
Abstract
BACKGROUND:
Black raspberry is a vital fruit crop with a high antioxidant function. MADS-box genes play an important role in the regulation of fruit development in angiosperms.
OBJECTIVE:
To understand the regulatory role of the MADS-box family, a total of 80 MADS-box genes were identified and analyzed.
METHODS:
The MADS-box genes in the black raspberry genome were analyzed using bioinformatics methods. Through an analysis of the promoter elements, the possible functions of different members of the family were predicted. The spatiotemporal expression patterns of members of the MADS-box family during black raspberry fruit development and ripening were systematically analyzed.
RESULTS:
The genes were classified into type I (Mα: 33; Mβ: 6; Mγ: 10) and type II (MIKC*: 2; MIKCC: 29) genes. We also obtained a complete overview of the RoMADS-box gene family through phylogenetic, gene structure, conserved motif, and cis element analyses. The relative expression analysis showed different expression patterns, and most RoMADS-box genes were more highly expressed in fruit than in other tissues of black raspberry.
CONCLUSIONS:
This finding indicates that the MADS-box gene family is involved in the regulation of fruit ripening processes in black raspberry.
Introduction
Rosaceae is an important angiosperm family that consists of more than 100 genera and 3000 species, including many economically important species, such as apple, pear, strawberry and rose [1]. Rosaceae also contains plants with highly unique fruits, such as drupe, kernel, and achene. Rubus is one of the largest and most diverse genus in this family, and its fruits are characterized by the aggregation of small drupes on the receptacle [2]. Specifically, black raspberry (Rubus occidentalis L.) is a vital fruit crop native to eastern North America [3]. Black raspberry is valued for its unique flavor and exhibits potential health benefits [4]. Black raspberry fruits contain anthocyanins and other phenolic compounds at higher concentrations than those found in fruits of red raspberry and blackberry [5–7]. The demand for this fruit crop as a rich source of antioxidants is increasing [7–9]. Despite the importance of black raspberry, little is known about the molecular and genetic mechanisms that control fruit development and ripening in this fruit crop. The fruit ripening process involves a series of physiological and biochemical changes, such as the accumulation of pigments, the softening of the fruit, and the formation of aroma and flavor substances [6]. A study of the MADS-box family in black raspberry will facilitate further studies on the molecular mechanisms of development and ripening of black raspberry fruit.
MADS-box genes play an important role in ovule development, flower organ characteristics, fruit maturation, plant growth and development and signal transduction processes in eukaryotes, particularly the regulation of flower organs and fruit development in angiosperms [10–13]. Among them, Lyu [12] had found two artificially selected MADS genes that control meristem maturation and inflorescence development. Dong et al. [13] showed that a tomato MADS-box transcription factor, SlDADS, acts as a negative regulator of fruit ripening. MADS-box family proteins can be roughly divided into two types according to their structure and evolutionary developmental states: type I and II proteins. Type I proteins are divided into Mα, Mβ, and Mγ. In general, type II proteins consist of six introns and seven exons, and the proteins encoded by MIKC-type genes mainly include four domains, namely, the highly conserved MADS (M) domain, the moderately conserved keratin-like (K) domain, the more conserved intervening (I) domain, and a changeable C-terminal (C) end [11, 14]. According to differences in their gene sequence and structure, structural MIKC-type MADS-box genes can be further divided into MIKC* and MIKCc types [15].
Previous genomic studies have provided an understanding of the roles of the MADS-box family in Arabidopsis [16], rice [17], soybean [18], cucumber [19], and poplar [20], but the role of MADS-box genes in black raspberry has not been reported. The black raspberry genome was recently published [4], which provides an opportunity to analyze the MADS-box gene family and its role in fruit development and ripening. Studying MADS-box transcription factors in black raspberry is conducive to exploring and analyzing the mechanism regulating the physiological development and maturity of fruits and can provide valuable information for the biotechnological breeding of Rosaceae plants and other species.
Materials and methods
Identification, distribution and characterization of MADS-box genes in black raspberry
The black raspberry genome was downloaded from the Genome Database for Rosaceae (Rubus occidentalis whole genome v3.0 assembly & annotation) (https://www.rosaceae.org/analysis/268) [21]. Using the hidden Markov model (HMM), files corresponding to MADS-box sequences (PF00319) in both FASTA and Stockholm formats were downloaded from the Pfam database (http://pfam.xfam.org/). To obtain candidate proteins, an alignment of MADS-box seed sequences in Stockholm format was generated using a tool in the HMMER program (hmmbuild), and all black raspberry proteins in the generated HMM model were searched using another tool (hmmsearch) with the default parameters [22]. Additionally, the FASTA profile downloaded from the Pfam website was aligned with black raspberry protein sequences using the BLASTP (e-value = 1-3) method [23]. Candidate black raspberry MADS-box genes were obtained by taking the union set and removing duplicates from the above-mentioned two sets of results. To further verify the accuracy of the results, the obtained candidate MADS-box gene sequences were submitted to the SMART website (http://smart.embl.de/) to confirm that the conserved MADS-box domain was present in each protein sequence [24]. Genes without the M domain or with an incomplete M domain were then eliminated. Moreover, for the prediction of gene amplification, the similarity among the aligned regions was set to≥70%.
The distributions of all identified MADS-box genes were plotted on black raspberry chromosomes using MapInspect software. A conserved sequence logo (60 aa) was generated from the candidate black raspberry MADS-box genes using the web-based application WebLogo3.7.4 (http://weblogo.threeplusone.com) with the default parameters [25]. In addition, the sequence length, molecular weight (MW), theoretical isoelectric point (PI), instability index and grand average of hydropathicity (GRAVY) for these Rubus occidentalis MADS-box (RoMADS) genes were obtained using the ExPASy ProtParam tool (https://web.expasy.org/protparam/). We also used the SOPMA program (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html) to predict the secondary structure of each protein.
Phylogenetic, conserved motif and gene structure analyses of the RoMADS-box gene family
In general, all MADS-box genes can be divided into two categories (type I: M-type and type II: MIKC-type). To better classify these genes by subgroup, a multiple sequence alignment of various sequences, including MADS-box genes from Arabidopsis thaliana [26] and conserved RoMADS-box domain-containing proteins, was performed using Muscle, and a neighbor-joining tree with 1000 bootstrap replicates was then built using MEGA 7.0 based on the alignment. To better reveal the structural features of RoMADS proteins, we predicted the conserved motifs in full-length RoMADS proteins using the Multiple Expectation Maximization for Motif Elicitation (MEME) online tool (http://meme-suite.org/) with the default parameters, with the exception that the number of motifs was set to 10 [27]. Moreover, the intron-exon structures of all black raspberry genes were contained in the file “Rubus_occ_V3.genes.gff3”. An intron-exon structure diagram of all RoMADS-box genes was obtained from the online tool GSDS (http://gsds.cbi.pku.edu.cn/).
Promoter and protein-protein interaction analyses of RoMADS-box genes
TBtools software [28] was used to extract the 2000-bp sequence upstream of the coding sequence (ATG), and the extracted sequence was analyzed using the online software PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) [29]. The DNA sequences of cis-acting elements on the side of the target genes that can be identified and specifically bound by transcription factors were then predicted. Protein-protein interaction (PPI) data were obtained from the online database STRING (https://string-db.org/cgi/info.pl) [30].
Plant material and RNA extraction
Stems, leaves, apex, and fruits at different developmental stages (green, red and purple-black stages) were collected from three different black raspberry plants of ‘Bristol’, which were used as biological replicates, from the Nanjing Lishui White Horse Industrial Park of the Institute of Botany (Jiangsu Province, Chinese Academy of Sciences). The plant materials were stored at –80°C for RNA extraction. Total RNA was extracted using the Universal Plant Total RNA Extraction Kit (Bioteke, Beijing, China). After the quality and purity were checked, the total RNA was reverse-transcribed using the HiFiScript cDNA Synthesis Kit (CWBIO, Shanghai, China) according to the manufacturer’s instructions. The resulting cDNA was used for further experiments.
Expression analysis of RoMADS-box genes
We used Oligo 6.0 to design primers specific for RoMADS-box genes, and the details of this process are provided in Table S1. The relative expression levels were calculated using the 2–ΔΔCT method. Each 15-μL reaction volume consisted of 7.5μL of TB Green Premix Ex Taq II, each primer at a concentration of 10μM, 0.4μL of ROX Reference Dye II, and 1μL of cDNA template, and the remaining volume was composed of ddH2O and TB Green Premix Ex Taq II (Tli RNaseH Plus) (Takara, Dalian, China). qRT-PCR was performed using an ABI ViiA 7 real-time PCR platform according to the following cycling protocol: denaturation at 95°C for 30 s, 40 cycles of 95°C for 3 s and 60°C for 30 s, 95°C for 10 s, 65°C for 5 s and 95°C for 5 s. All data analyses were performed using data from three biological replicates and IBM SPSS Statistics version 22.
Results
Forty-nine RoMADS-box genes were classified as type I, and the remaining 31 RoMADS-box genes were type II. All (80) MADS-box genes were distributed across the seven chromosomes of the black raspberry genome and named RoMADS1 to RoMADS80 based on their position on the chromosomes (Fig. 1). A high frequency of segmental duplication was observed, and a total of six genes contained corresponding homologous segments (Fig. 1). The pI values varied greatly from 4.58 to 10.67 (Table 1). Seven MADS-box proteins (RoMADS13, RoMADS14, RoMADS19, RoMADS27, RoMADS41, RoMADS42, and RoMADS63) were stable, and all remaining 73 proteins were unstable, which indicated that most of the MADS-box proteins are unstable.
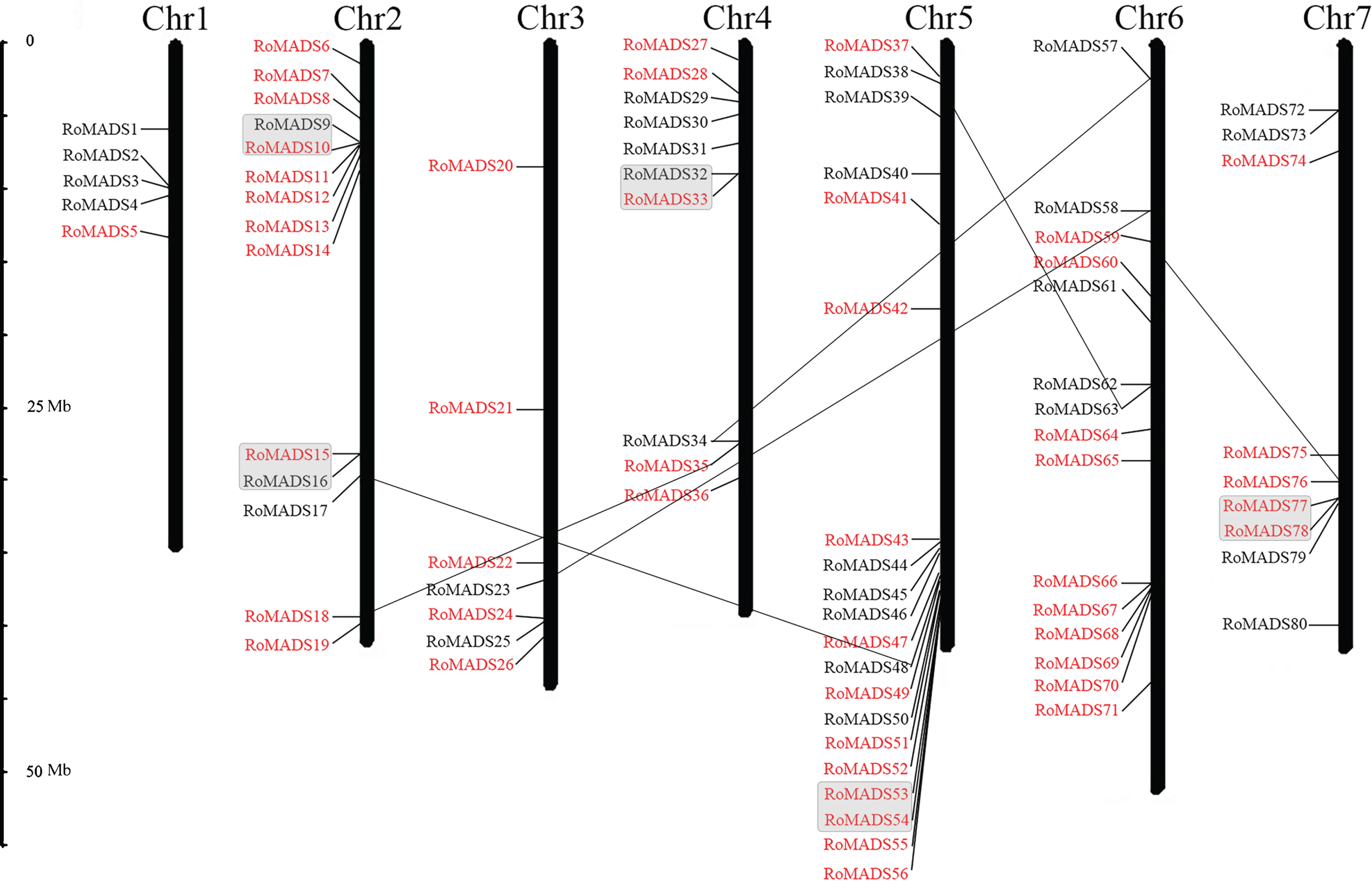
Chromosomal locations of the MADS-box genes on the seven chromosomes of the black raspberry genome. Chrs1–7 represent the seven chromosomes. The positions of the chromosomes at which each MADS-box gene is located are displayed proportionately. The black lines on bars indicate the locations of MADS-box genes. The type I and II MADS-box genes are shown in red and black, respectively. The tandemly duplicated genes are joined with a gray box. Genes that show segmental duplications are indicated by black lines.
Physicochemical characteristics of MADS-box genes in black raspberry
Notes: MW, molecular weight; pI, theoretical isoelectric point; GRAVY, grand average of hydropathicity; Hh, alpha helix; Ee, extended strand; Cc, random coil.
According to the presence or absence of the K-box domain, MADS-box family members were divided into two types (types I and II). A phylogenetic tree of these two types of MADS-box protein sequences in black raspberry and A. thaliana was constructed using the neighbor-joining method (Fig. 2). Based on the phylogenetic tree analysis, the 49 type I RoMADS-box genes were divided into three clades: Mα, Mβ, and Mγ. Clade I (Mα) contained 33 type I RoMADS-box genes, clade II (Mβ) contained six type I RoMADS-box genes, and clade III (Mγ) contained 10 type I RoMADS-box genes. Moreover, the type II genes could be divided into two clades: MIKC*, which contained two type II RoMADS-box genes, and MIKCC, which contained 29 type II members.
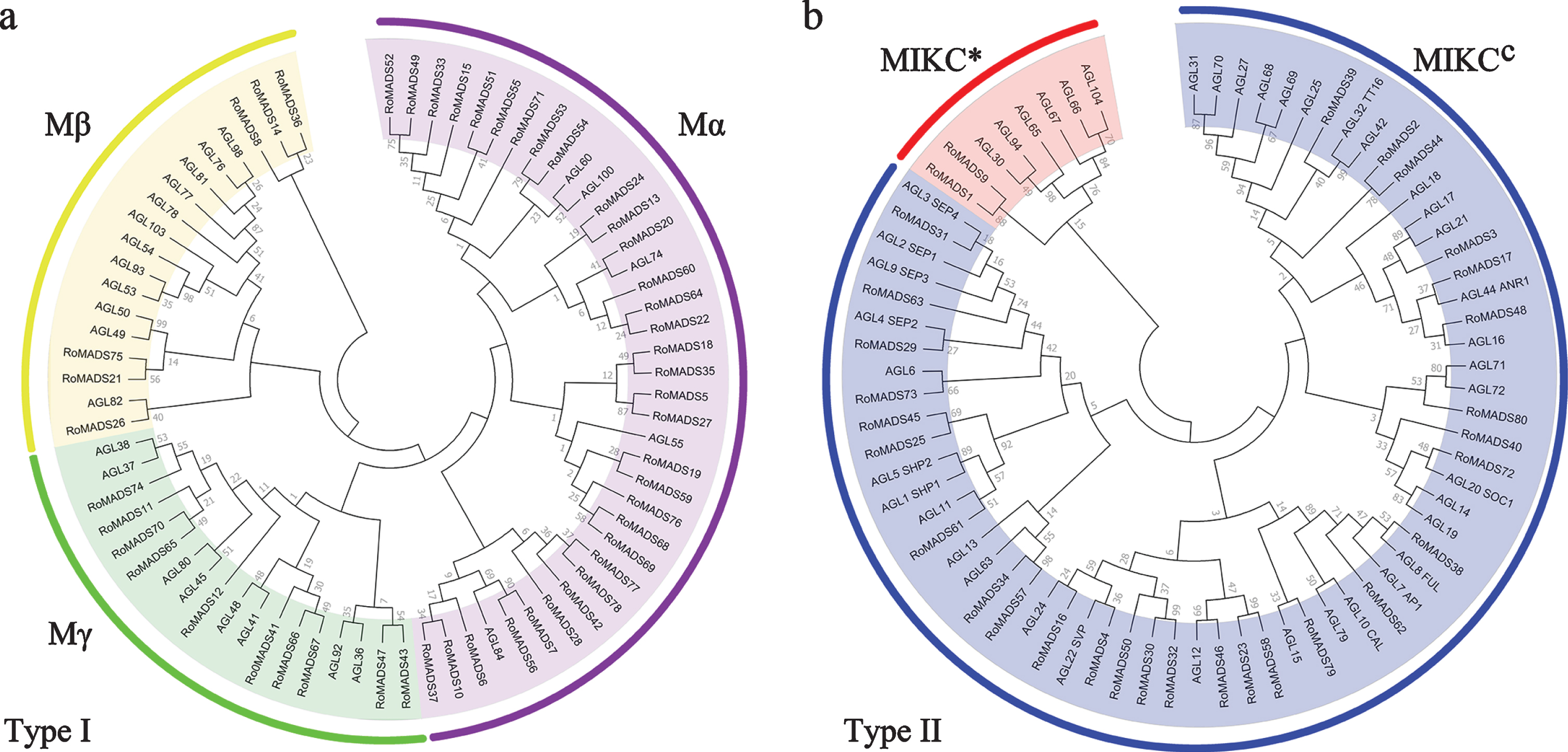
Phylogenetic tree analysis of type I (a) and type II (b) MADS-box genes. These genes from black raspberry and Arabidopsis thaliana were analyzed using MEGA 7.0 software with the neighbor-joining method and 1000 bootstrap replicates.
In addition, we compared the number of RoMADS-box gene family members in black raspberry with those in other species and found that several angiosperms, including monocotyledons such as rice and dicotyledons such as Arabidopsis, tobacco, poplar and apple, contained a higher number of members of the MADS-box gene family than gymnosperms (Table 2). Thus, the number of MADS-box gene family members varies greatly among different species.
Number of MADS-box genes in different species
Phylogenetic trees were constructed (Fig. 3a), and conserved motifs were identified (Fig. 3b) using type I and II MADS-box protein sequences. We then used the corresponding nucleic acid sequences for the analysis of RoMADS-box gene structures. To further investigate the characteristic regions of RoMADS proteins, the conserved motifs among the 80 RoMADS proteins were analyzed using the online program MEME. We constructed a schematic diagram of 10 conserved motifs to characterize the structures of the RoMADS proteins. The results from the motif analysis showed that motifs 1 and 2 in type I proteins and motifs 1 and 3 in type II proteins were highly conserved. Motif 1 represents a typical MADS-box domain (Fig. S2).
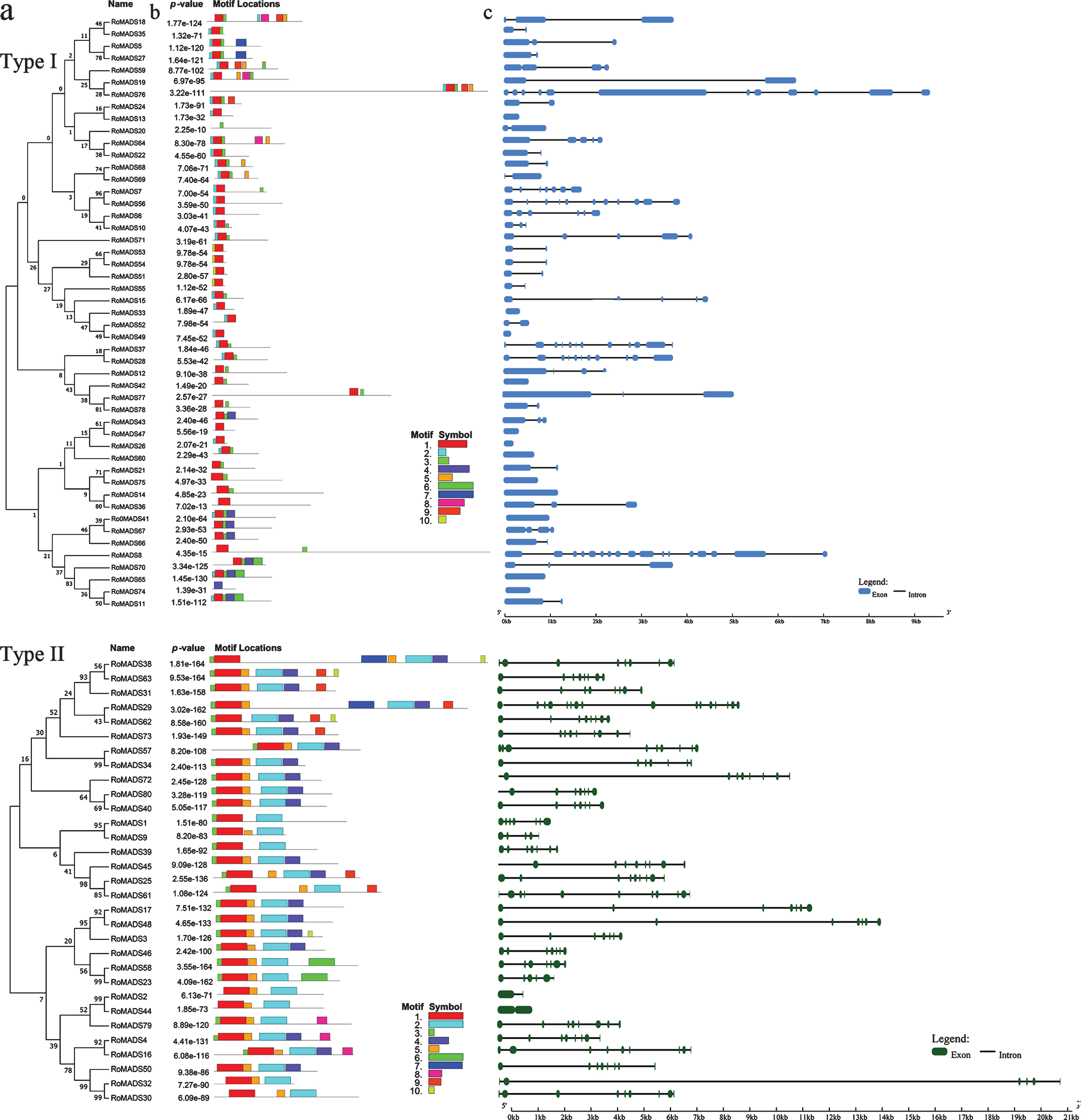
Phylogenetic relationship (a), conserved motif (b) and exon-intron structure (c) analyses of RoMADS-box proteins. Motifs 1 to 10 are indicated by differently colored boxes. The black lines represent nonconserved sequences, and 10 conserved motifs are indicated by different colors with numbered boxes. Exons and introns are represented by boxes and lines, respectively.
To better understand the structural components of RoMADS-box genes, various exon and intron structures were obtained through comparison with the corresponding genomic DNA sequences. We obtained the structure of each gene by comparing its open reading frame with the corresponding genomic sequence. In general, the closest genes exhibited similar exon/intron structures in terms of exon/intron number (Fig. 3c). With the exception of RoMADS2 and RoMADS44, both of which contained one intron and two exons, other type II RoMADSs contained more than two exons (Fig. 3c). The average number of exons in type II RoMADSs was 7.7, which was significantly higher than the average number of exons in type I genes (3.5), and this finding indicated that type II MADS-box genes might be more complex.
The promoter of a gene is an important factor in determining the initiation of gene transcription, and its sequence contains some basic elements that can reflect the potential function of the gene. To further explore the function of RoMADS-box genes, the promoter elements upstream of these genes were predicted and analyzed using PlantCARE software. The results showed that all of the cis elements contained in the MADS-box transcription factor family could be roughly divided into eight categories (Table 3). Light-responsive elements, including parts of a light-responsive element, Box 4, the GT1 motif, the G box and ACE, were the most commonly found category. We identified several plant hormone-responsive elements, such as the TGA element, TCA element, TCTC box, P box, ABRE, TGACG motif and CGTCA motif. Interestingly, most of the RoMADS-box transcription factors contain hormone-related response elements, which can be divided into five categories based on the hormone to which they respond: auxin (IAA, TGA element), gibberellic acid (GA, P box or TATC box), salicylic acid (SA, TCA element), abscisic acid (ABA, ABRE), and methyl jasmonate (MeJA, TGACG or CGTCA motif). Among these, IAA, GA and SA are factors that promote fruit growth and development, and ABA and MeJA are factors that promote fruit ripening.
Conservation of cis-acting elements in MADS-box genes in black raspberry
Conservation of cis-acting elements in MADS-box genes in black raspberry
Note: The frequency of occurrence is the ratio of genes containing cis-acting elements to the total number of RoMADS-box genes.
To explore the potential molecular mechanisms of RoMADS-box proteins, we constructed a PPI network using protein expression patterns in the STRING database. The interaction analysis revealed that directly linked genes might exhibit a tight functional connection (Fig. 4). In the network, thicker connecting line indicated a stronger interaction between the two corresponding proteins. Markov clustering revealed three distinct groups, and the largest group was associated with MADS-box proteins (part of the red) and exhibited strong interactions (AGL6, 8, 15, 16, 20, 24, SVP and AP1) corresponding to predicted RoMADS-box orthologs (RoMADS73, 38, 79, 48, 72, 16, 4 and 31, respectively) (Fig. 4). The SEP3, STK, TT16, AGL4, and SEP4 proteins (green) showed strong interactions with the predicted orthologs RoMADS63, 39, 61, 29 and 31, respectively. These specific associations suggest that these proteins jointly contribute to a shared cis or trans function in plants. The AGL12 protein exhibited a strong interaction with AGL21. Accordingly, the RoMADS46 and RoMADS3 genes might have similar functions.
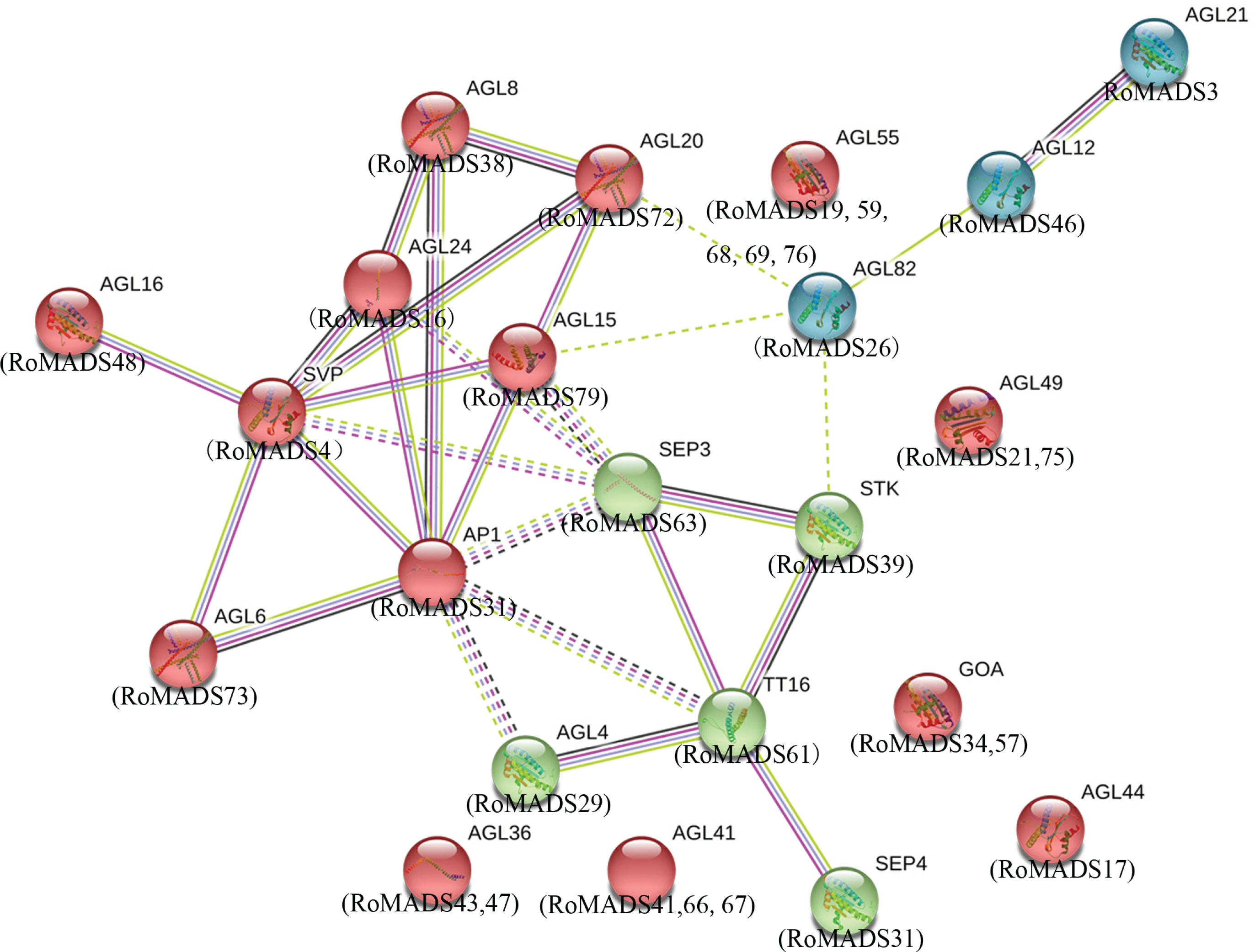
Protein-protein interactions between RoMADS-box transcription factors based on predicted AtMADS orthologs.
We investigated the spatial expression of MADS-box genes in black raspberry leaf, stem, and apex tissues and in the fruit at different stages of development. As shown in Fig. 5, 20 RoMADS-box genes (some of these 20 genes had tandem or diffuse repeats) were selected for analysis of their expression patterns in different tissues of black raspberry by qRT-PCR. These RoMADS-box genes showed different expression patterns in different tissues. The majority of the RoMADS-box genes showed higher expression in fruit than in other tissues, which was consistent with the original identification of MADS-box genes as genes that regulate fruit ripening. In particular, the RoMADS35 and RoMADS58 genes showed extremely high expression in purple fruit and might be associated with the regulation of fruit development and ripening. An expression analysis revealed that the expression of RoMADS9, RoMADS32, and RoMADS59 decreased during fruit ripening, which indicated that these genes might be negatively involved in fruit development or anthocyanin accumulation.

Expression analysis of RoMADS-box genes. The expression patterns of 20 selected RoMADS-box genes in different tissues of black raspberry were determined by qRT-PCR. Mean expression values were calculated from three independent replicates. The vertical bars indicate standard deviations.
The MADS-box family consists of important transcription factors uniquely found in plants that play an important regulatory role in many aspects of plant growth and development. Different species contain different MADS-box family members with specific functional differences. MADS-box transcription factors underwent many gene duplication events during the evolution of plants (particularly angiosperms) and form a multigene family [38]. Gene replication events are thought to be an important source of biological evolution. The presence of two or more homologous genes on a single chromosome is defined as a tandem repeat event, whereas the distribution of homologous genes on different chromosomes is known as a diffuse repeat event. Herein, we identified repeat events among 11 homologous gene pairs (22 RoMADS-box genes). Among these events, five were tandem repeat events, and six were diffuse repeat events, which indicated that the expression of RoMADS-box gene family members might be affected by homologous genes. In addition, clusters of multiple copies of some genes on chromosomes might facilitate the rapid synthesis of a large number of transcription products. Some of these clusters confer a certain selective advantage in the evolutionary process. However, the distribution of gene clusters on chromosomes is irregular, and relevant studies have shown that the exact locations and directions of members in gene clusters are not well conserved [39]. Through phylogenetic tree and conserved motif analyses, we found that the RoMADS-box genes clustered in the same subgroup harbored an essentially conserved motif, which indicated that members of the same subgroup might exhibit functional similarity. In general, a large number of motifs with different structures and functions are found in the RoMADS-box gene family, which indicates that MADS-box genes play a variety of important roles in the regulatory network of black raspberry genes. To further understand the structural diversity among black raspberry MADS-box genes, we analyzed the distribution of introns and exons in the RoMADS-box genes. With the exception of the type II genes that contain fewer introns (RoMADS2 and RoMADS44), other type II genes contain more introns, which is consistent with the general rule [17–20, 31].
Studies have found that the genomes of gymnosperms are generally larger than those of angiosperms, but gymnosperms contain fewer MADS-box gene family members than angiosperms, which might be due to the fact that the MADS-box gene family mainly functions in the growth and development of flower organs because gymnosperms have no obvious flowers [32, 40]. Gymnosperms have fewer flowering organs than angiosperms, and thus, gymnosperms have fewer MADS-box genes. In general, functional studies of the MADS-box gene family in gymnosperms are less comprehensive than those in angiosperms.
Further study of the expression levels of RoMADS-box genes in different black raspberry tissues indicated that the majority of RoMADS-box genes were more highly expressed in fruit than in other tissues, which is consistent with the original identification of MADS-box genes as genes that regulate fruit ripening [41, 42]. In particular, the RoMADS35 and RoMADS58 genes exhibited extremely high expression in purple fruit. Vrebalov et al. [43] found that the mutation of one MADS-box gene in tomato resulted in fruit abortion. Liu et al. [41] obtained one MADS-box gene encoding a transcription factor from banana that plays an important role in the regulation of the fruit ripening process during climacteric respiration. Seymour et al. [42] silenced the FaMADS9 gene in strawberry and found that this gene is an upstream regulator of strawberry fruit ripening and is involved in the simultaneous ripening of fruit during both climacteric and nonclimacteric respiration. Thus, the specific regulatory role of the RoMADS-box genes examined in this study on the development of black raspberry fruit remains to be further studied. The analysis of transcription factor promoter elements provides important reference information for predicting the potential function of genes and an important theoretical basis for determining the mechanism of action of genes. Most RoMADS-box transcription factor promoter elements were found to contain hormone-responsive elements, which indicates that members of the MADS-box family might play different regulatory roles in fruit development and maturation processes by responding to different hormone signals. The ethylene-free homeopathic elements in the RoMADS-box promoter might be due to the nonclimacteric respiration of black raspberry. Ethylene might not participate or play a major role in the ripening process, but ABA might initiate and participate in the black raspberry fruit ripening process. Previous studies have shown that GA, IAA and cytokinin can promote fruit growth and development but delay fruit color transformation and ripening [44]. Furthermore, ethylene, MeJA, and ABA can promote fruit color and ripeness [45]. The inhibition or promotion of fruit ripening is mainly determined by the interaction between endogenous and exogenous hormones in fruit, and this effect is ultimately the result of the spatiotemporal expression of endogenous genes in fruit [46].
Through an analysis of the promoter elements of members of the MADS-box family, the possible functions of different members of the family were predicted. The spatiotemporal expression patterns of members of the MADS-box family during black raspberry fruit development and ripening were systematically analyzed, and the possible roles of members of different groups within this family in fruit development were suggested or proposed. This study lays an important foundation for obtaining a more in-depth understanding of the regulatory functions of MADS-box genes in fruit development and ripening.
Footnotes
Acknowledgments
This research was supported by the Jiangsu Agricultural Science and Technology Innovation Fund (No. CX(19)2013) and the National Natural Science Foundation of China (No. 31600553).
Conflict of interest
The authors have no conflicts of interest to report.