
Abstract
BACKGROUND:
Due to spatial disorientation reported in space, spatial memory and navigation performances could be more largely impaired by gravity changes. Hippocampus, a key structure for spatial memory, receives inputs from gravity-sensing otolith organs.
OBJECTIVE:
To determine the key molecules in the rat hippocampus that contribute to an adaptation to altered gravity in terms of spatial memory performance.
METHODS:
Gene expression of hippocampus and spatial memory after continuous two-weeks exposure to 2 G hypergravity (HG) were examined using a microarray analysis followed by real-time PCR methods and radial arm maze testing, respectively. Serum cortisol levels during HG load were measured as a stress marker.
RESULTS:
Accuracy to enter the correct arms in HG rats was significantly lower than that of controls, indicating an impaired spatial memory due to gravity changes. Microarray analysis followed by real-time PCR confirmed an upregulation of insulin like growth factor binding protein 2 (IGFBP2) gene. Serum cortisol level was the same level as controls at the last day of hypergravity, suggesting the adaptation to HG-induced stress.
CONCLUSIONS:
Given that the IGF systems are involved in neurotrophic and synaptic plasticity mechanisms, IGF system might contribute to the adaptation to altered gravity in terms of spatial memory.
Keywords
Introduction
In space, lack of stable external cue, namely, gravity causes spatial disorientation for astronauts [9, 46], however, it disappears within a few days perhaps because any adaptive mechanisms to altered gravity would operate in the nervous system. Due to spatial disorientation reported in space, spatial memory and navigation performances could be also more largely impaired by gravity changes. Indeed, spatial learning was impaired after exposure to gravity changes in animals and this impaired performance was also compensated for after long-time exposure to the altered gravity [6, 23], which is similar as seen in the adaptive process of spatial disorientation [9, 46].
Regarding the spatial memory functions, place cells in the hippocampus that fire only when the animal is locating at a specific area in the environment have an important role in these mechanisms [26, 27]. Vestibular system is essential for place-dependent firing of hippocampal place cells by providing an ego-centric cue [33, 45]. Therefore, vestibular information is important in maintaining hippocampal spatial memory function via place cell signaling [10, 37–40], although the anatomical route from the central vestibular system to the hippocampus has not been fully clarified [39, 40].
Given that the essential role of vestibular system in not only sensing gravity but also in spatial memory functions is considered, we hypothesized that molecular and functional rearrangement perhaps occurring in the hippocampus in response to exposure to altered gravity would contribute to the recovery of impaired spatial learning as well as that of spatial disorientation induced by altered gravity. Among the variety of subregions in hippocampus, we chose CA1 area because it contains exclusively place cells that have an important role in spatial orientation and navigation. To test this hypothesis, gene expression of CA1 area of hippocampus in rats after two-weeks exposure to 2 G hypergravity (HG) was examined using a microarray analysis followed by real-time PCR methods. Spatial memory and hypergravity-associated stress were also evaluated by the radial arm maze testing and serum cortisol levels, respectively.
Methods
All experiments were approved by the Animal Care Committee of Osaka University Graduate School of Medicine.
Animals
Male Wistar rats (Japan SLC, Shizuoka, Japan) at seven-weeks old weighing about 150–170 g were used. They were housed individually under a 12-h light/12-h dark cycle (light on at 8 a.m.).
Centrifugation
After handling and shaping, rats (n = 10) were exposed to hypergravity in a centrifuge device for continuous 14 days under the same 12-h light/12-h dark cycle (light on at 8 a.m.) [23, 42]. Rats were placed in a centrifuge cage individually and exposed to 2 G constantly. They could freely access food and water in the cage during centrifugation. Therefore, animals received the Coriolis force during movement in the cage. In order to supply food and water and clean the cage, the centrifuge was stopped for about 20 min at the fourth, 7th and 10th day. The control rats (n = 10) were placed close to the centrifuge device individually but not exposed to hypergravity.
Spatial memory task
Spatial memory task was performed using a radial eight-arm maze as we previously described [23]. Briefly, the animal was transferred to the maze room and placed on the platform of the maze five min later. Then the animal was allowed to run on the maze until all four baits were consumed or five minutes had elapsed. This task was repeated three times a day for two days. Spatial learning was evaluated by the accuracy, which was the rate of correct arm entries divided by total entries. Difference in accuracy of total of six sessions was compared between the animals exposed to hypergravity for two weeks and control animals using Mann-Whitney U test. P < 0.05 was considered significant.
Microarray analysis
Immediately after the last spatial learning task, brains were removed following decapitation under over-dose pentobarbital (100 mg/g, i.p.). CA1 area of bilateral hippocampi was carefully dissected using a micropunch technique [18]. Briefly, coronal sections of the brain block were cut at 200 μm thickness on a cryostat. Each slice was mounted on a slide glass. Under the microscopic guidance, CA1 area was dissected on a chilled aluminum plate with blunted 22 gauge needle using a micropunch technique. Total RNA was extracted from the specimen using RNeasy Mini Kit (Qiagen, Hilden, Germany) and forwarded to a microarray analysis using Code Link Bioarray Rat Whole Genome (Amersham Biosciences, Piscataway, NJ, USA).
Real-time PCR
As described below, some possible genes upregulated in HG rats were identified in CA1 hippocampal area by microarray analysis. To validate the results of the microarray analysis, a real-time quantitative RT-PCR was performed on these genes as we previously described [16]. Briefly, total RNA used for microarray was digested of their genomic DNA, reverse transcribed, and forwarded for TaqMan PCR in ABI7700 Sequence Detection System (Perkin Elmer, USA) with adequate primers designed by Primer Express Software (Table 1). Difference in gene expression was compared between the HG animals and control animals using Mann-Whitney U test. P < 0.05 was considered significant.
Sequences for Igfbp2 primers and TaqMan probe
Sequences for Igfbp2 primers and TaqMan probe
Total of 10 animals were used for serum cortisol measurement. After four-days handling, silicone catheter was placed in the right jugular vein under pentobarbital anesthesia (40 mg/kg, i.p.) for collecting blood samples. After handling to the collecting maneuver for more than two days, five animals were exposed to hypergravity for continuous two weeks. Not only two weeks after exposure to hypergravity but also after one, three, five, and seven days of hypergravity exposure, centrifuge was stopped for 20 min and blood samples were collected. Five control animals were just placed close to the centrifuge device individually without hypergravity exposure. Blood samples were collected at 9 a.m. to control the circadian variation and immediately precipitated and sera were frozen at –80°C. Serum cortisol level was measured using RIA kit (Rat Corticosterone (125 I) Biotrak Assay System with Amerlex-MTM Magnetic Separation, Amersham, U.S.A) according to the manufacturer’s instructions. Difference in serum cortisol was compared between the HG animals and control animals using Mann-Whitney U test. P < 0.05 was considered significant.
Results
Spatial learning task
The accuracy to enter the correct arms in HG rats was significantly lower than that of the controls (Fig. 1).
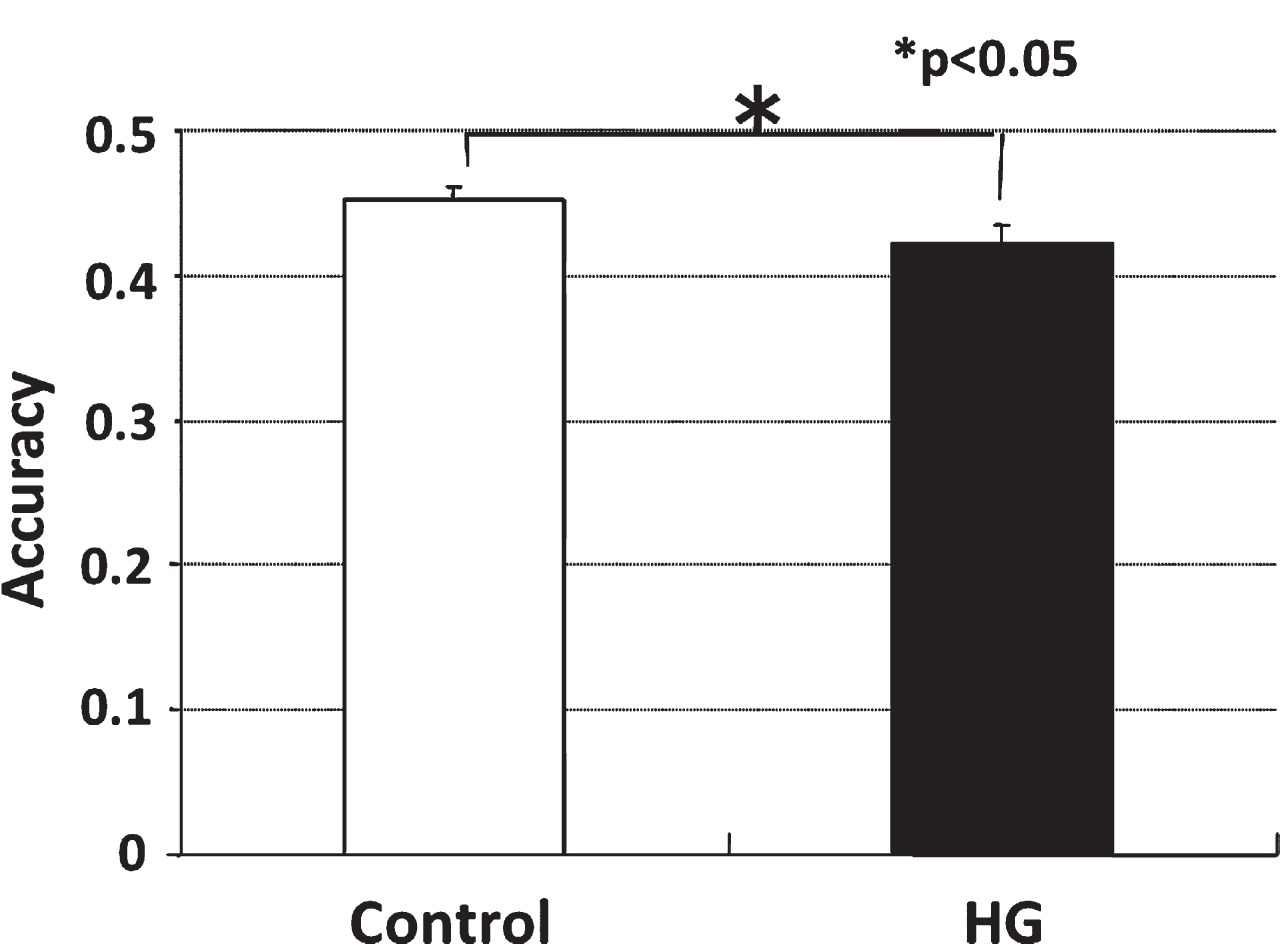
Accuracy of radial arm maze testing of control and HG rats. The accuracy to enter the correct arms in HG rats was significantly lower than that of the controls.
At the first and third day of hypergravity exposure, serum cortisol level was significantly higher in HG rats than controls. After that, serum cortisol level decreased gradually day by day and it was the same level as controls at 14th day of hypergravity (Fig. 2). Although the centrifuge was stopped for short time to collect the samples, provide food and water, and clean the cage, these gradual decreases in serum cortisol levels indicated that the short-term stoppings of the centrifugation seemed to have no actual influences on animals’ status.
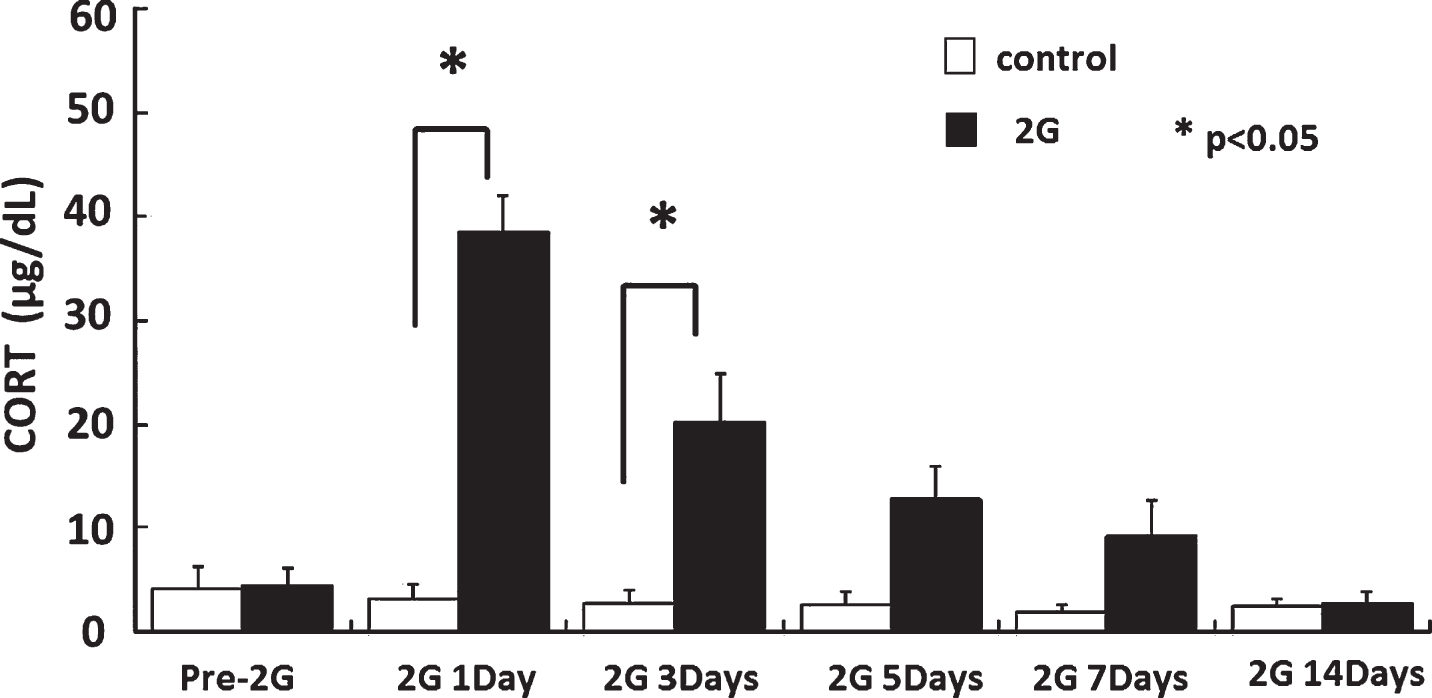
Serum cortisol levels during hypergravity exposure. At the first and third day of hypergravity exposure, serum cortisol level was significantly higher in HG rats than controls. After that, serum cortisol level decreased and it was the same level as controls at 14th day of hypergravity.
The microarray analysis revealed 227 genes upregulated more than 1.3 times in CA1 area of hippocampus of HG rats. They were divided into 37 known genes and 190 expressed sequence tag (EST) sequences. Functional classification of 37 genes was shown in Table 2. Among these 37 genes, we focused on three genes related to insulin-like growth factor (IGF) systems because of their neurotrophic nature and possible role in plasticity mechanisms: insulin-like growth factor 2 (Igf2), insulin-like growth factor binding protein 2 (Igfbp2) and insulin-like growth factor binding protein 3 (Igfbp3). To confirm the increased expression of these genes, specimens were forwarded to the quantitative real-time PCR analysis.
Functional classification of genes upregulated more than 1.3 times after two-weeks hypergravity load
Functional classification of genes upregulated more than 1.3 times after two-weeks hypergravity load
The expression of Igfbp2 was significantly higher in the CA1 area of the HG rats than of the controls (Fig. 3). The expression of Igf2 tended to be higher in HG rats but the difference was not significant and no change was evident in that of Igfbp3 (data not shown).
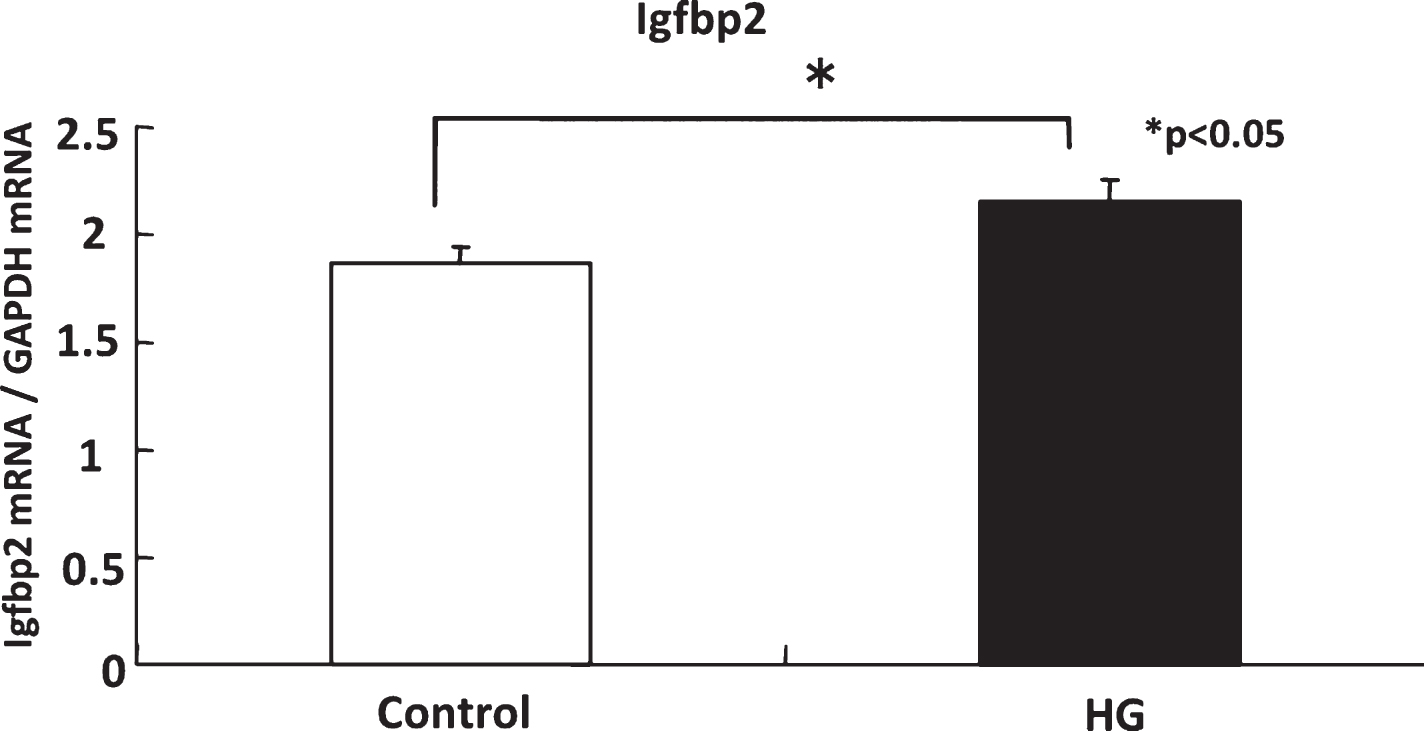
Gene expression of Igfbp2 in control and HG rats. The expression of Igfbp2 in hippocampus was significantly higher in the HG rats than of the controls.
Consistent with our previous studies [23], spatial learning was impaired after two-weeks exposure to hypergravity (Fig. 1), probably because HG rats had already adapted to 2 G environment during two-weeks exposure to hypergravity and that acute changes in gravity from 2 G to 1 G caused a disruption of adaptation. Regarding the molecular background of adaptation to altered gravity, changes in expression of IGF family might be one of candidates as discussed below. Moreover, normal blood cortisol levels at this stage suggest that these molecular and behavioral changes were not caused by non-specific stressful stimuli by altered gravity.
Gravity is being sensed by otolith organs of the vestibular system, and in turn, altered gravity could also affect both of the peripheral and central vestibular systems [15, 44]. Therefore, these changes in peripheral and central vestibular systems might contribute to the adaptation to the altered gravity by changing the sensitivity to linear acceleration including gravity. Recently, accumulated evidence suggested that the vestibular system has an important role in higher cognitive brain function especially of the spatial memory through its influences on place cell functions in hippocampus [37–40]. Therefore, in the present study, we hypothesized that more central mechanisms such as vestibulo-hippocampal pathways could be involved in the adaptation to altered gravity. In order to explore the molecular background of adaptation to the altered gravity, microarray analysis obtained from hippocampal tissue of HG rats as well as control rats was performed for a screening purpose. Microarray analysis revealed 227 genes upregulated more than 1.3 times in CA1 area of hippocampus of HG rats compared with controls. They were divided into 37 known genes (Table 2) and 190 expressed sequence tag (EST) sequences. Del Signore also conducted microarray analysis in the mouse hippocampus which underwent one hour daily exposure to 2 G for five days and reported an upregulation of several genes related to DNA/RNA metabolism, protein processing, intermediate metabolism, cytoskeleton and motility, cell cycle and apoptosis, signal transduction, and neuronal structure/function [12]. Genes related to IGF system were not identified by their study, probably because of differences in animals used (mouse vs. rat) and pattern of hyprgravity exposure (daily one hour for five days in their study vs. consecutive 14 days in the present study). Among the 37 known genes, we concentrated on genes of IGF family because of its neurotrophic nature and its role in synaptic plasticity [2, 34]. Among three genes examined, upregulation of Igfbp2 was confirmed by the real-time PCR methods (Fig. 3).
One might suggest that upregulation of Igfbp2 is just a stress effect of long-term exposure to hypergravity. It is well known that stress signals can activate hypothalamo-pituitary-adrenal gland axis via hippocampal pathways, resulting in elevation of serum cortisol levels [11]. As shown in Fig. 2, at the first and third day of hypergravity exposure, serum cortisol level was significantly higher in HG animals than controls. After that, serum cortisol decreased gradually day by day and it was the same level as the controls at the 14th day of hypergravity. These results suggest that rats adapted to HG environment at this time point in terms of HG-induced stress, at which microarray analysis and spatial memory task were performed. It is not likely that upregulation of Igfbp2 was due to non-specific stressful stimuli by hypergravity exposure. Furthermore, it was reported that prenatal stress [7] or dexamethasone [20] decreased but not increased the Igfbp2 gene expression in the hippocampus or serum Igfbp2 levels followed by down regulation of IGF-1 in rats or humans. Although the mechanisms of upregulation of Igfbp2 gene were not clear, this change may be followed by the upregulation of IGF-1 and it would exert the neurotrophic effects on the CNS. Given that the IGF system is involved in dynamic mechanisms in neuronal plasticity as described below, we suggest that IGF system might help the HG rats to adapt the environment of altered gravity by its role in synaptic plasticity in terms of spatial memory.
Insulin-like growth factor system, which is broadly expressed in the central nervous system particularly in hippocampus, has a role in plasticity mechanisms with neurotrophic effects [2, 34]. Overall, IGF has a significant role in neurogenesis, interference of apoptosis, myelination, and synaptic plasticity in cellular level [1, 21]. In functional level, IGF is important for the recovery of cognition, aging, and traumatic brain injury [4, 35]. For instance, administration of IGF-1, one of insulin-like growth factors, increases excitatory post-synaptic potential in the CA1 area of hippocampus [28] and it modulates expression of NMDAR2A and NMDAR2B subtypes of NMDA receptor, which is known to involve the synaptic plasticity in hippocampus of rats [36]. Moreover, IGF-1 ameliorates age-related impairment in the spatial memory task [19]. IGF binding proteins including IGFBP2 are considered to modulate IGF-1 function. Taken into consideration of this cellular and functional significance of IGF systems, we suggest that elevation of Igfbp2 in hippocampus might help the HG rats to adapt the altered gravity and recovery of spatial memory by its role in synaptic plasticity. Although significant, differences in Igfbp2 gene expression between HG rats and controls were not so large. To investigate the functional role of IGF, further studies such as to examine the effects of anti-IGF sera in spatial learning are required to clarify this point.
In conclusions, two-weeks exposure of hypergravity induced spatial memory deficits in rats. In these animals, Igfbp2 gene was upregulated without elevation of serum cortisol levels. Upregulation of IGF systems might contribute to the adaptation to altered gravitational environment.
Footnotes
Acknowledgments
This study was partly supported by Grants-in-Aid for scientific research from the Ministry of Education, Culture, Sports, Science and Technology of Japan for AH (No. 15591812).