
Abstract
Radiation plays an important role in the treatment of hepatoma. In order to improve its therapeutic ratio, there has been much interest in augmenting the effect of radiation on tumors by combining it with molecularly targeted therapeutics. Hypoxia-inducible factor-1 (HIF-1) is an excellent potential candidate for targeted molecular therapy to improve radiation outcome. In this study, HIF-1α-targeted small interfering RNA (siRNA) expression vector was constructed and transfected into human hepatoma SMMC-7721 cells, which followed by culture in CoCl2-induced hypoxia. HIF-1α downregulation by siRNA inhibited proliferation, induced apoptosis, and enhanced radiosensitivity in chemical hypoxic SMMC-7721 cells in vitro. These findings suggest that specific inhibition of HIF-1α expression in combination with radiotherapy would be expected to exert a strong antitumor effect on human hepatoma.
Introduction
Owing to an imbalance in oxygen supply and demand, hypoxia is an important characteristic feature of solid tumors, such as hepatoma. Hypoxia is an important contributor to tumor radioresistance. 1 A large body of clinical data has been published demonstrating that hypoxia negatively influences radiotherapy. Oxygen can chemically modify the radiation-induced DNA damage, thereby producing adducts that are difficult to repair by cells. The phenomenon that oxygen increases the cytotoxicity of radiation, resulting in roughly a three-fold difference in radiosensitivity between hypoxic and aerobic cells, is termed the oxygen effect. The oxygen effect has been proposed as one of the mechanisms underlying hypoxia-induced radioresistance. 2 Hypoxia can abolish the oxygen effect and lead to decreased radiosensitivity.
In addition, hypoxia may also modulate tumor radiosensitivity through biological effects. 3 Signaling pathways stimulated by hypoxia are commonly activated in tumors. Hypoxia-inducible factor 1 (HIF-1) is one of the key mediators of hypoxia signaling pathways. 4 HIF-1 is a heterodimeric transcription factor consisting of two subunits, HIF-1α and HIF-1β. HIF-1β is constitutively expressed, whereas HIF-1α is rapidly degraded in normoxic conditions and can be stabilized by hypoxia and hypoxic mimicking agents including cobalt chloride, nickel chloride, and desferrioxamine. Therefore, hypoxia can induce HIF-1 expression, which modulates more than 100 genes involved in regulating important processes such as angiogenesis, metabolism, proliferation, and apoptosis. 5 Recently, several reports have demonstrated that HIF-1α plays an important role in tumor radioresistance, and downregulating its expression could improve the tumor response to irradiation in certain cancer types. 6 –8
RNA interference (RNAi) is a phenomenon in which the introduction of double-stranded RNA (dsRNA) into a diverse range of organisms and cell types causes degradation of the complementary mRNA. RNAi is characterized by high efficiency, sequence-specific gene suppression, and simplicity of experimental procedure. RNAi has emerged as a powerfully post-transcriptional gene silencing technique widely utilized in cancer research. 9 In the current study, HIF-1α-targeted small interfering RNA (siRNA) expression vector was constructed and transfected into human hepatoma SMMC-7721 cells, which followed by culture in CoCl2-induced hypoxia. Cell proliferation, apoptosis, radiosensitivity, and some hypoxia related genes expression were detected to explore the effect of HIF-1α-short hairpin RNA (shRNA) on radiosensitivity of chemical hypoxic SMMC-7721 cells and its mechanism.
Materials and Methods
shRNA design and plasmid construction
The cDNA sequence of HIF-1α was obtained from GenBank (NM-001530). The siRNA target design tools from Ambion were used to design HIF-1α-shRNA. HIF-1α-shRNA was designed and synthesized as follows (Sangon, Inc., Shanghai, China): sense: 5′-GATCCTGTGAGTTCGCATCTTGATTTCAAGAGAATCAAGATGCGAACTCACATTTTTTGGTACCGGAAA-3′, antisense: 5′-AGCTTTTCCGGTACCAAAAAATGTGAGTTCGCATCTTGATTCTCTTGAAATCAAGATGCGAACTCACAG-3′. The oligonucleotides were annealed and inserted into the BamHI and HindIII sites of pGenesil-2 according to the manufacturer's instructions (Genesil, Inc., Wuhan, China). The recombinant vector pGenesil-HIF was confirmed by the digestion analysis of restriction endonuclease, and all inserted sequences were verified by DNA sequencing.
Cell line and cell culture
Human hepatoma SMMC-7721 cell line was cultured in Roswell Park Memorial Institute 1640 (RPMI 1640) (Gibco BRL, Grand Island, NY) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, 100 mg/mL streptomycin, and various concentrations CoCl2 (Sigma, St Louis, MO) in a 37°C incubator in a 5% CO2 humidified atmosphere.
Transfection conditions
SMMC-7721 cells were divided into three groups: the recombinant vector pGenesil-HIF transfected cells, the negative control vector pGenesil-negative transfected cells, and untransfected cells taken as control. SMMC-7721 cells were seeded in six-well plates at 2.0×104 cells/well and cultured to 80% confluence. Transfection was performed by using LipofectAMINE (Gibco BRL, Gaithersburg, MD) according to the manufacturer's instruction. Solution A was prepared by diluting 5 μg pGenesil-HIF or pGenesil-negative into 100 μL serum-free media. Solution B was prepared by diluting 10 μL LipofectAMINE into 100 μL serum-free media. The two solutions were gently mixed and incubated at room temperature for 30–45 minutes. The mixture and 800 μL serum-free media were added into each well. The cells were incubated at 37°C for 6 hours, and then, the transfection media were replaced by fresh complete growth media. CoCl2 was added into the media 12 hours after transfection.
Reverse transcription–polymerase chain reaction analysis of HIF-1α mRNA expression
Total cellular RNA was isolated by using Trizol reagent (Sangon, Inc.) according to the manufacturer's protocol after 24 hours of chemical hypoxic culture. Total RNA was reverse transcribed into cDNA, which was used for polymerase chain reaction (PCR) amplification. The thermal cycles were as follows: 1 cycle of 94°C for 5 minutes; 30 cycles of 94°C for 30 seconds, 56°C for 30 seconds, 72°C for 45 seconds; followed by a final elongation step of 72°C for 10 minutes. The primer sequences were as follows: HIF-1α (311 bp), forward: 5′-GTGAGTTCGCATCTTGATA-3′, reverse: 5′-CCATTTCTGTGTGTAAGCA-3′; β-actin used as internal control (474 bp), forward: 5′-AAATCGTGCGTGACATTAA-3′, reverse: 5′- CTCGTCATACTCCTGCTTG-3′. Reverse transcription (RT)-PCR products were electrophoresed in a 1.5% agarose gel with ethidium bromide. Quantification of HIF-1α gene expression was based on densitometry readings (BioRad Laboratories, Hercules, CA).
Western blot analysis of protein expression
Cells were harvested by exposure to trypsin–ethylene diamine tetraacetic acid (EDTA), washed with phosphate-buffered saline (PBS), and lysed in a solution containing1% Triton X-100, 10% glycerol, 1 mM EDTA, 1 mM phenylmethylsulfonylfluoride, and 10 mM Tris–HCl [pH 7.4]. Cell debris was removed by centrifugation at 10,000g for 10 minutes at 4°C. Equal amounts of lysate protein were fractionated by sodium dodecylsulfonate–polyacrylamide gel electrophoresis at 100 V for 80 minutes at room temperature. The separated proteins were transferred to a nitrocellulose membrane, which was then probed for 2 hours at room temperature with rabbit monoclonal anti-HIF-1α, glucose transporter-1 (GLUT-1), vascular endothelial growth factor (VEGF), B-cell leukemia-2 (Bcl-2), survivin, proliferating cell nuclear antigen (PCNA) (Santa Cruz, Inc., Santa Cruz, CA), and rabbit polyclonal anti-β-actin (Sigma). Immune complexes were detected with horseradish peroxidase-conjugated goat antibodies to rabbit immunoglobulin G (Amersham Biosciences, Little Chalfont, UK). Immunoblots were visualized by chemiluminescence by using a chemiluminescence kit (Invitrogen, Carlsbad, CA), and the specific bands were recorded on an X-ray film. Actin protein levels were used as a control to verify equal protein loading.
Cell proliferation assay
The cell viability of SMMC-7721 cells was measured by a 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay (Sigma) after 24, 48, and 72 hours of chemical hypoxic culture. The above mentioned three groups of SMMC-7721 cells, at 4.0×103 per well, were seeded into 96-well culture plates. In fact, 200 μL MTT (5 mg/mL) was added to each well, and the cells were incubated at 37°C for an additional 4 hours. After incubation, MTT-containing medium was removed, and 150 μL of dimethyl sulfoxide was added to each well to dissolve formazan crystals. Optical densities were determined on a Versamax microplate reader (Molecular Devices, Sunnyvale, CA) at 490 nm.
Flow cytometric analysis of cell cycle
SMMC-7721 cells were harvested and fixed overnight with 70% ethanol at 4°C, followed by resuspension in 500 μL of PBS. After addition of 10 μL RNase (10 mg/mL), cells were left for 30 minutes at 37°C and stained with 10 μL propidium iodide (PI) (1 mg/mL). Cellular DNA content was determined on a flow cytometer Beckton Dickinson (BD) FACScan (BD Biosciences, San Jose, CA) with an exciting wavelength of 488 nm. The relative proportion of cells in the G0/G1, S, and G2/M phases of the cell cycle were determined by flow cytometry data.
Flow cytometric analysis of apoptosis
Quantification of apoptotic cells was performed according to the Annexin-V-fluorescein isothiocyanate (FITC) manufacturer's instructions (KeyGen Biotech. Nanjing, China). Briefly, cells were collected and resuspended in 500 μL binding buffer, and 2 μL Annexin-V-FITC and 5 μL of PI were added. Analyses were performed by a flow cytometer (BD FACScan) with Ex=488 nm, Em=530 nm. FITC-positive and PI-negative cells were regarded as apoptotic cells. The sample was incubated for 5 minutes in the dark before analysis by a flow cytometer.
Clonogenic cell survival assay
After 72 hours of chemical hypoxic culture, 1.0×103 SMMC-7721 cells plated in six-well plates were irradiated with 0, 2, 4, 6, and 8 Gy X-ray irradiation (6 MV, the dose rate was 200 cGy/min) by a PRIMUS accelerator (SIEMENS Medical Solutions, Erlangen, Germany) at room temperature. Then, the cells were incubated for 8 days. Colonies were stained with crystal violet, and colonies of more than 50 cells were counted. Plating efficiency was calculated as follows: plating efficiency=(colony number/plating cell number) ×100%, in triplicate. Further, the cell survival fraction was counted, and the cell survival curve was drafted by the standard model, S=1−(1−e −D/D0) N (S, cell survival fraction; D, radiation dose; e, the bottom of the natural logarithm; D 0, the mean death dose; N, extrapolate number). Finally, the radiosensitization ratio was calculated (a ratio of D 0).
Data analysis
All data were calculated as mean±standard deviation. Comparisons between treatment groups and controls were made by a t-test. A p-value less than 0.05 between groups was considered to be a significant difference.
Results
HIF-1α protein expression in SMMC-7721 cells induced by various concentrations CoCl2
HIF-1α protein expression in SMMC-7721 cells induced by various concentrations (up to 200 μmol/L) CoCl2 were measured by Western blot. Experiments were repeated thrice. The relative levels of HIF-1α protein expression were normalized against protein levels of an internal control gene, β-actin, performed in the same run. As shown in Figure 1, no expression of HIF-1α protein was detected in SMMC-7721 cells without CoCl2 treatment,, probably due to the quick degradation of the protein under normoxic conditions. HIF-1α protein expression was observed in SMMC-7721 cells treated with 150 or 200 μmol/L CoCl2 for 12 or 24 hours. Since no difference in HIF-1α protein expression was observed after treatment with 150 or 200 μmol/L CoCl2 for 12 hours (51.33%±3.51% and 56.20%±3.02% respectively; p=0.146) or 24 hours (59.43%±4.62% and 63.03%±8.50% respectively; p=0.257), 150 μmol/L CoCl2 was chosen for the rest of the experiments.

Detection of HIF-1α protein expression in SMMC-7721 cells induced by various concentrations CoCl2 for 12 and 24 hours by Western blot. Immunoblots were visualized by chemiluminescence, and the specific bands were recorded on an X-ray film. Actin protein levels were used as a control to verify equal protein loading. Experiments were repeated thrice. HIF-1α, hypoxia-inducible factor-1α.
Specific downregulation of HIF-1α expression by HIF-1α-shRNA in SMMC-7721 cells after 24 hours of chemical hypoxic culture
The mRNA and protein expression intensities of HIF-1α gene, inhibited by HIF-1α-shRNAs in chemical hypoxic SMMC-7721 cells, were analyzed by semiquantitive RT-PCR and Western blotting. Experiments were repeated thrice. The relative levels of HIF-1α mRNA and protein expression were normalized against mRNA and protein levels of an internal control gene, β-actin, performed in the same run. As shown in Figure 2, the level of HIF-1α mRNA in SMMC-7721 cells transfected with pGenesil-HIF was significantly decreased compared with that of SMMC-7721 cells (42.27%±3.33% and 117.23%±8.21% respectively; p=0.0036). The inhibitory rate of HIF-1α mRNA expression was 63.91%. In contrast, no obvious change of HIF-1α mRNA expression was observed in SMMC-7721 cells transfected with pGenesil-negative compared with that of SMMC-7721 cells (102.60%±12.91%; p=0.166). Additionally, Western blot analysis showed a 58.75%-downregulated level of HIF-1α protein expression in SMMC-7721 cells transfected with pGenesil-HIF compared with that of SMMC-7721 cells (22.97%±2.78% and 55.67%±2.52% respectively; p=0.0008). No obvious change of HIF-1α protein expression was observed in SMMC-7721 cells transfected with pGenesil-negative compared with that of SMMC-7721 cells as shown in Figure 3 (52.67±4.04%; p=0.136). The results just mentioned indicated that the expression of HIF-1α gene in chemical hypoxic SMMC-7721 cells could be specifically and effectively downregulated by siRNA expression plasmid pGenesil-HIF.

Effect of HIF-1α-shRNA on HIF-1α mRNA expression in SMMC-7721 cells after 24 hours of chemical hypoxic culture. Total cellular RNA was isolated by using Trizol reagent after 24 hours of chemical hypoxic culture. Total RNA was reverse transcribed into cDNA, which was used for PCR amplification. Reverse transcription–PCR products were electrophoresed in a 1.5% agarose gel with ethidium bromide. Quantification of HIF-1α gene expression was based on densitometry readings. Experiments were repeated thrice. PCR, polymerase chain reaction.

Effect of HIF-1α-shRNA on HIF-1α protein expression in SMMC-7721 cells after 24 hours of chemical hypoxic culture. HIF-1α protein expression in chemical hypoxic SMMC-7721 cells was detected by Western blot. Actin protein levels were used as a control to verify equal protein loading. Experiments were repeated thrice. shRNA, short hairpin RNA.
Effect of HIF-1α-shRNA on chemical hypoxic SMMC-7721 cell proliferation
We investigated the effect of HIF-1α-shRNA on chemical hypoxic SMMC-7721 cell proliferation in vitro by MTT assay. Cell viability was detected after 24, 48, and 72 hours of chemical hypoxic culture. Figure 4 showed that transfection with siRNA expression plasmid pGenesil-HIF decreased viability of chemical hypoxic SMMC-7721 cells in a time-dependent manner (p<0.05 or p<0.01), and the highest inhibitory rate was 31.18% after 72 hours of chemical hypoxic culture. All these results indicated that shRNA-mediated downregulation of HIF-1α expression significantly suppressed chemical hypoxic human hepatoma SMMC-7721 cell proliferation in vitro.

Effect of HIF-1α-shRNA on chemical hypoxic SMMC-7721 cell proliferation. Cell viability was detected by MTT assay after 24, 48, and 72 hours of chemical hypoxic culture. Data represent the average of five experiments. Error bars represent one standard deviation. *p<0.05, **p<0.01 versus control.
Effect of HIF-1α-shRNA on chemical hypoxic SMMC-7721 cell cycle
To investigate the mechanism underlying the HIF-1α-shRNA mediated cell proliferation suppression, we analyzed the cell cycle of SMMC-7721 cells by flow cytometry after 72 hours of chemical hypoxic culture. As shown in Figure 5, the percentage of G0/G1 phase cells in SMMC-7721 cells transfected with pGenesil-HIF was increased (p<0.01), and no significant change was seen in the G2/M phase cells, although the percentage of S phase cells was significantly reduced compared with that of SMMC-7721 cells (p<0.01). These results indicated that chemical hypoxic SMMC-7721 cells transfected with pGenesil-HIF were arrested in the G0/G1 phase of cell cycle.

Effect of HIF-1α-shRNA on chemical hypoxic SMMC-7721 cell cycle. Cell-cycle distribution was detected by flow cytometry after 72 hours of chemical hypoxic culture. Data represent the average of six experiments. Error bars represent one standard deviation. *p<0.01 versus control.
Effect of HIF-1α-shRNA on apoptosis of chemical hypoxic SMMC-7721 cells
Apoptosis of SMMC-7721 cells was quantitatively measured by using flow cytometry after 24, 48, and 72 hours of chemical hypoxic culture. As shown in Figure 6, the average apoptotic rate of SMMC-7721 cells transfected with pGenesil-HIF was significantly increased compared with that of SMMC-7721 cells (p<0.01) after 48 and 72 hours of chemical hypoxic culture. No obvious change of apoptotic rate was observed in SMMC-7721 cells or SMMC-7721 cells transfected with pGenesil-negative after 24, 48, and 72 hours of chemical hypoxic culture.
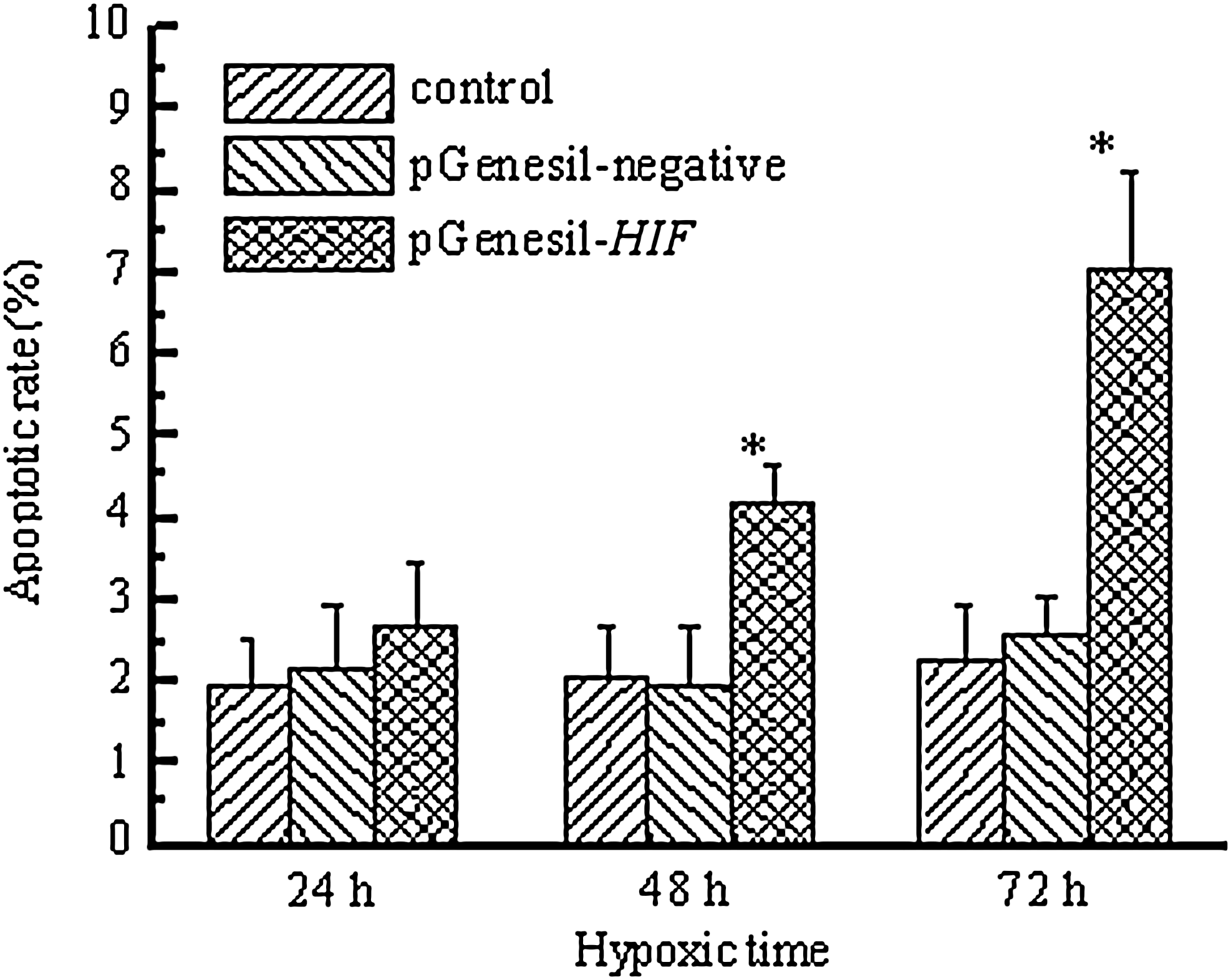
Effect of HIF-1α-shRNA on apoptosis of chemical hypoxic SMMC-7721 cells. Cell apoptosis was detected by flow cytometry after 24, 48, and 72 hours of chemical hypoxic culture. Data represent the average of six experiments. Error bars represent one standard deviation. *p<0.01 versus control.
Effect of HIF-1α-shRNA on radiosensitivity of chemical hypoxic SMMC-7721 cells
After exposure to 0, 2, 4, 6, and 8 Gy X-ray irradiation, SMMC-7721 cell survival fractions were examined by using a clonogenic assay. The survival fraction of chemical hypoxic SMMC-7721 cells transfected with pGenesil-HIF at the same dose of radiation was significantly decreased compared with that of chemical hypoxic SMMC-7721 cells (p<0.05 or p<0.01). The survival fraction of chemical hypoxic SMMC-7721 cells transfected with pGenesil-negative at the same dose of radiation showed no difference compared with that of chemical hypoxic SMMC-7721 cells. The survival curves of SMMC-7721 cells, as shown in Figure 7, were obtained from data fitting according to the linear quadratic model. It is clear that chemical hypoxic SMMC-7721 cells transfected with pGenesil-HIF (D 0=3.97) were more sensitive to X-ray irradiation than chemical hypoxic SMMC-7721 cells (D 0=5.75), and the radiosensitization ratio was 1.45. Radiosensitivities of chemical hypoxic SMMC-7721 cells and chemical hypoxic SMMC-7721 cells transfected with pGenesil-negative (D 0=5.62) showed no obvious difference. In addition, normoxic SMMC-7721 cells showed the highest radiosensitivity (D 0=2.60). Thus, we concluded that the downregulation of HIF-1α expression could radiosensitize chemical hypoxic SMMC-7721 cells.

Survival curves of chemical hypoxic SMMC-7721 cells exposed to X-ray irradiation determined by clonogenic assay. After exposure to 0, 2, 4, 6, and 8 Gy X-ray irradiation, SMMC-7721 cell survival fractions were examined by using a clonogenic assay. Curves represent data fitting according to the linear quadratic model. Data represent the average of three experiments. Error bars represent one standard deviation.
Effect of HIF-1α-shRNA on protein expression of hypoxia related genes in chemical hypoxic SMMC-7721 cells
To investigate the molecular mechanism underlying the HIF-1α-shRNA mediated biological effect just mentioned, we analyzed protein expression of hypoxia related genes in SMMC-7721 cells after 72 hours of chemical hypoxic culture by Western blot. Experiments were repeated thrice. The relative levels of hypoxia related protein expression were normalized against protein levels of an internal control gene, β-actin, performed in the same run. As shown in Figure 8, the level of HIF-1α, GLUT-1, VEGF, PCNA, Bcl-2, and survivin protein expression in SMMC-7721 cells transfected with pGenesil-HIF was significantly decreased compared with that of SMMC-7721 cells (p=0.017, 0.038, 0.009, 0.020, 0.006, and 0.019 respectively). The inhibitory rate of HIF-1α, GLUT-1, VEGF, PCNA, Bcl-2, and survivin protein expression was 61.65%, 44.12%, 51.89%, 48.11%, 57.87%, and 46.84% respectively. No obvious change in these hypoxia related protein expressions was observed in SMMC-7721 cells transfected with pGenesil-negative compared with that of SMMC-7721 cells (p=0.35, 0.37, 0.39, 0.42, 0.34, and 0.30 respectively). These results indicated that shRNA-mediated downregulation of HIF-1α expression significantly suppressed protein expression of hypoxia related genes in chemical hypoxic SMMC-7721 cells.

Detection of hypoxia related proteins expression in chemical hypoxic SMMC-7721 cells after 72 hours of chemical hypoxic culture by Western blot. Actin protein levels were used as a control to verify equal protein loading. Experiments were repeated thrice.
Discussion
Human hepatoma remains one of the most common cancer in Africa and Asia and a leading cause of cancer death in Asia, and its incidence is also rising in Western countries. Management of human hepatoma continues to be challenging because of high recurrence rate after surgical resection and resistance to chemotherapy and radiotherapy. 10 –12 Radiation plays an important role in the treatment of hepatoma. In order to improve its therapeutic ratio, there has been much interest in augmenting the effect of radiation on tumors by combining it with molecularly targeted therapeutics.
HIF-1 is an excellent potential candidate for targeted molecular therapy to improve radiation outcome. 13 HIF-1 is a heterodimer consisting of HIF-1α and HIF-1β subunits, both of which are transcription factors. HIF-1β, a nuclear protein also known as the aryl hydrocarbon receptor nuclear translocator, ARNT, is constitutively expressed and independent of oxygen tension; whereas HIF-1α is a cytoplasmic protein responsive to oxygen level. HIF-1α contains an oxygen-dependent degradation domain that is central to the oxygen-regulated stability of this protein and two transactivation domains, N-terminal (N-TAD), and C-terminal (C-TAD) in its C-terminal region. 14,15 These transactivation domains mediate interaction of the HIF transcription factor with other co-activators of gene transcription, such as p300/CBP, that confer target gene specificity. 16,17 In normoxic cells, HIF-1α is continuously degraded, whereas under hypoxic conditions, it translocates to the nucleus and heterodimerizes with HIF-1β to form the active HIF-1 protein, binding and activating hypoxia responsive genes that contain hypoxia response elements, such as VEGF, growth factors, and glycolytic enzymes. Activation of these target genes serves as compensatory mechanisms, thus allowing tumor cells to facilitate the cellular adaptive responses to hypoxia. 18 HIF-1α is overexpressed in many human cancers and has been associated with tumor aggressiveness, vascularity, treatment failure, and mortality. 19,20 There are at least two mechanisms mediating the effect of HIF-1α on tumor radioresistance. First, HIF-1α could directly protect cancer cells from radiation-induced cell death by inhibiting apoptosis signal. Second, HIF-1α increases the expression of proangiogenic cytokines and promotes the survival of stromal endothelial cells, which, in turn, provide support to cancer cells. All these activities make the HIF-1α transcription factor an attractive target for the development of new anticancer therapeutics. 21 However, HIF does not mediate the cellular response to hypoxia independently. A number of additional transcription factors have been shown to be responsive to hypoxia. Among these is the major transcription factor nuclear factor-kappa B (NF-κB). NF-κB is both activated by and induces the expression of cytokines and chemokines. 22 In addition, NF-κB is a regulator of proliferation, apoptosis, angiogenesis, and metastasis. 23 NF-κB activation is a critical component in the transcriptional response to hypoxia. Rocha et al. have found that NF-κB activation after hypoxia is very rapid and persistent in an inhibitor of NF-κB (IκB) kinase (IKK)-dependent manner. 24 Aggarwal et al. have found that inhibition of NF-κB by nimbolide can sensitize tumor cells to chemotherapeutic agents through interaction with IKK. 25 These findings suggest that NF-κB may be another effective therapeutic target for hypoxic tumor cells.
Cobalt is essential for human health because of its critical role in the synthesis of vitamin B12; however, excess exposure of cobalt can lead to the loss of cell viability, the elevation of apoptotic rate, the formation of reactive oxygen species, and the disruption of mitochondrial transmembrane potential. 26 CoCl2 has been widely used as a hypoxia mimicking agent in both in vivo and in vitro studies. 26 –28 CoCl2 treatment induces HIF-1α expression by binding to the prolyl hydroxylases domain, thus resulting in blockage of binding between HIF-1α and the von Hippel-Lindau tumor suppressor protein and thereby HIF-1α stability. 29 In this study, we investigate the effects of the hypoxia-mimetic CoCl2 on HIF-1α gene expression at protein levels in human hepatoma SMMC-7721 cells. We demonstrated that CoCl2 treatment leads to an increase in HIF-1α gene expression, and the optimal CoCl2 concentration was 150 μmol/L.
To explore the possibility of HIF-1α as an effective therapeutic target, we employed RNAi technique to silence HIF-1α expression in chemical hypoxic SMMC-7721 cells and analyzed phenotypic changes after transfection mediated by cationic liposome. 30 Here, we constructed a HIF-1α-targeted siRNA expression plasmid pGenesil-HIF with U6 promoter, which coded for an RNA composed of two identical 19-nucleotide sequence motifs in an inverted orientation, separated by a 9 bp spacer to form a hairpin dsRNA. After transfecting the plasmid into human hepatoma SMMC-7721 cells, we found that the mRNA and protein expression of HIF-1α gene could be specifically downregulated and effectively measured by semiquantitative RT-PCR and Western blot. The downregulation of HIF-1α led to significant cell growth inhibition by MTT assay, cell arrest in the G0/G1 phase of cell cycle, and increased apoptotic rate by FCM assay in chemical hypoxic SMMC-7721 cells.
The role of HIF-1 in radiation sensitivity of cancer cells in vitro has been investigated in several reports. However, the results of these experiments are somewhat controversial. Moeller et al. reported that HIF-1 inhibition enhances the cytotoxic effects of radiation in a clonogenic survival experiment under hypoxic conditions. 13 Zou et al. have reported that the functions of HIF-1 are independent of radiation sensitivity in multicellular tumor spheroids under either normoxic or hypoxic conditions. 31 Vordermark et al. observed a cell type-specific association between HIF-1 protein accumulation levels under hypoxia and cellular radiation resistance. 6 To investigate the effect of HIF-1α-shRNA on radiosensitivity of chemical hypoxic SMMC-7721 cells, cell survival fractions were examined by using a clonogenic assay. 32 Our results showed that the downregulation of HIF-1α expression could radiosensitize chemical hypoxic SMMC-7721 cells. The protein expression of HIF-1α and its related genes, including GLUT1, which facilitates glucose uptake by the cells, VEGF, which directly facilitates tumor angiogenesis, thus accelerating tumor growth, and PCNA, which is a nuclear cell proliferation marker and anti-apoptotic genes Bcl-2 and survivin, were decreased in chemical hypoxic SMMC-7721 cells transfected with pGenesil-HIF. These results indicated that HIF-1α inhibition could enhance tumor radiosensitivity through decreasing glucose metabolic activity and proliferation rates and increasing apoptotic potential in hypoxic SMMC-7721 cells.
Conclusions
In summary, our study indicated that HIF-1α downregulation by siRNA inhibited proliferation, induced apoptosis, and enhanced radiosensitivity in human hepatoma cells in vitro. These findings suggest that specific inhibition of HIF-1α expression in combination with radiotherapy would be expected to exert a strong antitumor effect on human hepatoma.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 30600160) and Program for Changjiang Scholars and Innovative Research Team in University (IRT0849).
Disclosure Statement
No competing financial interests exist.