
Abstract
BACKGROUND:
Methylglyoxal (MG) is a highly reactive compound derived from sugar metabolism, precursor of cytotoxic products named Advanced Glycation End products (AGEs). Increased MG levels and MG-related AGEs have been long associated with diabetes and its complications. The association between MG and hypercholesterolemia – induced by high-fat diets- has not been studied previously, neither the effect of olive oil addition to this diets.
OBJECTIVE:
To analyze the variations of MG and cholesterol levels under the effect of high-fat and olive oil-enriched diets.
METHODS:
Sixteen healthy male adult (New Zealand White, 6 months old) rabbits were fed with commercial pellets (control, four rabbits), or with 14% of bovine grease added to the pellets (high-fat diet, twelve rabbits). After six months, bovine grease was reduced to 7% (half fat diet, eight rabbits). Then, 7% of olive oil was added to the pellets (protective diet, four rabbits). Glycaemia, cholesterolemia, HDL-cholesterolemia, triglyceridemia, fructosamine and MG-H1 levels were assessed in all experimental conditions (n = 4).
RESULTS:
The high-fat diet promoted a hypercholesterolemia associated with increased MG-H1 levels (p < 0.05) in non-diabetic and non-obese rabbits. Replacement of the high-fat diet with olive oil decreased significantly the cholesterolemia and MG-H1 levels (p < 0.05).
CONCLUSIONS:
Our data indicates that the replacement of a high-fat diet with olive oil can recover normal blood cholesterol values and can decrease MG-H1 levels.
Introduction
Methylglyoxal (MG) is a physiological reactive dicarbonyl compound mainly produced during the glycolisis from glucose and fructose [1–3]. MG is a potent glycating agent, which reacts with DNA, lipids, and proteins producing Advanced Glycation End products (AGEs) [4]. Glycation of proteins by methylglyoxal is directed toward arginine residues, forming mainly the hydroimidazolone MG-H1—a quantitatively and functionally important AGE product in physiological systems [5]. The effects of MG on diabetes and hypertension have been long recognized [6, 7]. Recent studies have also shown that MG and AGEs may contribute to the obesity related to diabetes [8, 9].
Furthermore, increasing evidence points to exogenous AGEs (derived mostly from food and tobacco) as minor contributors to the body’s AGE pool, where they become indistinguishable from endogenous AGEs, both in structure and function [10]. In some cases, glycotoxins (AGEs) derive from diet (Reviewed in [11]), but in others they are a consequence of diabetic pathology [12, 13]. However, the relationship between high-fat diets (enriched in saturated lipids) and MG is poorly understood.
On the other hand, several compounds have been developed to scavenge MG and inhibit AGEs formation (Reviewed in [14]), but we are still far from having an effective strategy to prevent MG-induced mechanisms.
The animal model used in this work, HyperCholesterolemic Adult Rabbits fed with high-fat Diet enriched in saturated lipids (HCARDA), display high risk to acquire vascular diseases [15] and infertility [16–18] without hyperglycemia. Interestingly, we found that virgin olive oil (enriched in unsaturated lipids) added to the high-fat diet recovers the serum cholesterol level and fertility parameters [17].
Based on the results, we hypothesize that high-fat diet increase MG-H1 levels, meanwhile, the replacement of the fat with olive oil in this diet decrease MG-H1 levels. In this work, we analyzed several biochemical markers under diets enriched with saturated and unsaturated lipids to establish a comparative vision between MG-H1 and cholesterol levels in rabbits.
Materials and methods
Ethics statement
Animal studies described here were reviewed and approved by the animal care and use committee: Institutional Committee for Use of Laboratory Animals (IACUC- http://fcm.uncuyo.edu.ar/paginas/index/cicual); protocol reference number: 06_150702.
Reagents
Unless otherwise stated, all chemicals and solvents of the highest grade available were obtained from Sigma (St. Louis, MO, USA) and Merck (Darmstadt, Germany).
First bovine juice: “Primer jugo bovino”, commercial preparation (composed by 55% saturated fat, Juan Lopez y CIA.), Argentina Alimentary Code (http://www.anmat.gov.ar/alimentos/codigoa/CAPITULO_VII.pdf; article 543 –resolution 2012, 19.10.84).
Olive oil was referred as OO and corresponds to virgin olive oil (Arauco varietals).
Rabbit model
Sixteen fertile male rabbits (White New Zealand, 6 months old) were purchased from a rabbit farm (Verde-Azul farm, Buenos Aires - Argentina). Rabbits were caged and maintained with a photoperiod of 12 hours light/day and a temperature ranging from 18–25°C. The animals were split in two groups (Fig. 1). The first group (normal serum cholesterol rabbits, designated NCR, Fig. 1) was formed by four animals fed ad libitum with a standard rabbit diet. The standard diet was composed of 17% crude protein, 16% fiber, 2% minimal ether extract (0% saturated fat), 5.3% minerals (data from the manufacturer’s analysis, GEPSA FEEDSH, Buenos Aires - Argentina). The second group (hypercholesterolemic rabbits, named HCARDA, Fig. 1, ED1) was formed by twelve animals fed ad libitum with standard diet plus 14% w/w of bovine fat (this amount of fat corresponded to an addition of 36% of calories consumed) following our previous model [16–18]. After six months, four animals in the HCARDA group were sacrificed. A new diet, consisting of half fat added to the diet, was applied to the rest of the animals (this group was called 1/2 HCARDA, Fig. 1, ED2:7% w/w bovine fat equal to 18% calories added). After six months with ED2 (total experimental time: 12 months, Fig. 1), four animals were also sacrificed and a new diet was also applied to the remaining rabbits, ED3 (ED3 consisted in ED2 plus 7% v/w olive oil equal to 36% calories added, called 1/2 HCARDA + 1/2 OO, following previous protocol [17]). The last group of animals was sacrificed six months later (total experimental time: 18 months, Fig. 1). In this sense, rabbits received consecutively different experimental diets (ED1, ED2 and ED3) during six months: 1/2 HCARDA group received ED1 and ED2, and 1/2 HCARDA + 1/2 OO group received ED1, ED2 and ED3.

Diets and experimental groups. Normal diet (ND = balanced food for rabbits) or experimental diets (ED1: ND + 14% of cow grease w/w; ED2: ND +7% of cow grease w/w; ED3: ED2 + 7% olive oil –OO- v/w) were applied to adult rabbits during 6months. The first group, fed with ND, was named NCR (normal cholesterolemic rabbits); the second group, fed with ED1, was named HCARDA (Hypercholesterolemia acquired by acutely feeding adult rabbits with standard diet supplemented with bovine fat); the group fed with ED2 was 1/2 HCARDA; and the group fed with ED3 was 1/2 HCARDA + 1/2 OO. All groups were composed for four animals (n = 4).
Blood was obtained fortnightly from marginal ear vein in all rabbit simultaneously. Immediately after bleeding, the blood was left in a microtube and centrifuged at 1100 xg by 10 minutes in a clinical centrifuge. The supernatant was carefully vacuumed and aliquots were processed using a GTlab kit (GTlab, Rosario, Argentina) to determine cholesterol; cholesterol-HDL, triglycerides, glucose, and fructosamine, following the manufacturer’s instructions.
In addition, aliquots of blood sample were anti-coagulated with EDTA (0,342 M - Wiener 1898552) and used for glycosylated hemoglobin determination (MANLAB, CABA, Argentina).Serum levels of MG-H1 protein adducts were quantified by Methylglyoxal Competitive ELISA kit (OxiSelect Methylglyoxal, Cell Biolabs, San Diego, California, USA). Briefly, aliquots taken were immediately frozen until analysis by ELISA. The ELISA assay was carried out according to the method of Engvall [19]. MG-H1 samples (unknown and standard) were added to each well of a 96-well MG-conjugate coated plate and incubated for 10 minutes. Then, anti-MG antibody was added to each well and incubated for 1 hour. The solution was removed, and the plate was washed with PBS three times. Each well was filled with 100μl of the secondary antibody horseradish-peroxidase-conjugate, and the plate was incubated for 1 hour at room temperature. After washing the plate five times with PBS, the hydrogen peroxide and o-phenylenediamine substrate were added to each well (substrate solution, kit component #310807). The reaction was finished by adding 2 mol/l sulfuric acid and the absorbance at 450 nm was measured on a micro-plate reader (Sunrise, Tekan Switzerland). MG-H1 values were obtained comparing the absorbance with the MG-BSA standard curve. Then, MG-H1 values were expressed as the variation between the initial (0 months) and the final (6 months) experimental time measure of MG-H1 (MG-H1 variation = MG final experimental time measure - MG initial experimental time measure).
Statistical analysis
First at all, parametric conditions of data were checked with kurtosis and bias values, and Kolmogorov and t tests. All data were expressed as means±standard deviation (SD). Significant differences between control and treatments were determined by one-way analysis of variance (ANOVA). Differences were considered significant when p < 0.05.
Results
Lipid metabolism
Serum cholesterol increased significantly under the high-fat diet (HCARDA rabbits, Fig. 2 A). When rabbits received half fat in the diet, serum cholesterol decreased significantly but did not reach normal values (ED2, 1/2 HCARDA, Fig. 2A). If OO was added (1/2 HCARDA + 1/2 OO, Fig. 2A), serum cholesterol reached similar levels to those of control rabbits (NCR). HDL cholesterol measurements showed a decrease in rabbits fed with the high-fat diet (HCARDA and 1/2 HCARDA, Fig. 2B), which recovered with the intake of OO (1/2 HCARDA + 1/2 OO, Fig. 2 B). In relation with serum triglyceride levels, HCARDA showed a slight increment (Fig. 2C).
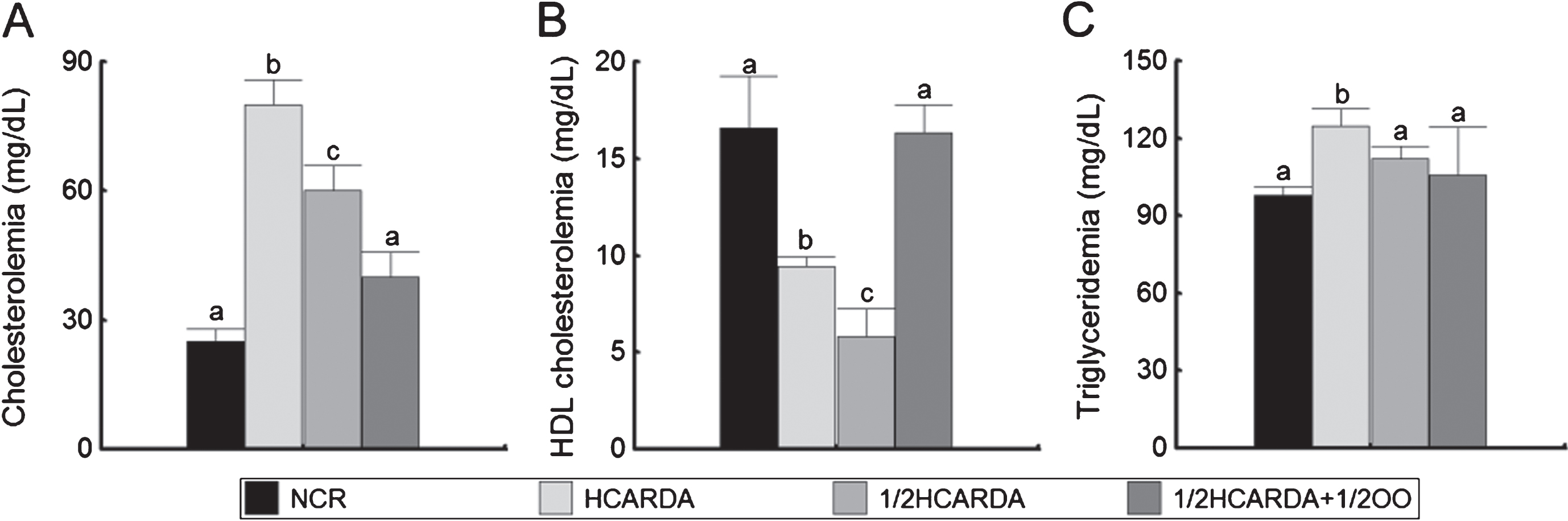
Serum lipids under high-fat and protected diets. Cholesterol (A), HDL cholesterol (B) and triglycerides (C) were measured in serum at the end of the experimental time and expressed in mg/dL. Bars represent the mean±SD of four separate experiments. The color bars correspond to different groups. Different letters indicate statistically different groups (p≤0.05).
Blood glucose was slightly elevated in HCARDA, 1/2 HCARDA and 1/2 HCARDA + 1/2 OO, but within normal values for the species [20] (Fig. 3A). Fructosamine did not change after 6 months of dieting (Fig. 3B). Other useful marker of diabetes, glycosylated hemoglobin did not show significant changes between groups (NCR: 3.4±0.1; HCARDA: 3.6±0.1; and 1/2 HCARDA + 1/2 OO: 3.0±0.1 mg/dl, p < 0.05).
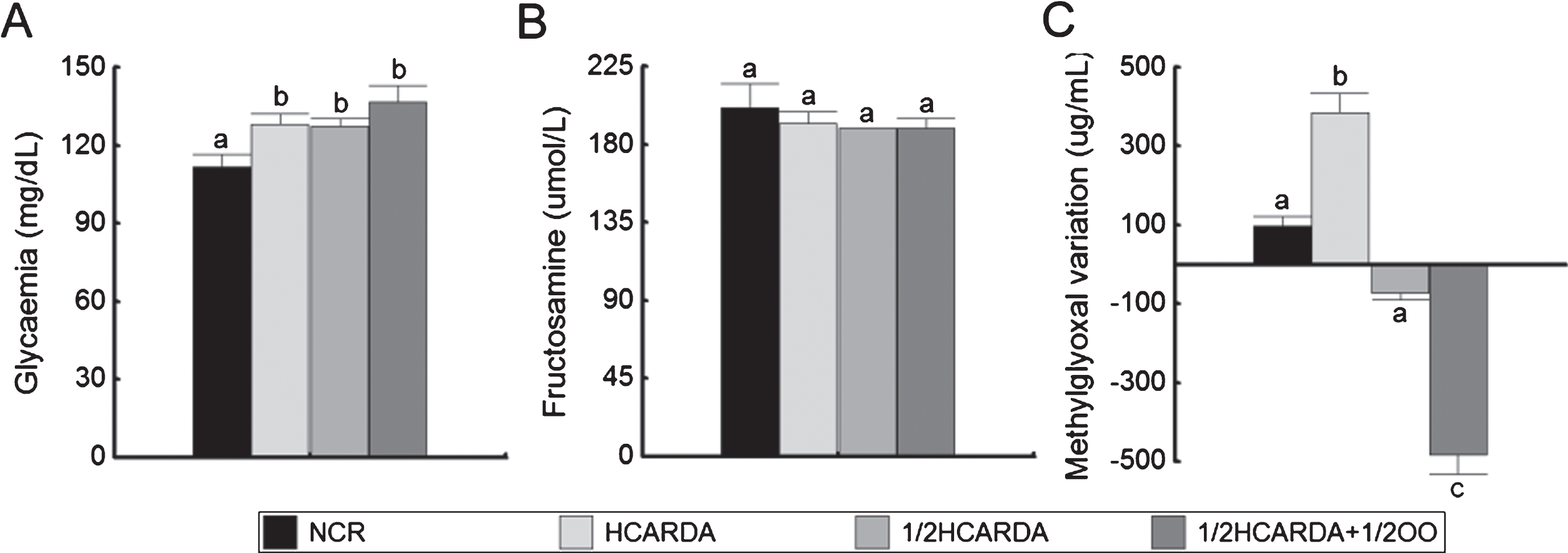
Glycaemia, fructosamine and methylglioxal (MG-H1) under high-fat and protected diets. Glycaemia (A) and fructosamine (B) were determined at the end of the experimental time. MG-H1 (C) was expressed as the difference between initial and final experimental time determinations: MG at the end of the experimental time (6 months) - MG at the beginning of the experimental time (0 months). Bars represent the mean±SD of four separate experiments. The color bars correspond to different groups. Different letters indicate statistically different groups (p≤0.05).
Methylglyoxal (MG-H1) was determined twice, at the beginning and at the end of the experimental period for each group. With this data, the variation of MG-H1 was calculated (Fig. 3C, bars show the differences between the final and initial experimental time determinations). HCARDA rabbits increased MG-H1 values by almost 4 times (Fig. 3C, HCARDA). However, 1/2 HCARDA and 1/2 HCARDA + 1/2 OO reduced their MG-H1 levels (Fig. 3C). Only with the OO intake, the MG-H1 values were reduced 5 times (Fig. 3 C, 1/2 HCARDA + 1/2 OO).
Total and HDL cholesterolemia (serum levels at the end of the experiment), and variation in MG-H1 values were compared under different diets in relation to normal diet (Fig. 4). The NCR values for cholesterol, HDL and MG-H1 were considered as 1, and the other values were relativized to NCR. Under high-fat intake (HCARDA), MG-H1 and cholesterol levels showed similar increasing tends (followed by a decrease in HDL), indicating that both were influenced by the high-fat diet. A reduction in fat intake (ED2 = 1/2 HCARDA) and/or its replacement with OO (ED3 = 1/2 HCARDA + 1/2 OO), promoted a drop in serum cholesterol and a markedly decrement in MG-H1 levels. As described elsewhere, HDL increased with the addition of OO (Fig. 4). In this sense, cholesterol and MG-H1 variations under different experimental diets had a statistical significant correlation (r = 0.96, p < 0.05).
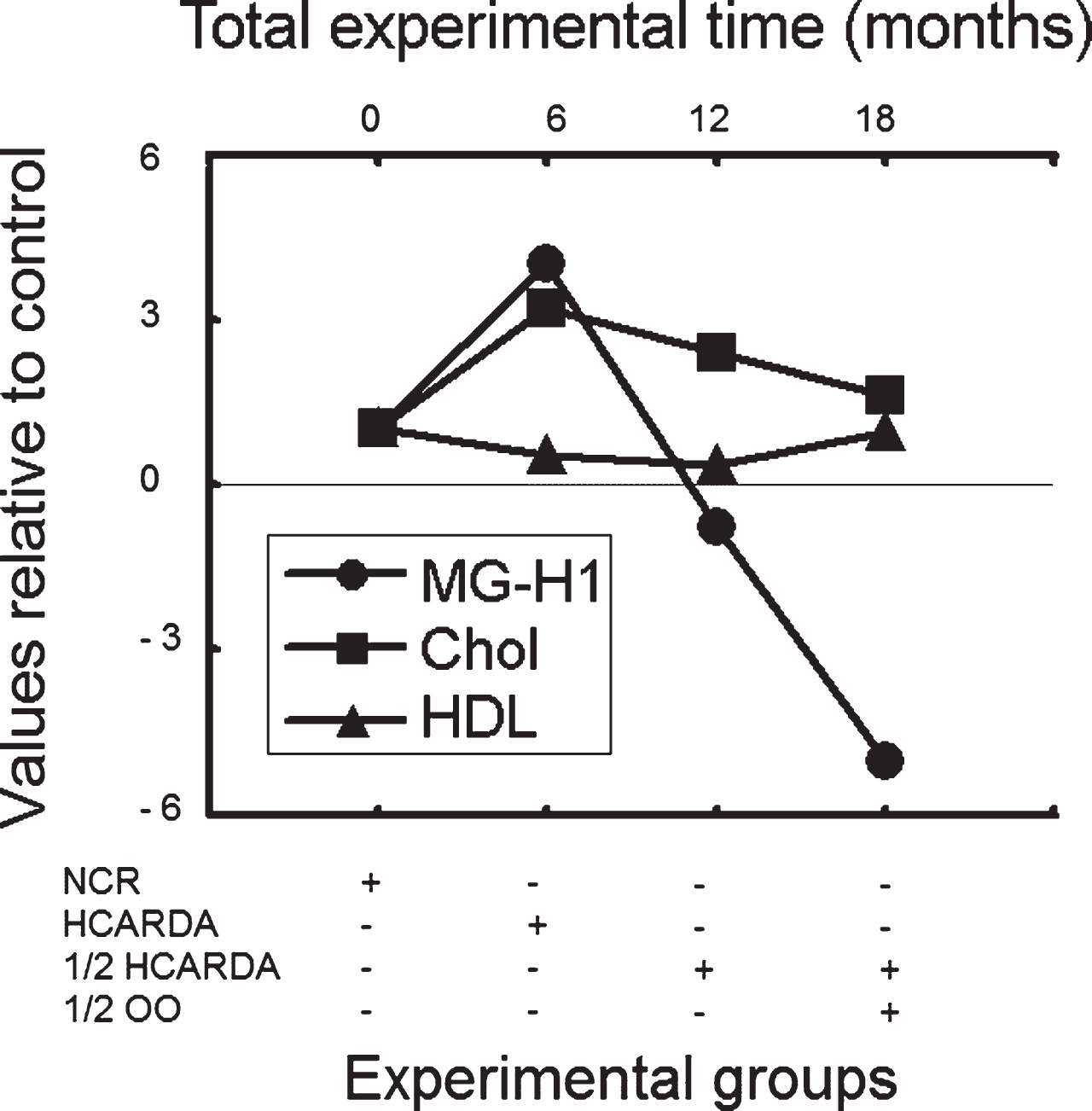
Methylglioxal variation (MG-H1), serum cholesterol (Chol) and HDL cholesterol (HDL) determined at different times and under different diets. The NCR values for cholesterol, HDL and MG-H1 variation were considered as 1, and the values of the other groups were relativized to control (NCR). Total experimental time (months) was plotted in x axis and was related to experimental groups.
We have developed an animal model that faithfully reproduces the human acquired hypercholesterolemic phenotype and its recovery by the olive oil addition to the high-fat diet [17]. These HCARDA rabbits showed hypercholesterolemia, hypertriglyceridemia, and low plasma HDL associated with sub-fertility parameters. In addition, the animals were not the typical diabetic animal model, they had glycaemia and other diabetic markers (such as fructosamine and glycated hemoglobin [21]) in the usual value range for this specie [15, 20]. However, we found that MG-H1 levels increased following serum cholesterol in HCARDA rabbits. Curiously, both parameters were recovered with the addition of olive oil to the diet.
Several authors suggest that methylglyoxal-protein modification is relatively fast and increases with aging, with marked increment in diabetes and kidney failure [22–24]. Glycation of proteins by methylglyoxal forms mainly the hydroimidazolone MG-H1—a quantitatively and functionally important AGE product in physiological systems [5]. Therefore, dicarbonyl glycation contributes significantly to decrease plasma HDL cholesterol, and to increase cardiovascular disease risk in the general population [25]. Moreover, increased MG levels have been correlated with progression from healthy to unhealthy obesity [26].
Elevated MG levels in plasma linked to diabetes complications had been previously attributed to its endogenous production derived from hyperglycemia [27–31]. It has also been suggested that MG can be absorbed from the diet [32, 33], although this is a controversial topic. The source of MG-H1 in HCARDA rabbits could not be hyperglycemia; rabbits had glucose level at usual normal range. We suggest that the source of MG-H1 is related to lipid metabolism, increased under high-fat diet. In this work, we show for the first time that increased MG-H1 is directly associated with serum cholesterol in rabbits (HCARDA). In this way, elevated serum cholesterol appears to be the cause of increased MG-H1 and not its consequence. The underlying mechanism remains unsolved.
Besides, MG has also been shown to adversely affect reverse cholesterol transport in subjects with diabetes, by glycation of apoA-I that inhibits LCAT–mediated esterification of cholesterol in HDL [34]. This impairs removal of cholesterol from peripheral tissues and macrophages, and is associated with atheromatous lesions [35]. In this sense, it was interesting to find a reduction in HDL cholesterol levels associated with an increment in MG-H1 levels in HCARDA rabbits. On the other hand, the addition of olive oil to the high-fat diet promoted an increment in HDL cholesterolemia and a reduction in MG-H1 levels.
The intake of olive oil has been associated with the reduction in cholesterol levels [17, 36]. Some authors have compared olive with other vegetable oils, attributing its beneficial effect to the amount of monounsaturated lipid content [36, 37]. Moreover, an oleic acid-rich diet is a better option in pathologies like diabetes type II, compared with linoleic acid-rich diet [38]. Besides, the antioxidant capacity of plasma increases with olive oil intake. Higher values of coenzyme Q and retinol lead to changes in lipid metabolism [37]. In addition, other components of olive oil, like polyphenols, hydroxytyrosol and α tocopherol, have been associated with beneficial effects [39, 41]. Taken these results together, olive oil intake increases monounsaturated lipids and antioxidants in plasma which could have effect over metabolism, reducing cholesterolemia and MG-H1 levels.
In conclusion, in this work we observed a clear and statistically significant simultaneous increase of MG-H1 and serum cholesterol levels under high-fat diet (enriched in saturated lipids), and a decrease of both with the replacement of the high-fat diet with olive oil (enriched in unsaturated lipids). A major effort must be done to find a link between these important outcomes for clinical treatment.
Conflicts of interest
The authors declare to have no conflict of interest.
Footnotes
Acknowledgments
This work was supported by the research programs from: Proyectos de Investigación Plurianuales (PIP), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Secretaría de Ciencia, Técnica y Posgrado (SECTyP), and Consejo de Investigaciones de la Universidad del Aconcagua (CIUDA).