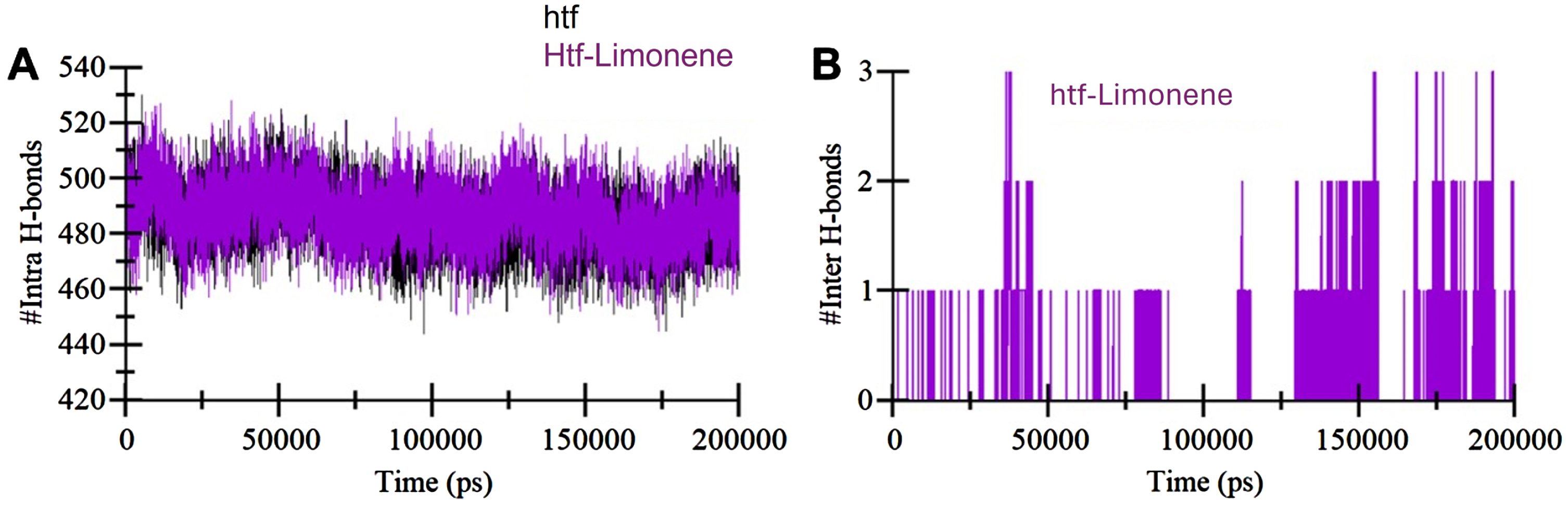
Temporal development of hydrogen bonds: A) The changes in hydrogen bonds within htf itself over time. B) Hydrogen bonds established between limonene and htf.
Principal component analysis
Principal component analysis (PCA) is a useful approach for examining global motions and conformational landscape of a protein [39]. Herein, PCA was employed to analyze the trajectories of htf and the htf-limonene complex, shedding light on their conformational dynamics. Figure 4 presents a summary of the conformational sampling of htf and the htf-limonene complex within the essential subspace (Fig. 4A). The observable pattern indicates that unbound htf explores phase spaces, displaying a large stable state. In the presence of limonene, htf exhibits a broader spectrum of conformational states, highlighting the influence of limonene on the protein’s dynamics. However, limonene does enhance the variety of conformational states in htf (Fig. 4B). These findings indicate that limonene binding does not influences the conformational dynamics of htf, preserving the overall stability of the complex.
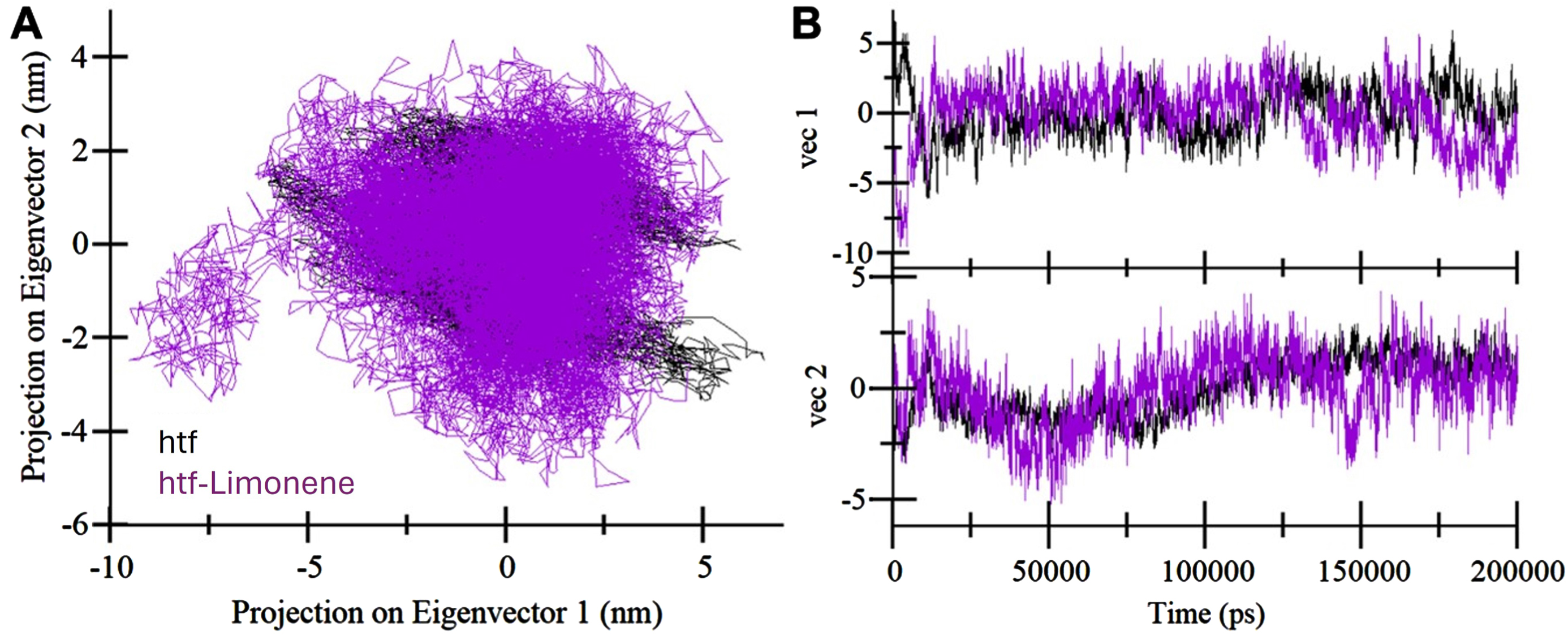
Exploring principal component analysis (PCA): A) 2D eigenvector projections displaying distinct orientations of htf throughout the trajectory. B) Trajectory projections onto eigenvectors over time. Black represents unbound htf, and purple represents the htf-limonene complex.
Free energy landscape analysis
To gain a deeper understanding of the complex molecular arrangements within our systems, we examined the free energy landscapes (FELs) from the simulated trajectories [40]. The FEL analysis delivers valuable understandings of the conformational dynamics and stability of htf and the htf-limonene complex (Fig. 5). For the htf’s FEL, two prominent global minima were evident, primarily confined in two basins (Fig. 5A). The presence of multiple energy minima in the FEL highlights the complex’s capacity to explore diverse conformational states. However, with the introduction of limonene, a major change in conformational tendencies became apparent (Fig. 5B). This indicates that bound htf predominantly adopts a specific conformational state with a stable energy minimum. This analysis offers vital insights into the dynamic conformational behaviors and stability of htf and the htf-limonene complex.
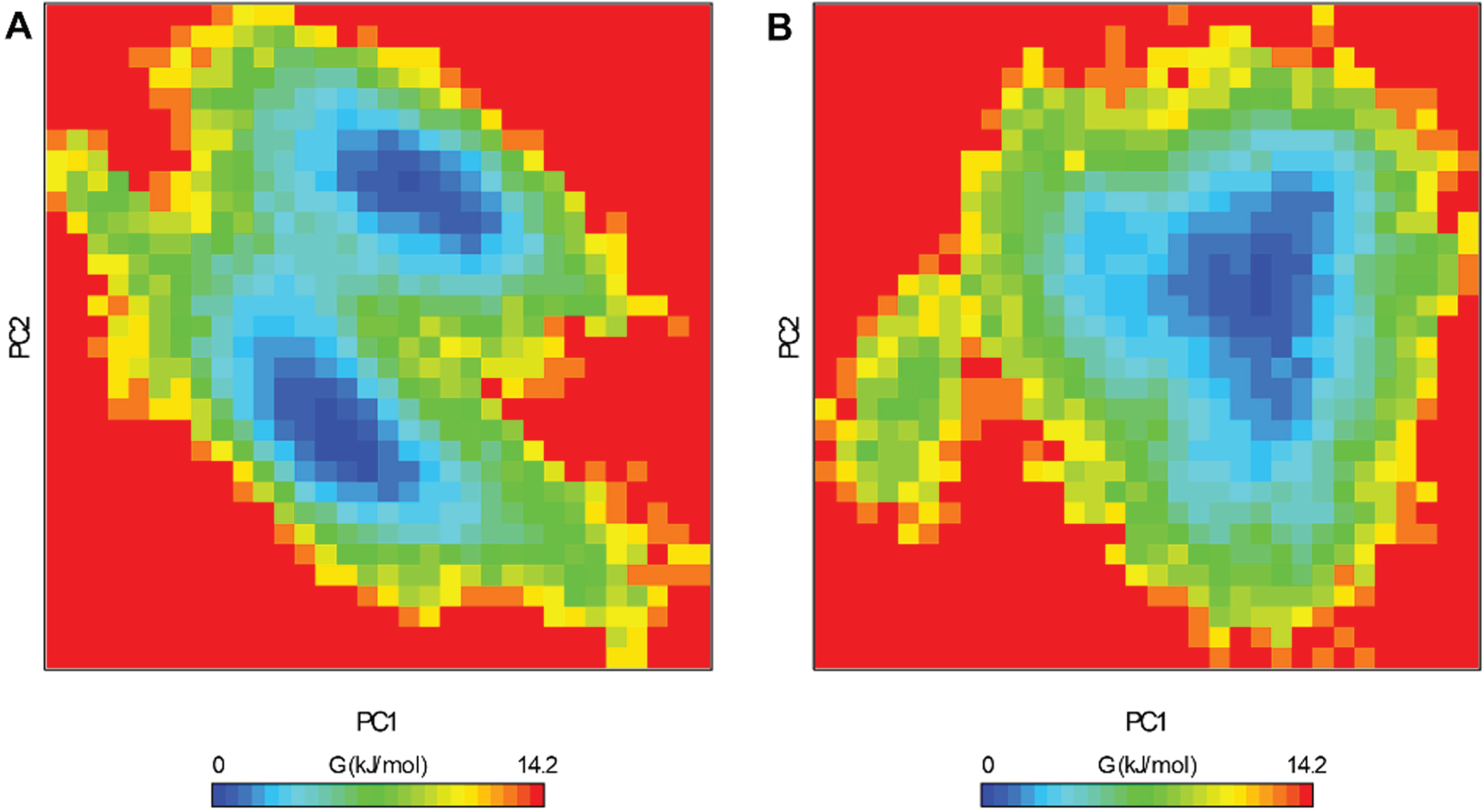
Free energy landscape obtained for (A) Unbound htf and (B) htf-limonene complex.
UV Absorbance measurements
UV-Vis absorption spectroscopy serves as a valuable tool for identifying the formation of complexes and the dynamic interactions between proteins and ligands. Complex formation is considered a static process that transpires at the ground state, impacting the electronic characteristics of the host protein. These alterations are evident through shifts in the UV-Vis absorption spectra. However, in situations where dynamic quenching predominates, it is expected that no observable changes will occur in the absorption spectra due to the interaction taking place at the excited state of the fluorophore. Therefore, UV-vis spectroscopy is commonly utilized for the characterization of protein-ligand complexes [27]. We conducted a titration of htf using a range of limonene concentrations (0–15μM) and observed a hyperchromic effect in the UV-visible spectra of htf, with alterations in peak position (Fig. 6). The obtained data was subjected to fitting using Modified Beer Lambert’s equation, and a double reciprocal plot was created, 1/(A – A0) versus 1/C (Fig. 6B). The binding constant (K) for the htf-limonene complex was determined by calculating the ratio of the intercept to the slope of this plot, as illustrated in Fig. 6B. The binding constant (K) for the htf-limonene complex was determined to be 4.7×104 M–1, indicating that limonene forms a stable complex with htf and exhibits a strong binding affinity.
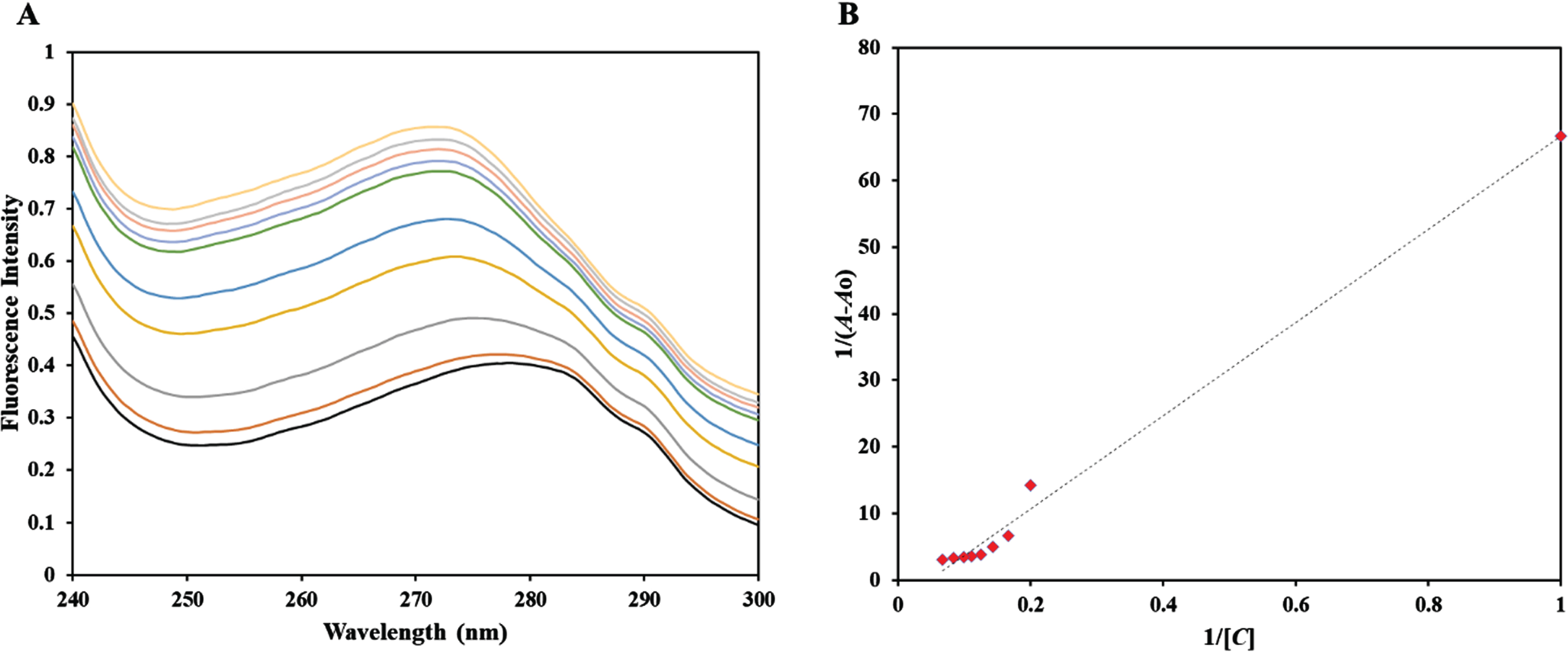
A) UV-Vis absorption spectra of htf in the absence and presence of limonene at concentrations ranging from 0 to 15μM. B) It presents a double reciprocal plot of the htf-Limonene complex.
Fluorescence quenching assay
Fluorescence-based binding assays are widely used to confirm the binding of a ligand to a protein, as they provide valuable insights into the protein-ligand complex by revealing a range of binding parameters [28]. This approach has been widely adopted in numerous studies to uncover the binding mechanisms of significant ligands and drugs with transferrin and other medically relevant proteins, aiming to clarify the characteristics of the binding process [41, 42]. Upon ligand binding to protein, it alters the local environment surrounding intrinsic fluorophores, predominantly tryptophan and these alterations are subsequently replicated in the corresponding fluorescence spectra. Fluorescence quenching is a phenomenon characterized by a reduction in the fluorescence of the protein as the ligand concentration increases [41]. The intrinsic fluorescence of htf exhibits a high level of sensitivity to its micro-environment and thus with increasing limonene concentration, microenvironment around htf changes that is reflected in the corresponding changes in the fluorescence spectra. Under its natural conditions, htf exhibited a distinct fluorescence peak around 335 nm when excited at 280 nm. Figure 7 depicts how varying concentrations of limonene affect the fluorescence intensity of native htf. The obtained data was fitted into equation to obtain the binding constant (K) (from intercept of this plot) of htf-limonene complex. The binding affinity of the complex was found to be K = 0.1×105 M–1, validating the earlier in silico observations, implying that limonene binds to htf with a significant affinity forming a stable complex.
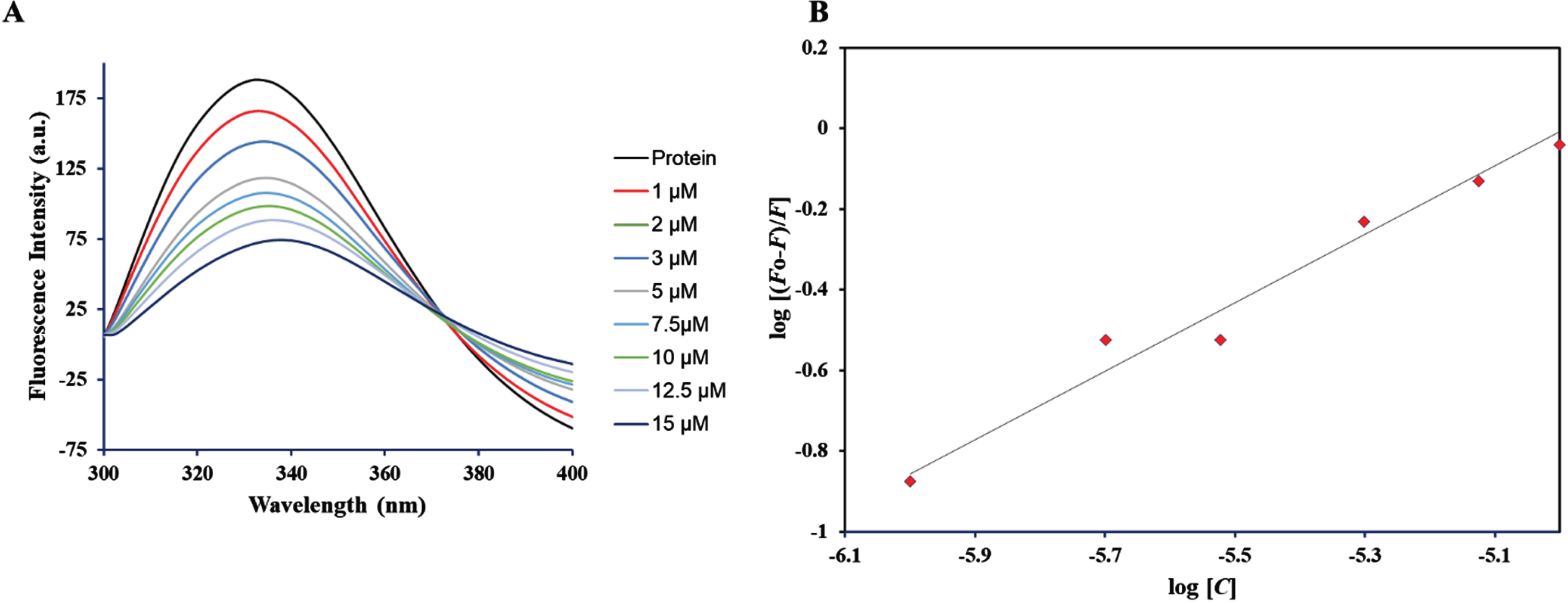
Depiction of fluorescence intensity for unbound htf and htf in the presence of varying limonene concentrations (ranging from 0 to 15μM).
DISCUSSION
The present study provides a comprehensive investigation into the interactions between limonene and htf, utilizing a multifaceted methodology that combines docking MD simulations and experimental approaches. The docking results are indicative of a robust interaction between limonene and htf, suggesting its potential as a htf binding partner. Furthermore, the investigation pinpointed specific amino acid residues within htf’s iron and hydrogencarbonate-binding domain, which played a crucial role in these interactions [43]. Notably, limonene established important interactions with other residues. These interactions align with previous findings, where similar interactions have been observed with iron and hydrogencarbonate [35]. It is hypothesized that limonene’s occupancy could hinder access to iron and hydrogencarbonate. The subsequent phase of the study involved comprehensive all-atom MD simulations, providing an in-depth exploration of the structural dynamics of the htf-limonene complex. The RMSD and RMSF analyses demonstrated that limonene binding did not induce major deviations in the htf structure. This observation suggests that limonene binding primarily affects local conformational dynamics rather than global structural changes. Additionally, investigations of Rg and SASA along with the hydrogen bond analysis indicated minor conformational modifications and alterations in solvent exposure due to limonene binding.
Further, UV-visible spectroscopy revealed a noticeable increase in absorbance of native htf as the limonene concentration increased, indicating the binding of limonene to htf and formation of a complex. Fluorescence spectra exhibited quenching as the limonene concentration increased, indicating the formation of the htf-limonene complex. The binding mechanism proposed here suggests that limonene and its derivatives hold promise as potential therapeutic interventions in neurodegenerative diseases, including AD. The results not only strengthen the case for limonene as a htf modulator but also elucidate the elaborate molecular details governing this interaction. The stability of the htf-limonene complex, the local conformational changes observed, and the alteration of the hydrogen bonding network all contribute to a more comprehensive understanding of the molecular mechanisms involved.
Conclusion
In summary, this study offers valuable insights into the binding mechanism, conformational stability, and contributes to our understanding of the potential use of limonene in the therapeutics of neurodegenerative diseases. The insights gained from this study serve as a solid foundation for future research endeavors in the development of novel therapeutic agents and interventions for AD therapeutics.
AUTHOR CONTRIBUTIONS
Mohammed Alrouji (Conceptualization; Formal analysis; Investigation; Methodology; Writing – original draft); Sabina Yasmin (Data curation; Formal analysis; Investigation; Methodology; Validation; Writing – original draft); Mohammad Furkan (Investigation; Methodology; Validation; Visualization; Writing – original draft); Fahad A. Alhumaydhi (Investigation; Methodology; Validation; Visualization; Writing – original draft); Sharaf E. Sharaf (Investigation; Methodology; Software; Validation; Writing – original draft); Rizwan Hasan Khan (Formal analysis; Investigation; Methodology; Validation; Visualization; Writing – review & editing); Anas Shamsi (Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Supervision; Validation; Visualization; Writing – original draft).
Footnotes
ACKNOWLEDGMENTS
The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University for funding this work through a small research group program under grant number RGP. 1/219/44. MA would like to thank the Deanship of Scientific Research at Shaqra University for supporting this work. AS is grateful to Ajman University for supporting this publication.
CONFLICT OF INTEREST
Anas Shamsi is an Editorial Board Member of this journal but was not involved in the peer-review process of this article nor had access to any information regarding its peer-review.
All other authors have no conflict of interest to report.
DATA AVAILABILITY
The data supporting the findings of this study are available within the article.