
Abstract
Background:
Altered cellular vesicle trafficking has been linked to the pathogenesis of Huntington’s disease (HD), a fatal, inherited neurodegenerative disorder caused by mutation of the huntingtin (HTT) protein. The Rab GTPase family of proteins plays a key role in regulation of vesicle trafficking, with distinct Rabs helping specify membrane identity and mediating cellular processes including budding, motility and tethering of vesicles to their targets. In recent years several Rab GTPases—notably, Rab5 and Rab11—have been linked to the pathogenesis of neurodegenerative disorders, including HD.
Objective:
We investigated whether Rab8, which regulates post-Golgi vesicle trafficking, is able to improve HD-relevant phenotypes in a well-characterised model.
Methods:
We overexpressed Rab8 in a Drosophila model of HD testing cellular, behavioural, and molecular phenotypes.
Results:
We found that Rab8 overexpression ameliorated several disease-related phenotypes in fruit flies expressing a mutant HTT fragment throughout the nervous system, including neurodegeneration of photoreceptor neurons, reduced eclosion of the adult fly from the pupal case and shortened lifespan. Rab8 overexpression also normalised aberrant circadian locomotor behaviour in flies expressing mutant HTT in a specific population of neurons that regulate the circadian clock. Intriguingly, expression of Rab8 increased the accumulation of SDS-insoluble aggregated species of mutant HTT.
Conclusion:
Collectively, our findings demonstrate that increased Rab8 levels protect against mutant HTT toxicity and potentiate its aggregation, likely reducing the accumulation of downstream toxic soluble species.
INTRODUCTION
Huntington’s disease (HD) is a fatal autosomal dominant neurodegenerative disorder caused by a CAG trinucleotide repeat expansion in the first exon of the huntingtin gene (HTT) [1], with a prevalence of ∼11–14 cases per 100,000 in Western populations [2]. HD individuals exhibit cognitive decline, psychiatric disturbances, and motor impairment, which are associated with dysfunction and progressive loss of striatal and cortical neurons within the brain [2]. The causative CAG repeat leads to the extension of a polyglutamine (polyQ) tract within the HTT protein, causing its misfolding and the subsequent formation of several aggregated species [3]. Although the exact pathogenic mechanisms remain elusive, mutant HTT toxicity is predominantly attributed to a toxic gain of function that disrupts several cellular processes, including vesicular trafficking [4].
Cellular vesicle trafficking is regulated by the Rab (Ras-related proteins in brain) family of small GTPases, which in mammals constitutes ∼70 members [5]. Rab GTPases localize to distinct cellular compartments where they bind to target membranes and recruit distinct effector proteins, helping specify membrane identity and mediate cellular processes including budding, motility, and tethering of vesicles to their targets [5]. The activity and localisation of Rab GTPases is dependent upon a suite of specific accessory proteins, including guanine exchange factors (GEFs) and GTPase activating proteins (GAPs) that catalyse cycling between active GTP bound and inactive GDP bound forms. Importantly, alterations in Rab function underlie or contribute to the pathogenesis of several diseases [6, 7], including the neurodegenerative disorders Alzheimer’s disease, Parkinson’s disease (PD), HD, and amyotrophic lateral sclerosis [8–10]. Notably, a familial form of PD is caused by loss of function mutations in RAB39B [11–13], while RAB29 is likely a risk locus for sporadic PD [14, 15]. Furthermore, the parkinsonian proteins LRRK2 and PINK1 phosphorylate a subset of Rab GTPases [16–18], suggesting that disruption of these normal physiological processes contribute to the onset of PD.
Rab GTPases have also been closely linked to the pathophysiology of HD, with impairment in function, localisation or trafficking of Rab3, Rab5, Rab8, and Rab11 observed in a number of studies [19–27]. Notably, Rab8 and wild-type HTT, which is involved in Golgi regulation and vesicle trafficking [23, 28], interact with the multifunctional adapter protein optineurin (OPTN), forming a complex that helps maintain Golgi morphology and post-Golgi trafficking to the plasma membrane [29, 30] and to lysosomes [26, 31]. Indeed, in cells expressing mutant HTT the Rab8/OPTN complex becomes de-localized from the Golgi with repercussions on both the secretory and the lysosomal pathways [26].
Downregulation of Rab5 has been found to exacerbate mutant HTT toxicity in fruit flies, while overexpression of Rab5 and Rab11 is protective in fly and mammalian cell models of HD [24, 33]. The beneficial effects of Rab5 are likely due to upregulation of autophagy [24], the main cellular pathway for clearance of misfolded proteins and damaged organelles, while the overexpression of Rab11 has been found to ameliorate compromised synaptic structure and function [32, 33]. To date, the potential benefits of Rab8 overexpression are untested in HD models.
Here we explore the pathological ramifications of overexpressing Drosophila Rab8 in a well-characterised fruit fly model of HD. We find that Rab8 overexpression reduces neuronal death and associated HD phenotypes, which are likely dependent upon the nucleotide binding state of Rab8. We also demonstrate that Rab8 overexpression promotes mutant HTT aggregation, suggesting that the observed protection may be due to reduction in levels of soluble mutant HTT species.
MATERIALS AND METHODS
Drosophila stocks and husbandry
Flies were grown on maize food (72 g/L maize meal, 80 g/L glucose, 50 g/L brewer’s yeast, 8.5 g/L agar, 0.27 g/L nipagin, and 0.3% propionic acid) under a 12 h light, 12 h dark regime (LD 12:12) at 25°C. Lines y1 w*; PUASp-YFP.Rab845 (#9782), y1 w*; PUASp-YFP.Rab8.Q67L05 (#9781), y1 w*; PUASp-YFP.Rab8.T22N12 (#9780), and w; P w +mW.hs = GawBelavC155 (#458) were obtained from the Bloomington Stock Centre (USA). The w; P UAS-Httex1Q934F1 (#68148) and w; P UAS-HTTex1Q20111F1L (#68412) lines were kindly provided by J. Lawrence Marsh and Leslie Thompson (University of California, Irvine, CA, USA). The yw; Pdf-GAL4 driver line was a gift from Michael Rosbash (Brandeis University, USA).
Eclosion analysis
Pan-neuronal expression of reporter genes was achieved by crossing UAS reporter females to males carrying the X-linked elav-GAL4 driver. Thus, F1 males carried but did not express the reporter. The eclosion ratio (females/females + males×100) is a measure of how reporter expression (limited to females in test genotypes) affects survival during development. A minimum of 10 vials containing reporter x driver crosses (with 5 females and 5 males each) were set up on standard maize food up at 25°C for five days. The F1 were collected, sexed and their number recorded daily.
Pseudopupil assay
Crosses were set up as above and flies were collected at day 1 and day 7 post-eclosion. Heads were glued onto a microscope slide with nail varnish and the number of rhabdomeres in 20–60 ommatidia scored per head. Analyses were carried out with an Optiphot-2 microscope (Nikon) using a Plan Apo 40×/1.0NA oil objective.
Survival analysis
Crosses were set up as above. For each genotype, 100 females were collected within 6 h of emergence, equally distributed among 10 vials, and maintained at 25°C, LD 2:12. The number of surviving flies were recorded daily. Flies were transferred to new vials every other day.
Locomotor activity
All activity experiments were performed at 25°C using DAM2 activity monitors (TriKinetics, USA). Newly eclosed males were individually placed in glass activity tubes (80 mm long, 5 mm in diameter, 1 mm thick) on sugar food (46 g/L sucrose, 46 g/L yeast, 10 g/L agar, 2 g/L nipagin). Flies were entrained under LD 12:12 for 3 days and then maintained under constant darkness (DD) for one week. Periodicity and period length were assessed with autocorrelation and CLEAN spectral analysis on 30 min bins intervals, using the MAZ software [34].
Filter retardation assay
Proteins were extracted from 100 fly heads/genotype in 100μl of PBS supplemented with a protease inhibitor cocktail (Roche) in 1.5 ml microcentrifuge tubes. After grinding the tissue with a plastic pestle, samples were sonicated in 5 cycles of 30 s ON/30 s OFF with a Bioruptor® (Dia-genode). Samples were cleared by centrifugation at 200×g for 5 min at 4°C. Total protein concentration was determined by Bradford assay relative to a bovine serum albumin (BSA) standard. Then 100μg of protein extract was suspended in 150μl of 2% SDS in PBS. Samples were boiled for 5 min and then vacuum filtered through a cellulose acetate membrane (0.45μm, Whatman) equilibrated in 2% SDS using a Hybri Slot 24 dot blotting apparatus (Core Life Science). Membranes were washed 3 times with 150μl of 2% SDS in PBS and subsequently air-dried prior to blocking (in 5% skimmed milk, 20 mM Tris, 150 mM NaCl, 0.1% tween-20) and incubation with α-HTT antibodies. Image analysis was performed with ImageJ (https://imagej.nih.gov/ij/). The α-HTT signal in elav > HTT93Q, Rab8 flies was normalized to the α-HTT signal in elav > HTT93Q individuals.
AGERA assay
The AGERA protocol [35] was adapted for use with fly samples. Briefly, proteins were extracted from 100 fly heads as described for the filter retardation assay. Protein extracts were normalised to OD595 = 0.5 using Bradford reagent (Sigma). For each sample, 30μl were diluted 1:1 with a non-reducing Laemmli buffer (50 mM Tris–HCl pH 6.8, 33% glycerol, 1.2% SDS, 0.001% bromophenol blue) and incubated at 95°C for 5 min. Samples were loaded onto an agarose gel (1.5% agarose, 375 mM Tris-HCl, pH8.8, 0.1% SDS) and run in Tris-Gly buffer (192 mM glycine, 25 mM Tris-base, 0.1% SDS) at 20 V O/N. Gels were blotted onto nitrocellulose membrane with pore size of 0.2μm (Amersham) using a semi-dry blotter (BioRad) and transfer buffer (25 mM Tris, 192 mM glycine, 0.1% SDS, 15% methanol). Membranes were incubated with α-HTT antibodies. Image analysis was performed with ImageJ. The α-HTT signal in elav > HTT93Q, Rab8 flies was normalized to the α-HTT signal in elav > HTT93Q individuals.
Antibodies
α-HTT (MAB5374, Millipore, mouse monoclonal, 1:2,000); α-mouse HRP (PI-2000, Vector, 1:10,000).
Statistical analyses
All statistical analyses were performed using GraphPad Prism software V7 or V8.
RESULTS
Overexpression of Rab8 ameliorates mutant HTT-induced eclosion defects and shortened lifespan
We employed the GAL4/UAS system [36] to test whether overexpression of Drosophila Rab8 (Rab8) modulates mutant HTT-induced toxicity in vivo. We used an X-linked elav-GAL4 driver to express a mutant HTT exon 1 fragment containing a stretch of 93 glutamines (HTT93Q) in all neuronal cells [37]. In agreement with published findings, this resulted in toxicity as exemplified by several phenotypes, including reduced eclosion of adult flies from the pupal case, shortened lifespan and degeneration of photoreceptors [38].
We first tested the effect of overexpressing Rab8 on the reduced eclosion of HTT93Q flies. This parameter reports the eclosion of females expressing HTT93Q compared to non-expressing male siblings (see Materials and Methods). Eclosion was significantly lower for elav > HTT93Q flies compared to parental controls (UAS-HTT93Q and elav-GAL4) (ANOVA, F8,81 = 11.76, p < 0.0001. Tukey’s multiple comparisons, elav > HTT93 vs. UAS-HTT93Q, p < 0.0001; elav > HTT93 vs. elav-GAL4, p < 0.0001. Figure 1A). We found that co-expressing HTT93Q and Rab8 (elav > HTT93Q, Rab8) increased eclosion but just missed statistical significance (Tukey’s multiple comparisons, elav > HTT93 vs. elav > HTT93Q, Rab8 p = 0.0537; Fig. 1A). To investigate this further, and determine whether Rab8 enzymatic activity was required, we assayed flies expressing HTT93Q together with constitutively active (Rab8-CA) or the dominant negative (Rab8-DN) forms of Rab8 [39]. We found a highly significant improvement in the eclosion of flies expressing both Rab8-CA and HTT93Q versus HTT93Q alone (Tukey’s multiple comparisons, elav > HTT93 vs. elav > HTT93Q, Rab8-CA, p = 0.0058; Fig. 1A). Conversely, Rab8-DN did not alter the HTT93Q-mediated eclosion defect (Tukey’s multiple comparisons, elav > HTT93Q vs elav > HTT93Q, Rab8-DN, P > 0.9999) (Fig. 1A). These results suggest that the enzymatic activity of Rab8 is required to ameliorate the eclosion defect observed in HTT93Q expressing flies. Moreover, such specific effects argue against limiting levels of GAL4 (‘titration’) as the cause of higher eclosion in elav > HTT93Q, dRAB8-CA compared to elav > HTT93Q flies. Western blot data support this conclusion (Supplementary Figure 1). Finally, to test the specificity of the interaction between HTT93Q and Rab8 we expressed a HTT exon1 construct containing a non-pathogenic polyglutamine tract (HTT20Q) either alone (elav > HTT20Q) or in combination with Rab8 (elav > HTT20Q, Rab8), and found no difference in the rate of eclosion (Supplementary Figure 2; Supplementary Table 1).
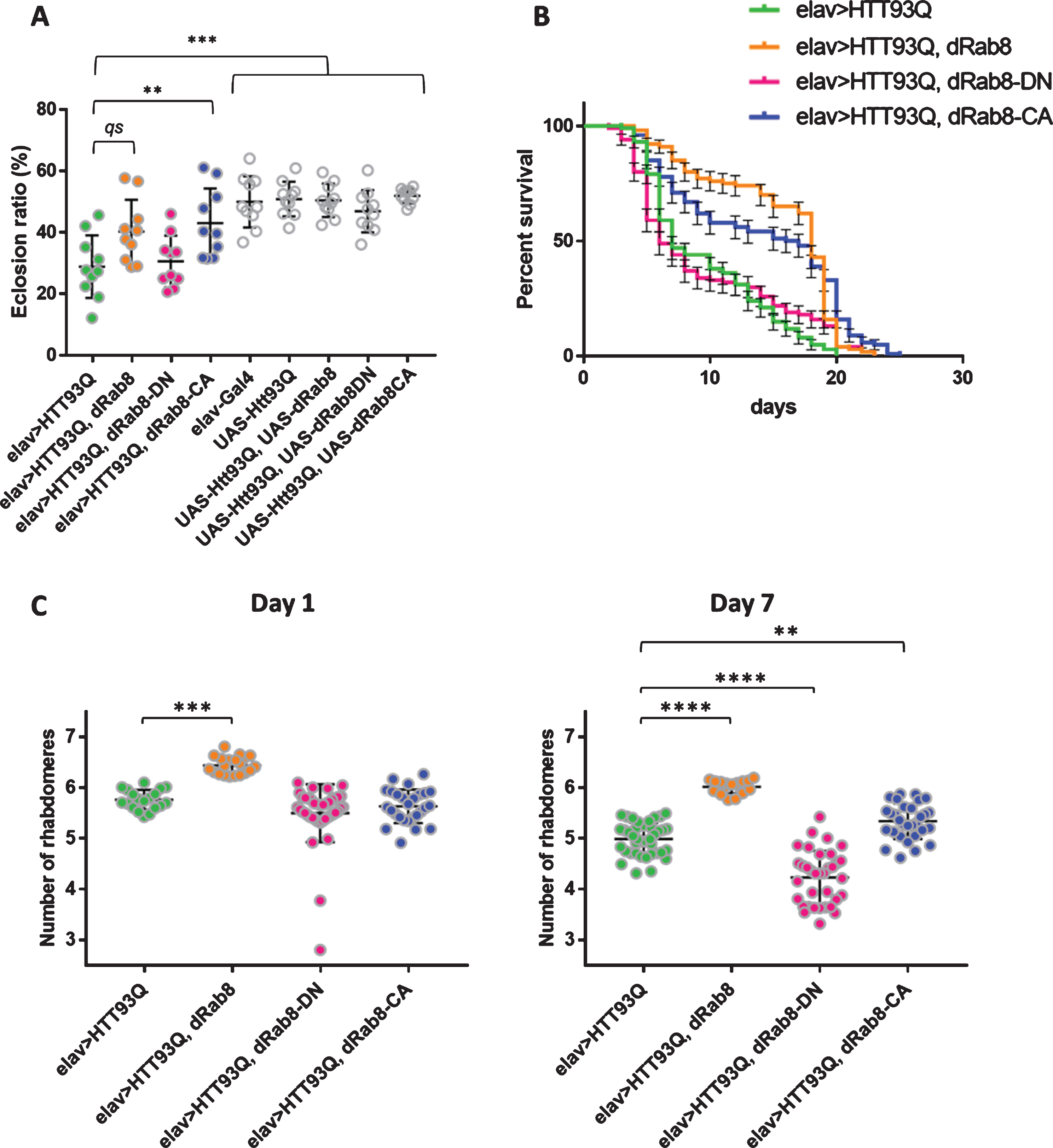
Overexpression of Rab8 ameliorates mutant HTT-induced phenotypes in a D. melanogaster model of HD. A) Eclosion. The eclosion ratio (females/females + males×100) is a measure of how reporter(s) expression (limited to females in test genotypes) affects survival during development. The different genotypes (full circle = ‘test’, open circle = control) were compared by one-way ANOVA (F8,81 = 11.76, p < 0.0001) and then multiple comparisons were carried out according to Tukey’ s test. qs = quasi significant = p=0.0537. **p < 0.01. ***p≤0.001. ‘Test’ genotypes: elav > HTT93Q (green), elav > HTT93Q, Rab8 (orange), elav > HTT93Q, Rab8-DN (magenta) and elav > HTT93Q, Rab8-CA (blue). Each point represents an independent replicate, mean±SD are shown. See Table S1 for descriptive statistics. B) Survival curves. Survival (%) versus days of 100 female flies (10 flies×10 replicates) under identical conditions. Genotypes : elav > HTT93Q (green), elav > HTT93Q, Rab8 (orange), elav > HTT93Q, Rab8-DN (magenta) and elav > HTT93Q, Rab8-CA (blue). The median survival was, 7, 18, 6 and 16.5 days, respectively. The comparison by Mantel-Cox log-rank test (χ2 = 70.24, df = 3, p < 0.0001) reported a significant difference. Error bars correspond to SE. C) Pseudopupil assay. Average number of rhabdomeres per fly at day 1 and day 7 post-eclosion. Genotypes: elav > HTT93Q (green), elav > HTT93Q, Rab8 (orange), elav > HTT93Q, Rab8-DN (magenta) and elav > HTT93Q, Rab8-CA (blue). Day 1. Comparisons were carried out by Kruskal-Wallis test (p < 0.0001) followed by Dunn’s multiple comparisons. ***p < 0.001. Day 7. Comparisons were carried out by ANOVA (F3,117 = 95.3, p < 0.0001) followed by Tukey’s multiple comparisons. **p < 0.01. ****p < 0.0001. Each point represents a fly, mean±SD are shown. See Supplementary Table 2 for descriptive statistics.
We next measured the lifespan of adult flies expressing HTT93Q on its own or in combination with Rab8, Rab8-DN or Rab8-CA (Fig. 1B). Pan-neuronal HTT93Q expression (elav > HTT93Q) resulted in early lethality (Fig. 1B; Supplementary Figure 3). Co-expression with Rab8-DN (elav > HTT93Q, Rab8-DN) did not significantly improve lifespan whereas co-expression with Rab8 (elav > HTT93Q, Rab8) or Rab8-CA (elav > HTT93Q, Rab8-CA) increased survival (Mantel-Cox Log-rank test, χ2 = 70.24, df = 3, p < 0001). A stronger effect was obtained when co-expressing wild-type Rab8, which extended the median lifespan by 11 days compared to flies expressing HTT93Q alone (Fig. 1B). Thus, Rab8 overexpression improved the longevity of HD flies, with the strongest effect observed when the protein retained its ability to cycle between GDP- and GTP-bound forms.
Rab8 reduces HTT93Q-induced neurodegeneration in the fly eye
We next examined the neuroprotective potential of Rab8 overexpression using the pseudopupil assay, which assesses the number of visible photoreceptor neurons (through their rhabdomeres) per ommatidium of the compound eye. We scored the number of rhabdomeres at days 1 and 7 post-eclosion in females expressing HTT93Q (elav > HTT93Q) alone or co-expressing one of the Rab8 forms (elav > HTT93Q, Rab8; elav > HTT93Q, Rab8-CA; elav > HTT93Q, Rab8-DN). For day 1 we adopted non-parametric statistics, due to one genotype failing the assumption of normality (elav > HTT93Q, Rab8-DN; D’Agostino and Pearson normality test, p < 0.0001). As expected, we observed a reduction in the number of rhabdomeres in elav > HTT93Q flies (Fig. 1C; Supplementary Table 2). The effect was mitigated by co-expressing HTT93Q together with Rab8, but not with the constitutively active or dominant negative forms of the protein (Kruskal-Wallis, p < 0.0001. Dunn’s multiple comparisons test, elav > HTT93Q vs. elav > HTT93Q, Rab8 p < 0.001; elav > HTT93Q vs. elav > HTT93Q, Rab8-CA P > 0.999; elav > HTT93Q vs. elav > HTT93Q, Rab8-DN p = 0.303; Fig. 1C; Supplementary Table 2). Further reductions in the number of rhabdomeres were observed for all genotypes at day 7 (Fig. 1C; Supplementary Table 2). Again, we observed a protective effect when co-expressing the wild type form of Rab8 (ANOVA, F3,117 = 95.3, p < 0.0001. Tukey’s multiple comparisons, elav > HTT93Q vs. elav > HTT93Q, Rab8 p < 0.0001). However, a protective role also emerged for Rab8-CA (Tukey’s multiple comparisons, elav > HTT93Q vs. elav > HTT93Q, Rab8-CA p < 0.01), whereas co-expression of Rab8-DN resulted in a significant worsening of the phenotype (Tukey’s multiple comparisons, elav > HTT93Q vs. elav > HTT93Q, Rab8-DN p < 0.0001) (Fig. 1C; Supplementary Table 2). Interestingly, pan-neuronal expression of Rab8-DN only (elav > Rab8-DN) did not result in any reduction in rhabdomeres numbers (Supplementary Table 2), suggesting a synergistic interaction between mutant HTT and Rab8 inactivation.
Rab8 improves abnormal locomotor activity rhythms due to HTT93Q expression
In addition to the motor, cognitive and psychiatric disturbances found in HD individuals, sleep and circadian problems have been described [40]. It has been previously shown that expression of mutant HTT in fruit flies under the control of a specific circadian driver compromises rhythmic locomotor activity [41, 42]. The circadian clock is a complex regulatory mechanism based upon cycling gene expression in several brain neurons, resulting in their coordinated interplay [43]. The role of the lateral neurons ventral small (s-LNv) and large (l-LNv) in maintaining circadian rhythms is particularly well described. These neurons produce the neuropeptide Pigment Dispersing Factor (PDF) that is required for sustained rhythmic locomotor activity under constant environmental conditions. In fact, flies that do not produce PDF, or whose PDF-producing neurons have been ablated, are arrhythmic or cycle with a shorter period [44]. Notably, the expression of mutant HTT in PDF-neurons results in compromised self-sustained rhythmicity [41, 42]. Thus, we tested whether Rab8 could exert a neuroprotective role in PDF-neurons. Using a Pdf-GAL4 driver we expressed HTT93Q and assessed the rhythmicity of Pdf > HTT93Q males after a week in constant darkness (DD) (Fig. 2A; Supplementary Table 3). About 80% of flies expressing HTT93Q (Pdf > HTT93Q) were arrhythmic, and the period of those rhythmic was more than 1 h shorter than controls (Fig. 2A–C; Supplementary Table 3). The concomitant overexpression of Rab8 (Pdf > HTT93Q, Rab8) ameliorated these phenotypes, decreasing the percentage of arrhythmic flies to about 30% (Pdf > HTT93Q vs. Pdf > HTT93Q, Rab8χ2 = 51.65, df = 1, p < 0001) and restoring the period to wild-type values (Kruskal-Wallis, p < 0.0001. Dunn’s multiple comparisons test Pdf > HTT93Q vs. Pdf > HTT93Q, Rab8 P < 0.0001; Pdf > HTT93Q, Rab8 vs. Pdf-GAL4 p = 0.176; Fig. 2A–C, Supplementary Table 3).

Rab8 ameliorates locomotor activity rhythms disturbed by mutant HTT. A) Average locomotor activity profiles. Activity levels (number of crossing/30 min) are shown on the Y-axis; time (96 intervals of 30 min) is on the X-axis. Data are double plotted (i.e., day1-day2, day2-day3, etc.). Flies were monitored for 2 days under light-dark (LD) and for 7 days under constant darkness (DD) at 25°C. Light is shown in yellow, darkness in blue highlight. To aid the visual appreciation of rhythmicity, activity levels are not to scale and standard deviation is omitted. Both rhythmic and arrhythmic individuals contributed to the graphs, number of flies as in B (see also Supplementary Table 3). B) Percentage of rhythmic flies across genotypes. Only about 20% of Pdf-GAL4 > HTT93Q flies resulted rhythmic while the percentage raised to almost 70% in Pdf-GAL4 > HTT93Q, Rab8 (χ2 = 51.65, df = 1, ****p < 0001). Number of flies are in brackets. C) Period of locomotor activity rhythms. Each data point represents the period of a fly, only individuals with simple (i.e., single) rhythmicity are considered. Number of flies are given in parentheses. The average period (±SD) in Pdf-GAL4 > HTT93Q was 22.63±1.83 whereas in Pdf-GAL4 > HTT93Q, Rab8 flies it raised to 24.88±1.58. The latter was not significantly different from the value of 24.28±0.52 recorded for Pdf-GAL4 controls. Not all data were normally distributed, hence non-parametric statistic were applied. Kruskal-Wallis, p < 0.0001. Dunn’s multiple comparisons test Pdf > HTT93Q vs. Pdf > HTT93Q, Rab8****p < 0.0001. Pdf > HTT93Q vs. Pdf-GAL4 p < 0.0001. Pdf > HTT93Q vs. UAS HTT93Q p = 0.0001. Pdf > HTT93Q vs. UAS HTT93Q, UAS-Rab8 p = 0.160. Pdf > HTT93Q, Rab8 vs. Pdf-GAL4 p = 0.176. Pdf > HTT93Q, Rab8 vs. UAS HTT93Q p = 0.004. Pdf > HTT93Q, Rab8 vs. UAS-HTT93Q, UAS-Rab8 p < 0.0001. Pdf-GAL4 vs. UAS HTT93Q p > 0.999. Pdf-GAL4 vs. UAS HTT93Q, UAS-Rab8 p < 0.001. UAS HTT93Q vs. UAS HTT93Q, UAS-Rab8 p = 0.94.
Overexpression of Rab8 promotes aggregation of mutant HTT in fruit flies
Overexpression of the Rab GTPase Rab5 has previously been found to decrease levels of aggregated mutant HTT by promoting macroautophagy [24]. Thus, we employed both filter retardation and AGERA (Agarose Gel Electrophoresis for Resolving Aggregates) assays to assess whether Rab8 overexpression affected the aggregation of HTT93Q in flies. The filter-retardation assay measures the retention of insoluble, SDS-resistant aggregates [45] while AGERA detects all aggregated species, including soluble oligomers [35]. Surprisingly, both assays revealed more mutant HTT aggregates in flies co-expressing HTT93Q and Rab8 (elav > HTT93Q, Rab8) compared to individuals expressing HTT93Q only (elav > HTT93Q) (Fig. 3A, B). Notably, AGERA revealed that elav > HTT93Q, Rab8 flies had larger mutant HTT-containing species (Fig. 3B, red arrowheads). These data suggest that the neuroprotection conferred by Rab8 is due to the promotion of mutant HTT into larger aggregates, thereby diminishing levels of toxic soluble mutant HTT species.
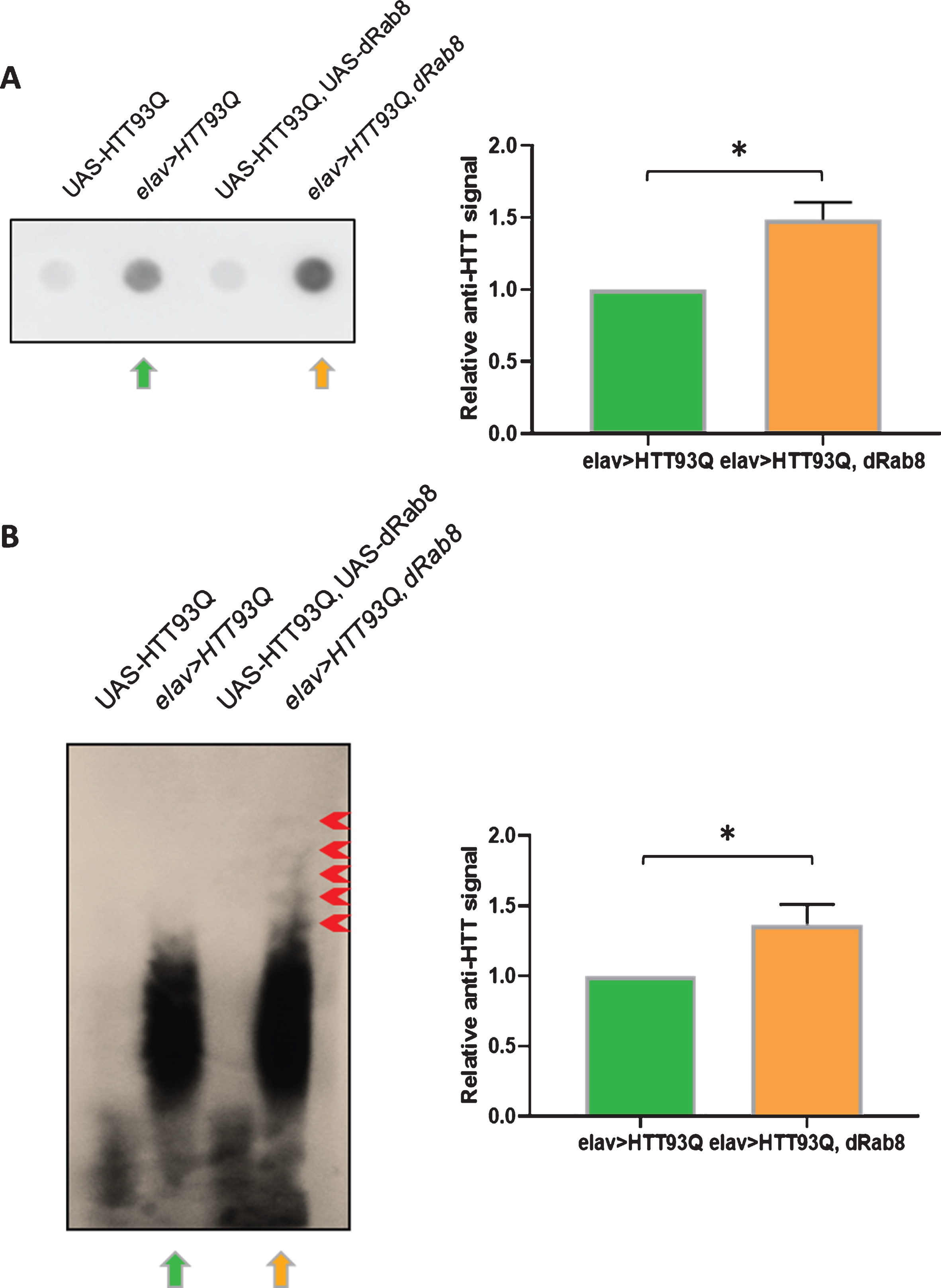
Co-expression of HTT93Q and Rab8 increases mutant HTT aggregation in vivo. A) Filter retardation assay. On the left, representative dot-blot (of three) showing HTT immuno-signal from head protein extracts. Green and orange arrows indicate genotypes elav > HT93Q and elav > HTT93Q, Rab8, respectively. Control flies not expressing HTT93Q are: UAS-HTT93Q, UAS-Rab8 and UAS-HTT93Q. B) AGERA. On the left, representative blot (of three) showing HTT immuno-signal from head protein extracts after electrophoresis. Green and orange arrows indicate genotypes elav > HTT93Q and elav > HT93Q, Rab8, respectively. Control flies not expressing HTT93Q are: UAS-HTT93Q, UAS-Rab8 and UAS-HTT93Q.
DISCUSSION
Rab8 localizes to the Golgi and the recycling endosomes and is a major regulator of vesicle delivery from the trans-Golgi network to the plasma membrane [46–50]. It is involved in exocytosis in several cell types, including polarized epithelial cells [47, 51], photoreceptor neurons [52–54] and neuronal dendrites [55, 56]. In neurons it has been implicated in the transport of several receptors, including the α-amino-3-hydroxy-5-methyl-4- isoxazolepropionic acid (AMPA) receptor [57–59], α2B- and β2- adrenergic receptors [60] and the metabotropic glutamate receptor 1 (mGluR1) [61]. In addition, Rab8 participates in autolysosome formation, the last step of autophagosome maturation [62–66].
Rab8 has previously been linked to wild-type HTT via interactions with the multifunctional adapter protein OPTN, which plays a key role in Golgi trafficking and morphology [23, 28–31]. Here we have found that Rab8 overexpression in mutant HTT expressing flies reduced neurodegeneration and improved a number of disease-relevant phenotypes, including shortened lifespan and behavioural alterations. Interestingly, we find that a dominant negative form of Rab8 (Rab8-DN) does not rescue these phenotypes, whilst a constitutively active form (Rab8-CA) does confer rescue, albeit to a lesser degree than wild-type Rab8. The Rab8-DN construct has a mutant GTP-binding domain such that GTP binding is defective [39], and thus it is likely that the GTP-bound active form of Rab8 is required for the protection observed. Interestingly, Rab8-CA—in which GTPase activity is defective, and thus GTP is constitutively bound—is not as efficacious as wild-type Rab8, suggesting that cycling between GDP- and GTP-bound forms of Rab8 is critical for achieving more robust rescue. We find that while Rab8-CA strongly rescues the mutant HTT eclosion defect in comparison to wild-type Rab8, its effect on survival is more modest. This may be due to GDP/GTP exchange being more critical in adult flies than during development. Indeed, we also find this to be the case when considering rhabdomere degeneration, where Rab8-CA is much less protective than wild-type Rab8.
Rab8 appears to promote aggregation of mutant HTT in these animals, leading to the presence of larger aggregated species that may sequester the more toxic soluble oligomeric species of mutant HTT. Notably, this model is in agreement with our previous observation that Rab8 enhances fibrillization of α-synuclein (aSyn), an aggregation-prone protein associated with PD [67]. This study found that Rab8 directly interacted with aSyn in the rodent brain and that its overexpression in mammalian cells and Drosophila ameliorated neurodegeneration and locomotor defects. These data suggest that Rab8 can potentiate aggregation of soluble misfolded aggregation-prone proteins, thereby decreasing downstream cellular toxicity, neuronal loss and consequent disease phenotypes. While the mechanism of this potentiation is not clear, it may be that increased levels of Rab8 lead to co-sequestration of Rab8 and mutant HTT in insoluble aggregates via indirect physical interactions orchestrated by OPTN. Indeed, a recent study detected Rab8a as a constituent of the insoluble mutant HTT fraction from the brains of R6/2 HD model mice, supporting this hypothesis [68]. As the GAL4-UAS system can lead to transgenes being expressed at ultra-physiological levels, it will be important to explore the effects of more modest modulation of Rab8 expression levels in the future. Futhermore, additional work will be needed to clarify whether direct interactions between Rab8 and mutant HTT are required for the modulation of mutant HTT aggregation and the downstream phenotypes observed. Notably, aSyn phosphostatus affects interactions with Rab8, suggesting that similar posttranslational modifications in mutant HTT may be relevant in the neuroprotection conferred by Rab8 [67]. Ultimately, our work supports past research suggesting that increased activity or expression of Rab8 may have therapeutic relevance in HD and other neurodegeneration disorders associated with protein misfolding.
Experiments using Drosophila
In the UK experimentation using Drosophila melanogaster is not regulated by the Animals (Scientific Procedures) Act 1986. We complied with experimentation guidelines from our institution.
CONFLICT OF INTEREST
The authors have no conflicts of interest to report.
Footnotes
ACKNOWLEDGMENTS
This study was supported by funding from the National Centre for the 3Rs to ER, FG, and CPK (NC/K001019/1) and the Medical Research Council to FG (MR/R011621/1). LD conducted the experimental analysis. The study was designed by all authors. The data was analysed by LD and ER. The manuscript was written and edited by all authors.