
Abstract
Malignant cells show increased glucose uptake, which is thought to be mediated by glucose transporters. Glucose transporter protein 1 (Glut-1) is critical for growth, proliferation, and migration of tumor cells and Glut-1 overexpression is associated with poor overall survival in osteosarcoma patients. The present study was designed to determine the role of Glut-1 in the growth and invasion of the osteosarcoma cell line MG63, using RNA interference technology in vitro. shRNA-expressing lentiviral vectors targeting the Glut-1 gene were constructed, which were stably expressed in MG63 cells. The level of Glut-1 mRNA was investigated using real-time reverse transcription–polymerase chain reaction, and the protein expression of Glut-1 mRNA was observed using western blotting. MG63 cellular glucose uptake, proliferation, and migration were detected by methyl thiazole tetrazolium assay and flow cytometry. A Glut-1–specific shRNA-expressing lentiviral vector was obtained, which could efficiently inhibit the mRNA and protein expression of Glut-1 to ∼82%–85% in MG63 cells. Downregulation of Glut-1 inhibited the cellular glucose uptake, growth, and invasion of MG63 cells in vitro. These results indicate that RNA interference targeting of Glut-1 could be an effective strategy for the treatment of osteoscarcoma patients.
Introduction
It has long been recognized that malignant cells exhibit increased glucose uptake and utilization in comparison to their nonmalignant counterparts. 1 Energy-independent glucose transport across the plasma membrane is carried out by members of the glucose transporter (Glut) family. The human genes encoding these proteins are named Glut l–5 and Glut 7–13; Glut-6 and Glut-14 are now known to be pseudogenes. 2 Glut-1 appears to be the most ubiquitously distributed of the 14 isoforms. 3,4 This isoform is overexpressed in many human cancer cells, and its appearance is correlated with poor clinical outcome. 5 Glut-1 expression is associated with an increased expression of some proteins, increased glucose uptake, and increased glucose metabolism that can provide extra energy for cancer cell growth to survive in a harsh microenvironment. 6 Endo et al. reported the overexpression of Glut-1 in bone and soft tissue sarcomas and the expression was closely correlated to the prognosis of the patients. 7,8 It was suggested that increased glucose uptake may provide extra energy for osteosarcoma cell growth.
RNA interference (RNAi) is the sequence-specific, post-transcriptional gene-silencing method initiated by double-stranded RNAs that are homologous to the suppressed gene. 9 RNAi is viewed as an important new therapeutic tool for treatment of human conditions, including chronic infection, cancers, and neurodegenerative diseases. 10
In this study, lentivirus encoding shRNA against Glut-l was constructed and transfected into the osteosarcoma cell line MG63 to determine the role of Glut-1 in cellular glucose uptake, growth, and invasion of MG63 cells. Further, the in vitro anticancer activity of this shRNA was investigated. Results from these experiments provide evidence that targeting of Glut-1 is a new strategy for the treatment of osteosarcoma patients who are limited by a 5-year survival rate of about 70%.
Materials and Methods
Cell culture and lenti-shGlut-1 construction and production
The human osteosarcoma cell line MG63 (ATCC) and 293T cells (ATCC) were cultured in RPMI 1640 medium supplemented with 10% fetal calf serum, 100 μg/mL streptomycin, and 100 units/mL ampicillin. Lenti-shGlut-1 was constructed by inserting annealed oligo DNA AGCTGACCCTGAAGTTCATCT (targeting nucleotides 126–144) to PLVTHM (kindly provided by Didier Trono) inserted at the MluI/ClaI site. Lentiviral vectors were produced by transient transfection of 293T cells. Briefly, the transfection mixture consisted of 20 μg lentiviral vector, 10 μg pMDlg/pRRE, 5 μg pMD2.G, and 5 μg pRSV-REV (kindly provided by Didier Trono). The final volume was adjusted to 250 μL with water, and 250 μL of 0.5 M CaCl2 was added to the solution and mixed well. Subsequently, 500 μL of HeBS2x (0.28 M NaCl, 0.05 M N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid, 1.5 M Na2HPO4) was added and the solution was incubated for 30 minutes. The culture dishes were placed into a 37°C humidified incubator with 5% CO2. Fourteen hours later, the medium was aspirated, and 10 mL of fresh Dulbecco's modified Eagle's medium Y with 10% fetal bovine serum (PAA), prewarmed to 37°C, was gently added. Cultures were further incubated for 28 hours, after which virus was collected. Concentrated supernatants were titrated with serial dilutions of vector stocks on 1 × 105 HeLa cells, followed by fluorescence-activated cytometric analysis (Beckton Dickinson Immunocytometry Systems). Titers of lenti-sh-Glut-1 vectors were calculated to be between 5 × 108 TU/mL. MG63 cells were subsequently infected with lentivirus. An RNAi control plasmid was constructed for the green fluorescent protein (GFP) gene.
Real-time reverse transcription–polymerase chain reaction
Total RNA was isolated from cultured cells and real-time reverse transcription (RT)–polymerase chain reaction (PCR) was performed using the RNeasy and one-step RT-PCR kit from Qiagen Corp. RT-PCR of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), a housekeeping gene, served as a control. The primer sequences used were 5′-TTTATTGCAGCCAGAGCCACCAGCG-3′ and 5′-ACAGAAAAGATGGCCACTGAG-3′ for Glut-1 (250 bp), 3 and 5-ACCACAGTCCATGCCATCAC-3′ and 5′-TCACCACCCTGTTGCTG TA-3′ (antisense) for GAPDH (100 bp). 6 For RT-PCR, two pairs of primers were added into a reaction tube, and real-time PCR was carried out according to KIT from Takara Bio. The program consists of an initial RT at 50°C for 30 minutes, denaturation at 95°C for 10 minutes, followed by 24 cycles of amplification (denaturation at 95°C for 30 seconds, annealing at 55°C for 1 minute, and extension at 68°C for 1 minute), and a final extension at 68°C for 10 minutes. The products were then resolved by 1.5% agarose gel electrophoresis, and the bands were visualized using ethidium bromide and UV light and analyzed by Genetools software.
Western blotting
Cells were harvested at 3 days post-transfection and washed twice with 10 mL of PBS. The cells were treated with radioimmunoprecipitation assay lysis buffer (Watson). Protein (25.2 g) was resolved by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrophoretically transferred onto a polyvinylidene difluoride membrane. Membranes were incubated with a primary anti–Glut-1 rabbit polyclonal antibody (1:500; Santa Cruz Biotechnology) for 48 hours at 4°C. The protein was quantified with an avidin–biotin–peroxidase complex (ABC) kit (Beyondtime) to control for equal sample loading in each group, and the quantification of individual bands was normalized by simultaneously incubating the membranes with antibodies against GAPDH (Kangcheng) as a standard.
Glucose stimulation experiment
Cell culture and transfection with siRNA-Glut-1 or empty expression vector
Nontransfected, mock-transfected, and siRNA-Glut-1–transfected MG63 cells were seeded in six-well plates (7.0 × 105 cells/well). After 24 hours incubation, 5, 10, or 20 mM glucose was added to begin the glucose stimulation test, as previously described. 11 After incubation for 0.5 or 1 hour, cell culture supernatants were collected.
Determination of glucose uptake in the supernatant of MG63 cells
Glucose uptake from the culture medium was determined using a detection kit (Shanghai Kehua Biotech). The glucose concentration in RPMI 1640 medium was 10.31 mmol/L. The glucose uptake in the supernatant of cells was calculated as follows: concentration of glucose uptake (mM) = concentration of glucose stimulation + glucose concentration of medium − concentration of glucose in supernatant.
Measurement of cell proliferation
Cell proliferation was measured using the methyl thiazole tetrazolium (MTT) assay. 12 Cell suspensions were seeded in 96-well plates at a density of 1 × 104 cells/well. After 24 hours incubation, 20 μL of 5 mg/μL solution of MTT (Sigma) in PBS was added to each well. The plates were then incubated for 4 hours at 37°C. The supernatant was then removed and the precipitate was solubilized in 150 μL dimethyl sulfoxide (Sigma-Aldrich) and shaken for 15 minutes. Absorbance was determined with an enzyme-linked immunosorbent assay reader (model 318) at 490 nm. Each assay was performed nine times. The results were expressed as mean ± standard error of controls.
Cell invasion assay
Cell invasion assays were performed using Transwell polycarbonate membrane inserts in 24-well plates (Corning) following the manufacturer's instructions. Briefly, the underside of each polycarbonate microporous membrane was coated with Matrigel (1:100) at 37°C for 5 minutes and allowed to sit overnight. Then, 20 μL Matrigel (1:6) and 200 μL sterile water were added to the upper compartment at 37°C. After 2 days, 200 μL of the invasion buffer (2 mL bovine serum albumin [2%] + 38 mL RPMI 1640) was added into the upper compartment, and 1 hour later, the upper compartment fluid was aspirated. Cells at a density of 1 × 105 cells/well were added into the upper compartment, and 300 μL serum-containing medium was added into the lower compartment. The cells were allowed to migrate for 48 hours. The inserts were stained with hematoxylin and eosin, fixed in 10% formalin for 10 minutes, and rinsed by dipping in water. The cells on the upper surface of the membrane were removed with a cotton bud. The membranes were air-dried overnight, excised from the insert, and mounted onto glass slides for microscopic analysis. The migrated cells were counted at high-power magnification (200 × ) from eight randomly selected fields. Each experiment was repeated three times.
Cell cycle analysis
MG63, GFP-MG63, and MG63-shGlut-1 cells were seeded overnight on 6-cm-diameter plates and placed in a serum-free RPMI 1640 medium for 24 hours to synchronize. RPMI 1640 medium containing serum was then added, and after 24 hours, cells were harvested. After washing with ice-cold PBS, cells were suspended in about 0.5 mL of 70% alcohol and kept at 4°C for 12 hours. Then cells were treated with RNase A (Boehringer Mannheim) and stained with propidium iodide (Sigma-Aldrich). DNA content was analyzed by flow cytometry using the FACScalibur cell analyzer (BD Biosciences). Cell cycle was analyzed using Multicycle-DNA Cell Cycle Analyzed Software. Each experiment was performed in triplicate.
Statistical analysis
All statistical analyses were performed using SPSS software (version 13.0; SPSS). Data were presented as mean ± standard deviation. Numerical data were analyzed by Student's t-test. A p-value of < 0.05 was considered to be statistically significant.
Results
Glut-1 mRNA inhibition in MG63 cells
Glut-1–specific shRNA-expressing lentiviral vectors, which could efficiently inhibit the expression of Glut-1, were generated (Fig. 1). Lenti-shGlut-1 coexpresses enhanced GFP, so that infected MG63 cells (MOI = 20) can be detected using fluorescent microscopy (Fig. 1A, B). Enhanced GFP expression efficiency was about 95% (Fig. 1C). Real-time RT-PCR results show that Glut-1 mRNA expression in MG63 was strongly inhibited by lenti-shGlut-1, and the inhibition efficiency was about 82% (Fig. 1D).
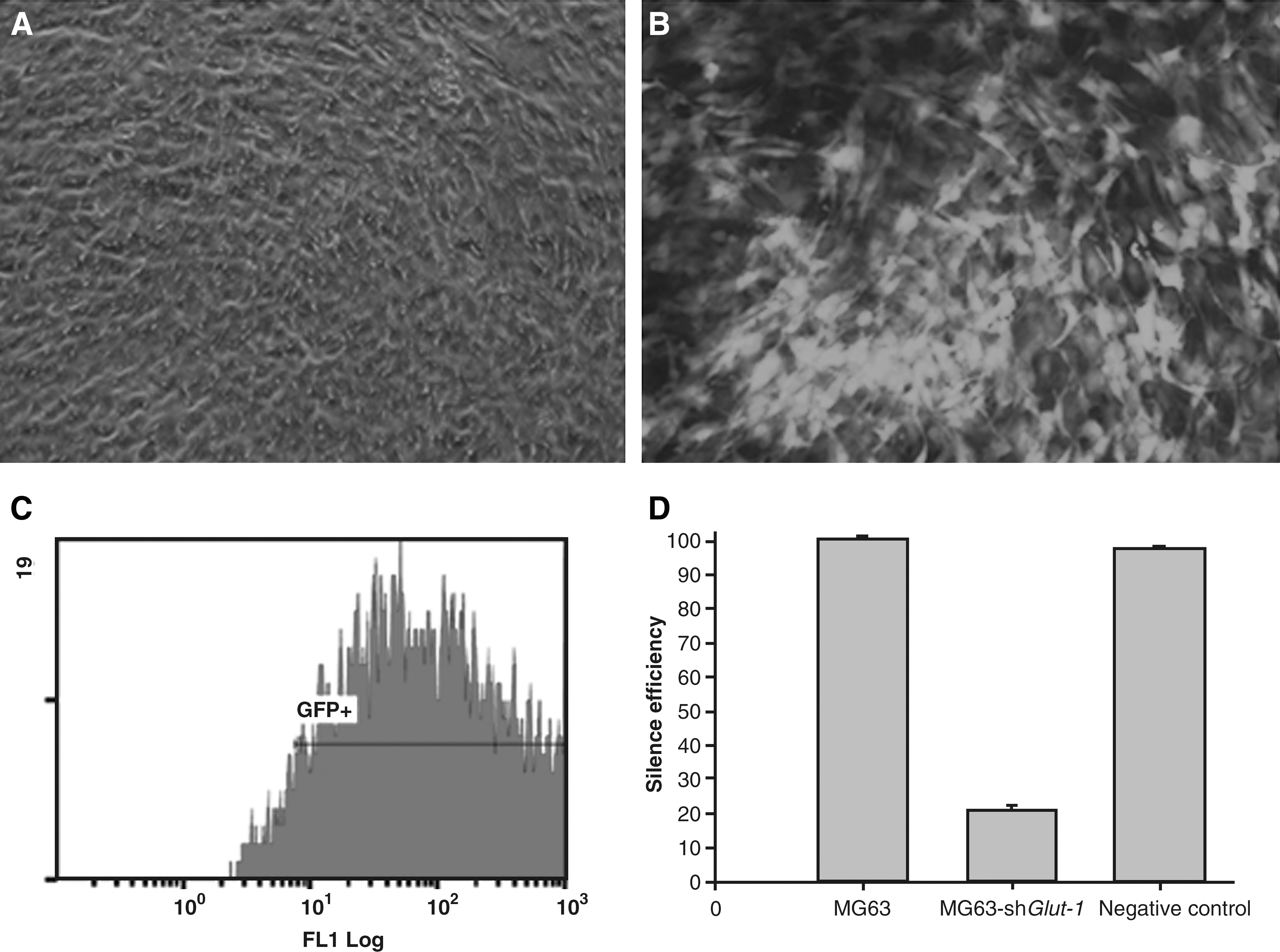
Lenti-shGlut-1–infected MG63 cells (MOI = 20) coexpress enhanced GFP
Glut-1 protein inhibition in MG63 cells
The inhibition of lenti-shGlut-1 in MG63 cells compared with the control group was detected by western blotting (Fig. 2). Stably transfected cells had a marked reduction in Glut-1 expression and about 85% reduction in Glut-1 synthesis (Fig. 2).

Expression of Glut-1 in MG63 cells. Western blotting was performed to examine the protein expression level of Glut-1 in
Effect of siRNA-Glut-1 on glucose uptake in MG63 cells
MG63 cells transfected with siRNA-Glut-1 showed decreased glucose uptake at 0.5 and 1 hour after glucose stimulation compared with non- and mock-transfected MG63 cells (p < 0.05; Table 1; Fig. 3).
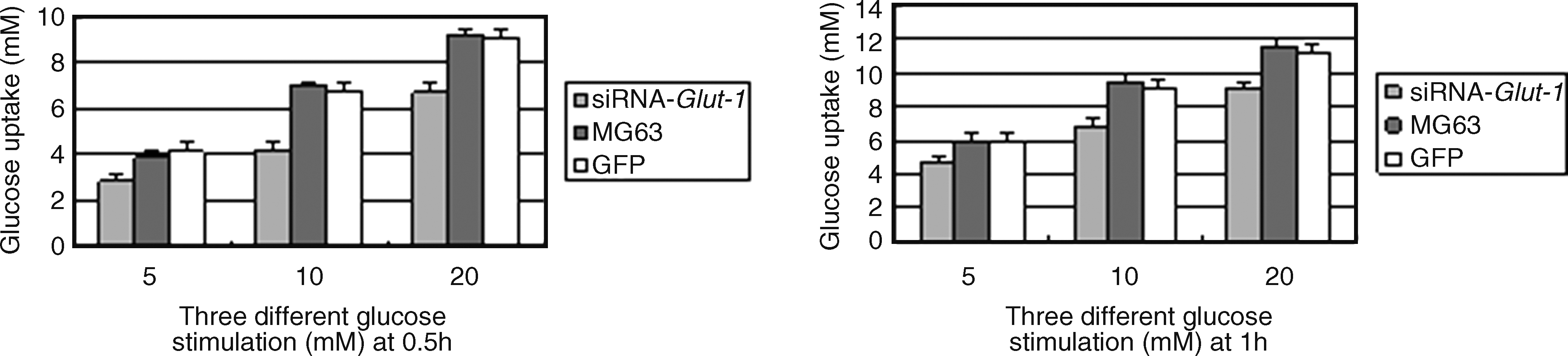
Effect of sh-Glut-1 on glucose uptake in MG63 cells. At 0.5 and 1 hour after glucose stimulation, MG63 cells transfected with shRNA-Glut-1 showed decreased glucose uptake compared with non- or mock-transfected cells.
The values are medians with standard errors. Compared with MG63 and empty vector, at 0.5 and 1 hour after glucose stimulation, the results showed decreased glucose uptake in the MG63 cells transfected with siRNA-Glut-1.
p < 0.05, compared with the results of siRNA-Glut-1 and MG63 or GFP.
GST, glucose stimulation time; GSC, glucose stimulation concentration; GFP, green fluorescent protein (empty vector; shown in black and white); Glut-1, glucose transporter protein 1.
Effect of siRNA-Glut-1 on proliferation of MG63 cells
The effect of siRNA-Glut-1 transfection on the proliferation rate of the MG63 cell line was investigated using MTT assay. The results summarized in Figure 4 demonstrate that transfection of siRNA-Glut-1 decreased MG63 cell proliferation over time compared with non- or mock-transfected cells (p < 0.05).
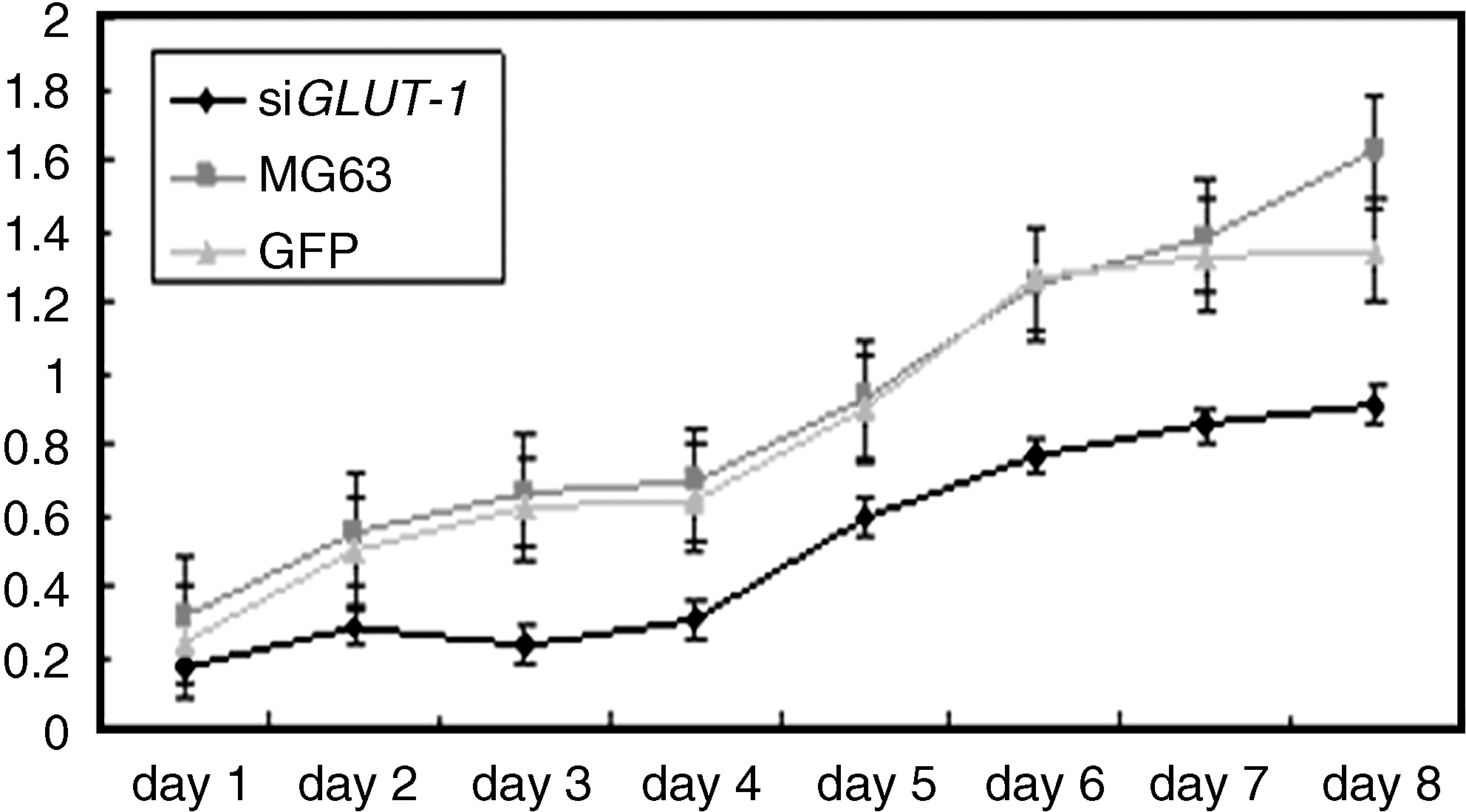
Inhibition of MG63 cell proliferation by siRNA-Glut-1 transfection. MTT reagent was added to each sample at 1–8 days post-transfection. Absorbance at 490 nm was determined for each sample. Differences in OD490 values between siRNA-Glut-1–transfected cells and GFP or parental MG63 cells were statistically significant (p < 0.05).
Effects of siRNA-Glut-1 on MG63 cell invasion/migration
As shown in Figure 5, the number of migrated MG63 cells was significantly reduced in the siRNA-Glut-1 group for each 200 × field under the microscope compared with nontransfected (29.75 ± 6.27 vs. 39.88 ± 6.36, p < 0.01) or mock-transfected (29.75 ± 6.27 vs. 37.38 ± 5.52, p < 0.01) cells.
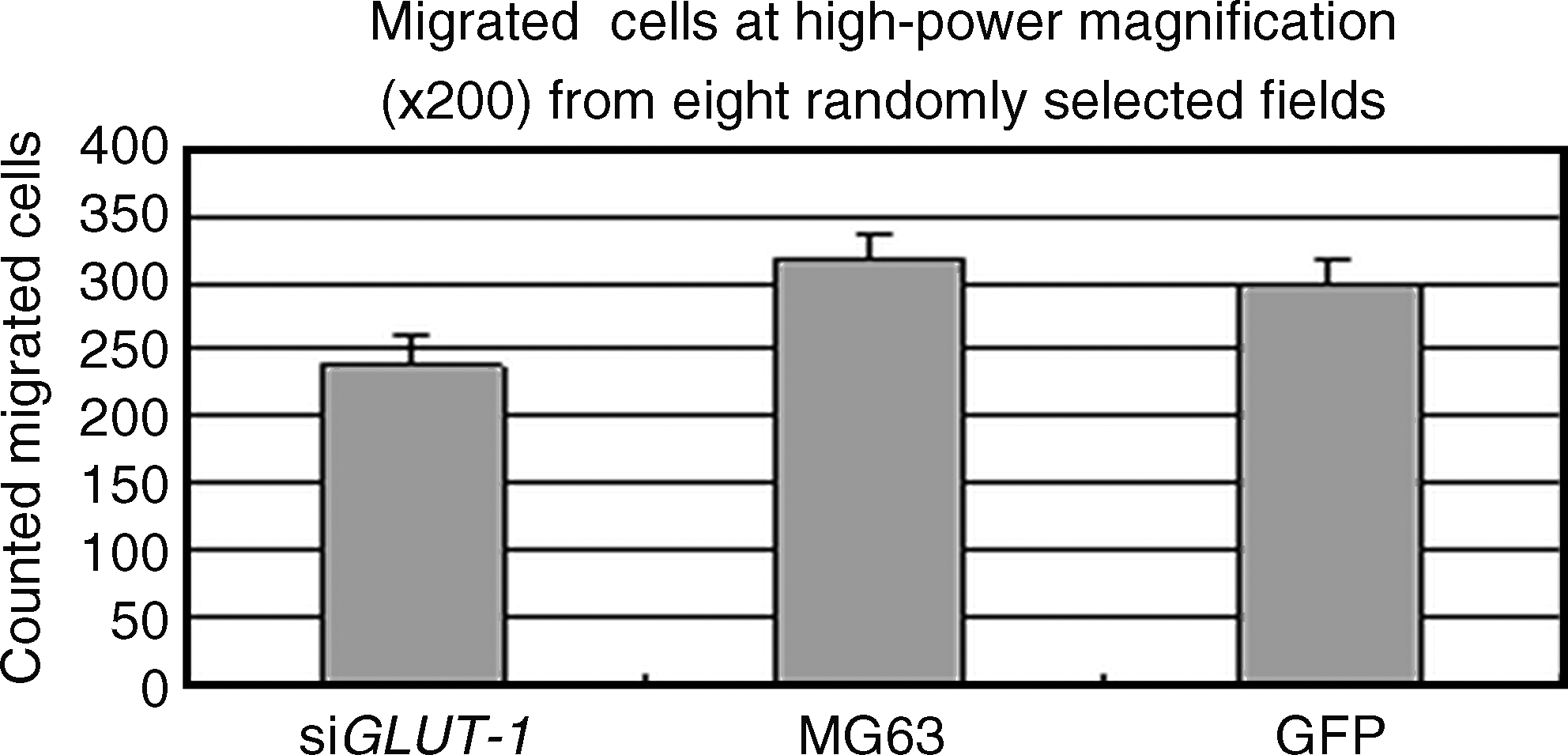
Effects of Glut-1 siRNA on MG63 cell migration. Migration assays were performed in Transwell polycarbonate membrane inserts in 24-well plates. The siGlut-1, MG63, and GFP medium were added to the lower chambers. The cells were allowed to migrate for 48 hours and the migrated cells were counted at high-power magnification (200 × ) from eight randomly selected fields.
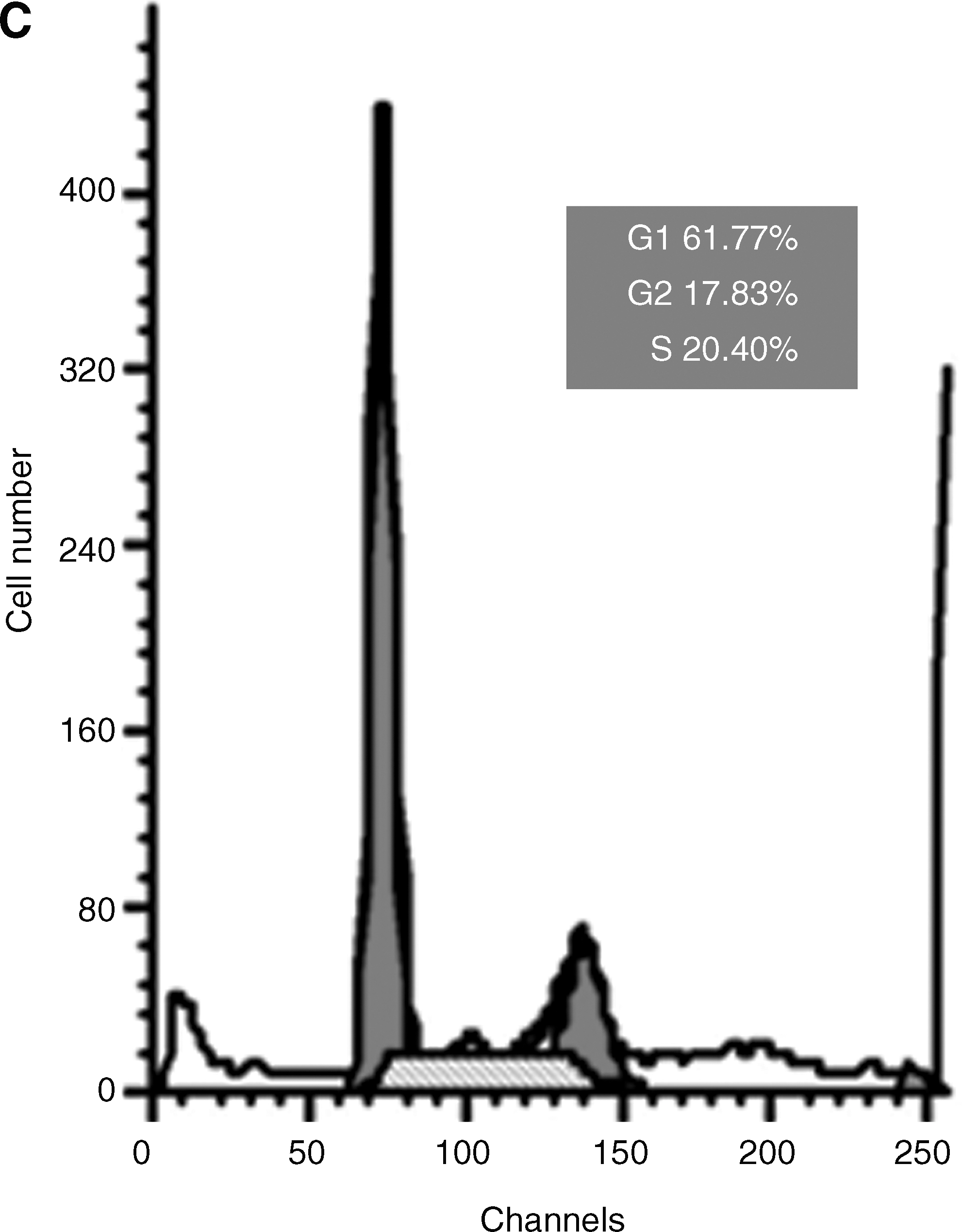
Transfection with siRNA-Glut-1 induces a block in G1 phase of the cell cycle
The conditioned medium of siRNA-Glut-1 was also found to have an effect on the MG63 cell cycle (Fig. 6). Cells transfected with siRNA-Glut-1 had an increased number of cells in the G1 phase (61.77%), which was significantly higher than in negative control cells (39.08%) or untransfected MG63 cells (28.82%). These results suggest that the conditioned medium of siRNA-Glut-1 arrests MG63 cells in the G1 phase.
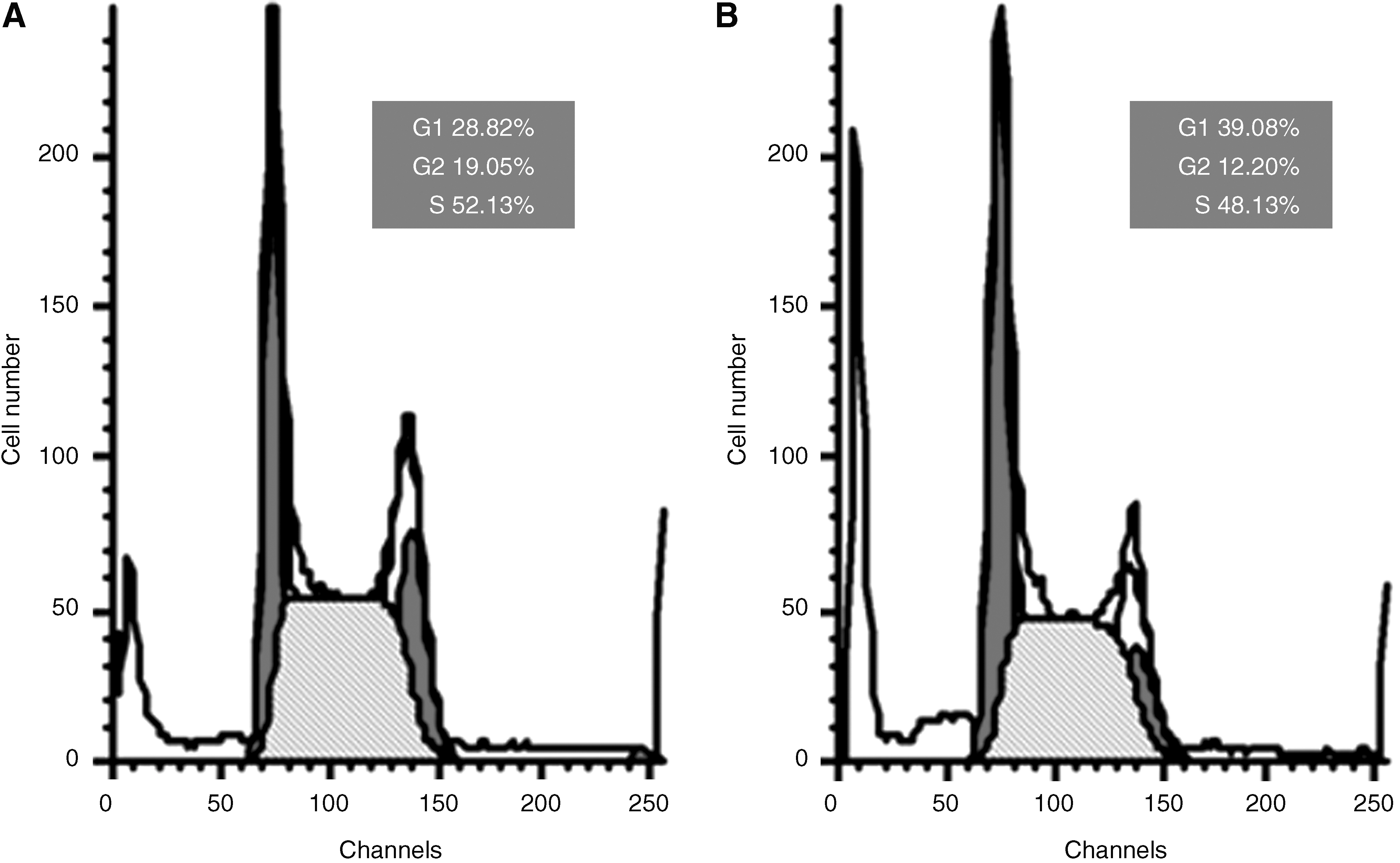
Effects of Glut-1 siRNA on MG63 cell cycle distribution. G1-phase block was induced by siRNA-Glut-1. The cell cycle profiles for
Discussion
Despite the advent of neoadjuvant chemotherapy, wide excision of tumors, and even the amputation of the affected limbs, the 70% 5-year survival rate in osteosarcoma patients has been scarcely increased. 13 The development of gene therapies using RNAi offers new therapeutic possibilities. It is the process of sequence-specific gene silencing and is believed to represent an ancestral form of nucleic acid–based immunity against intracellular pathogens, which results in a sequence-specific silencing of gene expression. 14,15 Gene products contributing to the transformation of cells, tumor growth, and invasion are all potential drug targets for RNAi. Therefore, RNAi is viewed as a powerful tool to define gene function and as a novel therapeutic strategy for treatment of human diseases and conditions, including cancer.
Glut-1 expression appears to reflect the biological behavior of cancer cells, because the expression of Glut-1 is regulated by conditions related to cell proliferation, differentiation, and nutritional starvation in many tumors. 3 The results of overexpressing Glut-1 are consistent with increased glucose uptake, increased adaptive changes to glycolytic metabolism, and increased cellular proliferation in cancer cells. Many studies have shown that Glut-1 is overexpressed in human tumors and increased expression of Glut-1 has been shown to be correlated with a poor prognosis and may be a malignant tumor marker. 6,16,17
Overexpression of Glut-1 was also found in osteosarcoma cells, and osteosarcoma patients with Glut-1 overexpression showed significantly poor overall survival. 7 However, the direct mechanism by which Glut-1 expression is associated with poor prognosis is not clearly understood and few studies have focused on the application of siRNA-Glut-1 as an osteosarcoma therapy. 18
In the present study, Glut-1-siRNA was constructed using lentivirus as a vector, to transduce MG63 cells. The knockdown of Glut-1 efficiently reduced the transcript levels of Glut-1 mRNAs and ultimately resulted in the reduction of Glut-1 protein levels compared with untransfected cells and cells transfected with control vector siRNAs. Further, this inhibition was shown to be highly selective and sequence specific, because transfection with nonspecific siRNA had no effects on Glut-1 expression, confirming the successful construction of Glut-1-siRNA-MG63 cells.
To further investigate the effect of selective Glut-1 inhibition and the role of Glut-1 in growth and invasion of the human osteosarcoma cell line MG63, the effects of Glut-1 knockdown on glucose uptake, proliferation, migration, and cell cycle status were investigated. Decreased glucose uptake in cells transfected with siRNA-Glut-1 was observed, which can suppress glucose metabolism in osteosarcoma cells, compared with the controls. In addition, knockdown of Glut-1 expression caused a statistically significant reduction in cell growth and inhibited cell proliferation and migration of MG63 cells in vitro. Further, cells transfected with siRNA-Glut-1 were blocked in G1 phase. This arrest in G1 phase likely contributes to the observed antiproliferative and antitumor effects observed in siRNA-Glut-1–treated tumor cells.
In conclusion, it has been demonstrated that siRNA-Glut-1 decreased Glut-1 expression in the osteosarcoma cell line MG63 and inhibited tumor cell activities, such as glucose uptake, proliferation, invasion, and cell cycle, in vitro. The findings of the present study also indicate that Glut-1 may represent a potential therapeutic target for strategies designed to inhibit the progression of osteosarcoma. To the authors' knowledge, the present study is the first to report lentivirus-mediated RNAi of Glut-1 suppression in osteosarcoma cells. The next step will be to determine the effects of Glut-1 knockdown in vivo as a prelude to possible applications in humans. However, the regulator family of Glut is very complicated, and it is still unknown how other positive or negative factors change following the downregulation of Glut-1 over a long period. Further research is needed to determine how to develop safer vectors and how to carry out multiple-target gene therapy through RNAi.
Conclusions
RNAi of Glut-1 inhibits the growth and invasion of the osteosarcoma cell line MG63 in vitro and its effects in vivo need to be tested further. RNAi of Glut-1 may be a promising approach in treating osteosarcoma patients.
Footnotes
Disclosure Statement
The authors declare that they have no proprietary, financial, professional, or other personal interest of any nature or kind in any product, service, and/or company that could be construed as influencing the position presented in this manuscript.