
Abstract
The purposes of this study were to test the effect of tetrandrine, alone or combined with radiation, on human esophageal cancer cell line TE1 (TE1 cells) and investigate the potential antitumor mechanism. Human esophageal cancer cell line TE1 was tested by methyl thiazolyl tetrazolium assay for cell proliferation, colony-forming assay for cell radiosensitivity, flow cytometry assay for cell cycle distribution, and western blot assay for cell cycle protein expression. When treated alone, tetrandrine had a time- and concentration-dependent cytotoxic effect on TE1 cells. The dose-enhancement ratio for combined tetrandrine and radiation was markedly increased when compared with tetrandrine alone. Further, expression of cyclin B1 protein increased after addition of tetrandrine when compared with radiation only. Radiation-induced G2 arrest was abrogated with treatment of tetrandrine. In conclusion, tetrandrine can enhance the radiosensitivity of TE1 cells and this may involve relief of radiation-induced G2/M arrest in TE1 cells.
Introduction
Esophageal carcinoma is one of the most common cancers, especially in P.R. China. However, treatment for esophageal carcinoma is not optimal with a 5-year survival rate remaining at about 5%–10% with conventional radiotherapy (RT). 1,2 In designing various treatment strategies for malignant tumors, the importance of combining chemotherapy and RT is increasing. However, chemotherapy and/or radiochemotherapy have not demonstrated any survival advantages so far. 3 Clinical trials of accelerated hyperfractionated RT have been carried out with the aim of overcoming this problem by shortening the overall treatment time. Some reports have already suggested improvement in local control and survival rates, but at the expense of increasing acute toxicity, particularly with the faster treatment schedule. 4 –6
Tetrandrine, a bisbenzylisoquinoline alkaloid, was isolated from the root of Stephania tetrandra and used in traditional Chinese medicine as an antirheumatic, anti-inflammatory, antihypertensive agent. 7 –9
Tetrandrine has been demonstrated in studies to have anti-inflammatory, immunosuppressive, and cytoprotective effects. 10,11 It suppresses T and B cells and inhibits the production of inflammatory factors and cytokines such as histamine, prostaglandins, and tumor necrosis factor-α (TNF-α). 12,13 Tetrandrine has been used for treatment of silicosis and can suppress inflammatory reactions associated with rheumatoid arthritis and uveitis or hepatitis in animals, which are provoked by bovine serum albumin, interleukin-1, or endotoxin. 9,12 Recently, several studies have shown that tetrandrine possesses antiproliferative activity, induces apoptosis of malignant cells, and enhances radiation sensitivity in cultured tumor cells. 14 –16 Inhibition of tumor cell proliferation and induction of apoptosis in several carcinomas, including breast and lung cancer, Burkitt's lymphoma, leukemia, malignant lymphoid and myeloid cells, hepatoma/hepatoblastoma, and HeLa cells, have been reported. 17 –20 Further, antitumor effects of tetrandrine and combined radiation to enhance radiation sensitivity on nasopharyngeal carcinoma cell line CNE, ceuroblastoma cells, and gliomas have been recently reported. 21,22 However, the molecular mechanisms of the antitumor effects and enhancing radiosensitivity of tetrandrine remain poorly understood.
Based on these reports, the present study hypothesized that the radiosensitivity of human esophageal cancer cell line TE1 could be enhanced by tetrandrine. In the present study, the effects of tetrandrine on human esophageal cancer cell line TE1 were tested and its potential mechanism of enhancing radiosensitivity was investigated.
Materials and Methods
Reagents and equipments
Tetrandrine was purchased from Sunshine Pharmaceutical Co. Ltd. RPMI-1640 medium (Sigma), fetal bovine serum (FBS; Sijiqing Ltd.), western blot kit and β-actin (Beyotime), cyclin B1 monoclonal antibody (Cell Signaling), ChemMateM EnVision secondary antibody (Dako), electrophoresis system (Bio-Rad), and a linear accelerator resource of X-ray (Siemens) were used in this study. The dose rate was 100 cGy/min and the distance to radiation source was 100 cm.
Cell line and cell culture
Human esophageal cancer cell line TE1 was purchased from the Institute of Medicinal Biotechnology. Cells were cultured under a humidified atmosphere containing 5% CO2 in RPMI-1640 medium supplemented with heat-inactivated FBS (10% by volume), penicillin G (50 U/mL), and streptomycin (50 μg/mL). The cell line was maintained at exponential growth phase and provided fresh medium every 2–3 days.
Preparation of tetrandrine
Tetrandrine was dispersed into a fine suspension in 0.1 N HCl at a concentration of 5 mg/mL. The preparation was sonicated and adjusted to pH 6.6–6.8 with sodium hydroxide. It was then passed through a 0.22-μm filter for bacteriological sterilization and preserved at −20°C for future use. The tetrandrine solution was diluted with RPMI-1640 medium before use.
Methyl thiazolyl tetrazolium assay for cell proliferation
Sensitivity of the TE1 cells to tetrandrine was determined in vitro by methyl thiazolyl tetrazolium (MTT) assay. Briefly, cells were seeded in 96-well plates at a density of 4.0×103 cells/well and allowed to adhere for 24 hours at 37°C. Cells were cultured in the presence of tetrandrine (with concentration at 0, 0.01, 0.1, 1.0, 5.0, and 10.0 μg/mL) for 20 or 44 hours, and then MTT was added to each well. Following a 4-hour incubation at 37°C, dimethylsulfoxide (DMSO) was added to dissolve the dark blue crystal product. The absorbance was measured at a wavelength of 570/630 nm using a microplate reader.
Colony-forming assay for cells survival rate
The TE1 cells in exponential growth phase were trypsinized with 0.25% trypsin and a single-cell suspension of 4×104 per mL was obtained. The cells were incubated in dishes at different densities (5×102, 1×103, 2×103, 5×103, 1×104, and 2×104 per mL) for 24 hours. Thirty minutes before radiation treatment, the cells were supplemented with tetrandrine working solution at different concentrations (0, 0.05, 0.1, and 0.5 μg/mL). Then these cells were treated at different irradiation doses (linear accelerator; Siemens; irradiation doses of 0, 2, 4, 6, 8, and 10 Gy). Twenty-four hours after radiation, the cells were washed free of drug with phosphate-buffered saline (PBS) and incubated in fresh, drug-free medium (10% FBS in RPMI-1640) for 12 days. Subsequently, the cells were washed with PBS, fixed with methanol, and stained with Giemsa solution. The number of colonies containing at least 50 cells was counted under a microscope. Then, colony-forming efficiency (number of colonies formed/number of cells plated) was determined. Graphpad prism 5 was employed for analysis of results. A multitarget click model [S=1 − (1 − e −D/Do) N ] was applied to delineate the survival curve. Radiobiological parameters, including mean lethal dose (D 0), quasi-threshold dose (D q), and survival fraction under X-ray of 2 Gy (SF2), were then calculated.
Flow cytometry analysis of cell cycle
A single-cell suspension was obtained from the cells in the exponential growth phase. These cells were seeded in a six-well plate and incubated for 24 hours. A cell cycle synchronization was achieved by incubation with serum-free RPMI-1640 medium for 12 hours. Thirty minutes before radiation, cells were treated with tetrandrine (5 μg/mL) and irradiated at a dose of 5 Gy. Subsequently, the cells were trypsinized at 12 hours after irradiation. Cells from each group were harvested and washed with PBS. Cold ethanol (70%) was added and stored at −20°C overnight. Cells were stained with propidium iodide (Sigma; 10 μg/mL) treated with RNase A (5 μg/mL) and subjected to flow cytometry (FACSCalibur) for cell cycle analysis.
Western blot analysis for cyclin B1 expression
The cells in the exponential growth phase were trypsinized and incubated for 24 hours and then treated at a dose of 10 Gy. Thirty minutes before irradiation, the cells were treated with 5.0 μg/mL tetrandrine. Also, a control group, with treatment by DMSO, was designed. Subsequently, the cells were cultured for 0, 12, or 24 hours after irradiation. Then the cells were washed twice with ice-cold PBS, scraped in ice-cold PBS, and centrifuged at 14,000 rpm for 10 minutes at 4°C. The cell lysate was collected and used as the cell protein extract. Protein concentration was measured using a protein assay kit according to the manufacturer's instructions. A total of 50 μg of proteins was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to nitrocellulose membrane, and blocked overnight with PBS containing 0.1% Tween 20 and 5% milk. Then, the membrane was incubated with primary antibodies (β-actin or cyclin B1 monoclonal antibody; 1:1000) for 2 hours at room temperature and then washed with PBS containing 0.1% Tween 20 for five times. Subsequently, the membrane was treated with anti-mouse IgG horseradish peroxidase–conjugated secondary antibody (1:1000) for 1 hour at room temperature and then washed with PBS containing 0.1% Tween 20. Color development was performed with an ECL kit (Beyotime). Representative data were from an individual experiment repeated at least three times.
Statistical analysis
Data are expressed as mean±SD. Statistical software SPSS version 13.0 (SPSS) was used to carry out one-way analysis of variance followed by a Student–Newman–Keuls test. The criterion of statistical difference was taken as p<0.05.
Results
Tetrandrine suppressive effect on TE1 cell proliferation
The antiproliferative effect of various concentrations of tetrandrine on TE1 cells is shown in Figure 1. It was observed by MTT assays that tetrandrine inhibited cell growth in a concentration-dependent manner. Compared with the control, 24 hours after tetrandrine treatment (with concentration of tetrandrine 1.0, 5.0, and 10.0 μg/mL), the proliferation of TE1 cells was significantly decreased (p<0.05; F=3.09, 10.43, and 24.00, respectively). Forty-eight hours after tetrandrine treatment (with concentration of tetrandrine 0.1, 1.0, 5.0, and 10.0 μg/mL), proliferation of TE1 cells was more significantly decreased than that at 24 hours (p<0.01; F=4.12, 12.77, 44.28, and 48.53, respectively). The cell viability was calculated using the following formula: Cell viability (%)=(Absorbance treatment/Absorbance control)×100%.
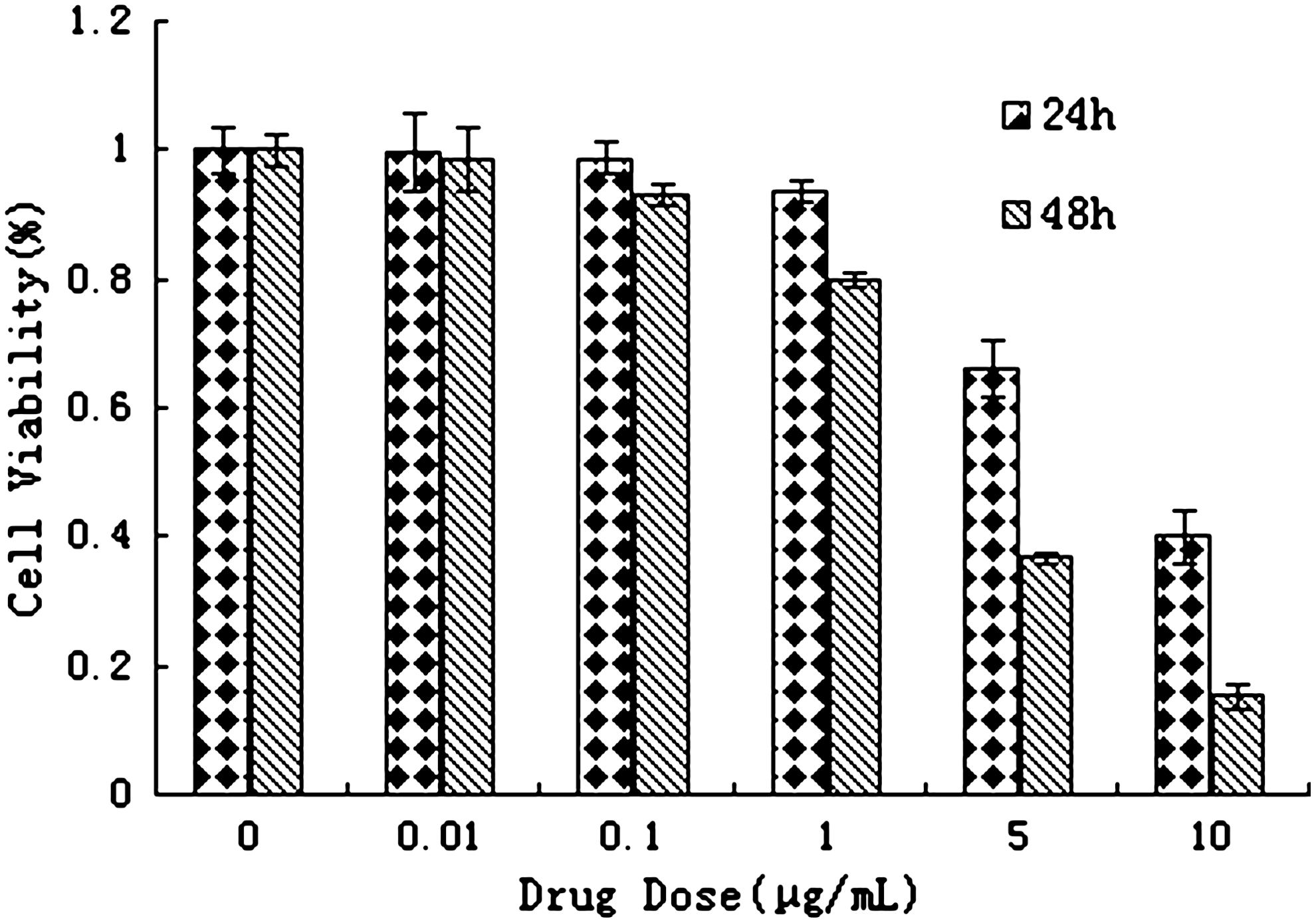
Tetrandrine effect on TE1 cell proliferation. Suppressive effect on TE1 cells was determined by methyl thiazolyl tetrazolium assay on cells continuously incubated in tetrandrine for 24 and 48 hours. Bars: SE of three independent experiments in triplicate. As the drug concentration increased, the growth inhibition ratio of TE1 cells increased. The image represents results from three independent experiments. TE1 cells, esophageal cancer cells.
Synergistic effects of combined tetrandrine and radiation on the TE1 cell proliferation
The proliferation of TE1 cells was gradually suppressed with increasing treatment of tetrandrine and radiation. Fewer colonies were formed by cells that were treated with combined tetrandrine (0.5 μg/mL) and radiation (10 Gy). Figure 2 displays the survival curve with different treatments. The survival rate of TE1 cells was decreased when the irradiation dose increased. The dose–response curve was characterized by the shoulder area under the low-dose treatment and the linear area under the high-dose treatment, which represents multitarget click model.
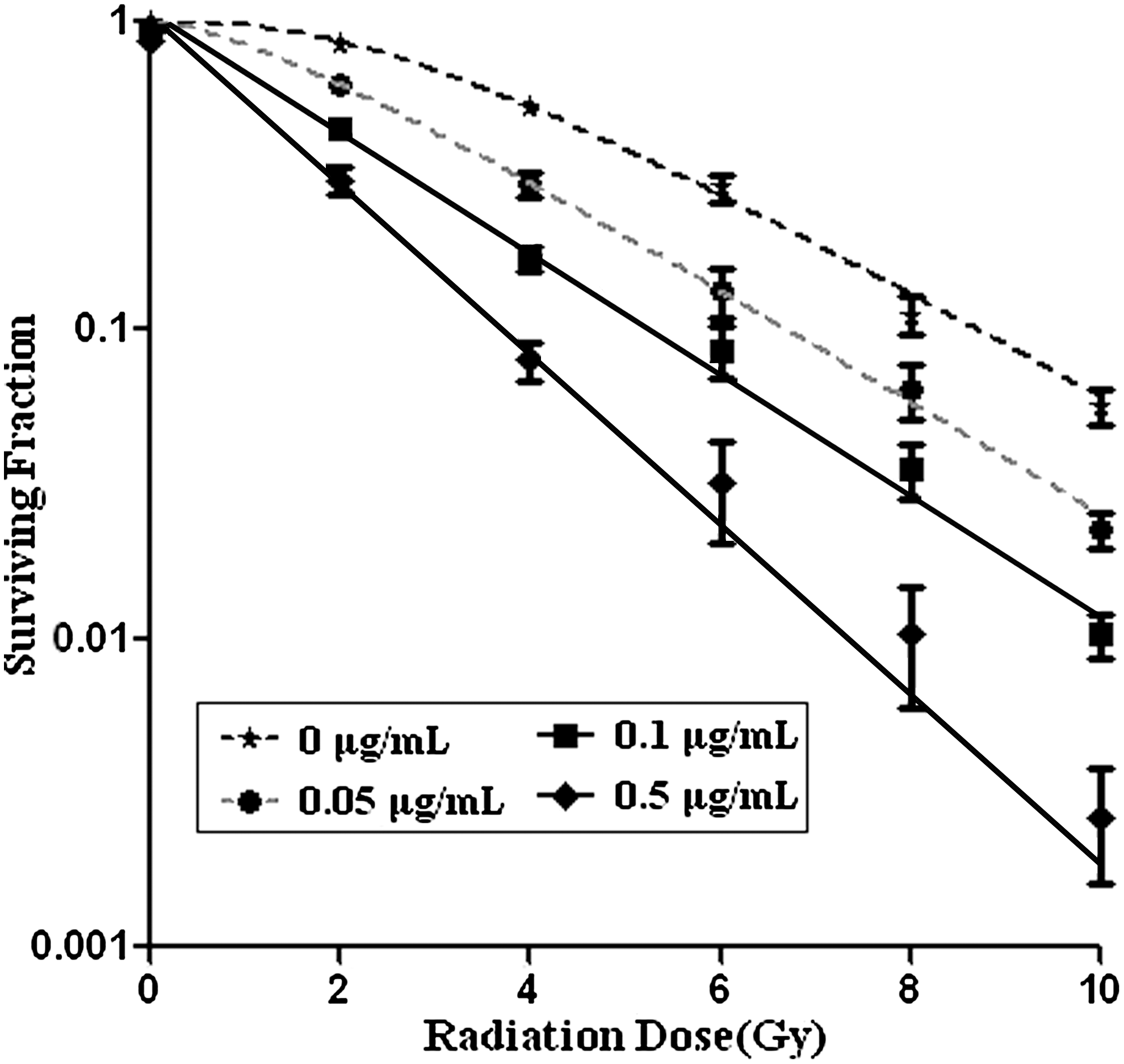
CFE of TE1 cells after treatment with tetrandrine and irradiation. Synergistic effects of combined treatment of tetrandrine and radiation on the CFE of TE1 cells were observed. After treatment with tetrandrine (0, 0.05, 0.1, 0.5 μg/mL) and radiation (0, 2, 4, 6, 8, 10 Gy), the survival rate of TE1 cells was decreased when the irradiation dose increased. Bars: SE of three independent experiments in triplicate. Fewer colonies were formed in which cells were treated with combined tetrandrine at 0.5 μg/mL and radiation at 10 Gy. CFE, colony-forming efficiency.
As shown in Table 1, the radiobiological parameters D 0, D q, and SF2 decreased with the elevation of the tetrandrine concentration. The sensitizing enhancement ratio of tetrandrine (0.5 μg/mL) was 1.62 and 16.71, respectively, when irradiation dose was at D 0 and D q.
Values are given as mean±SD; n=3; p>0.05 versus 0 μg/mL group, (1) F=1.427, 2.481; p<0.05, (2) F=3.25–27.30. Biological parameters of radiation were determined by multitarget click model.
Sensitizing enhancement ratio SERD0 (SERDq)=[D 0 (D q)]control/[D 0 (D q)] treatment.
D 0, mean lethal dose; D q, quasi-threshold dose; SF2, survival fraction under X-ray of 2 Gy; SER, sensitizing enhancement ratio.
Effects of combined tetrandrine and radiation on cell cycle
To explore the relationship between the tetrandrine radiosensitization effect and tetrandrine-induced cell cycle changes, the cells were treated with tetrandrine and irradiation and cell cycle was determined by flow cytometry (Fig. 3). Compared with the control group, the G2/M arrest was significantly increased by irradiation (27.82%:20.02%, p<0.01). The number of TE1 cells with G2/M arrest was dramatically decreased after combined treatment with tetrandrine and irradiation compared with that with tetrandrine treatment alone (22.47%:27.82%, p<0.01).

Cell cycle distribution of TE1 cells at 12 hours after tetrandrine treatment (5 μg/mL). The G2/M arrest was determined by flow cytometry.
Effects of tetrandrine on cyclin B1 expression of TE1 cell
Western blot assay indicated that, after treatment with tetrandrine at 5 μg/mL or irradiation at 10 Gy alone, the expression of cyclin B1 was increased to different extent at different time points (0, 12, and 24 hours). Compared with irradiation treatment alone, the expression of cyclin B1 was markedly increased after combined treatment with tetrandrine and irradiation. No significant changes in the expression of β-actin, an internal reference, were observed (Fig. 4).
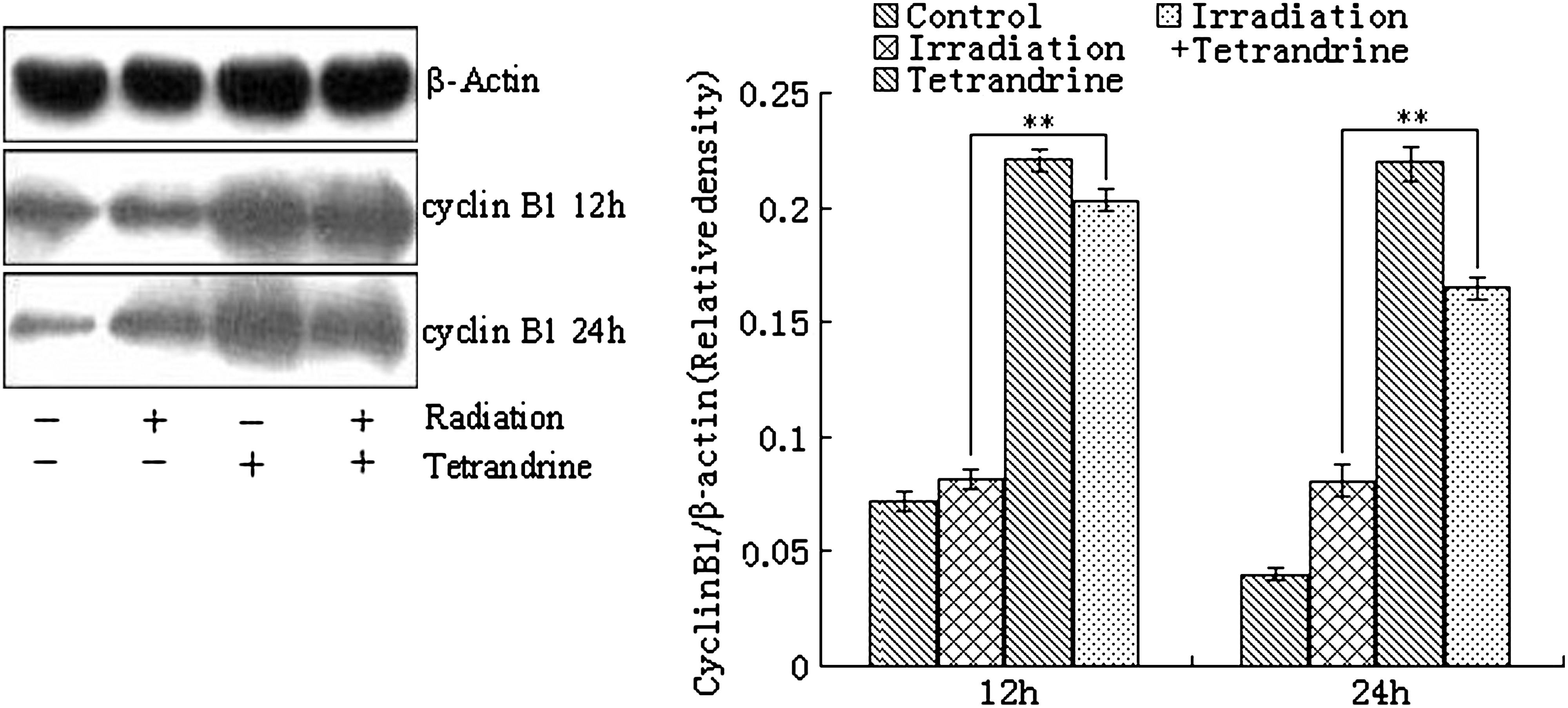
Cyclin B1 expression in TE1 cells after combined treatment with tetrandrine and irradiation. Western blot assay indicated the ability of tetrandrine upregulation of cyclin B1 expression. Twelve or 24 hours after treatment, the cyclin B1 expression dramatically increased in the cells treated with tetrandrine at 5 μg/mL alone or combined with tetrandrine at 5 μg/mL and irradiation at 10 Gy. Band intensity was analyzed by densitometry using the program Bandscan5.0. Results are shown as the mean±SD of three independent experiments conducted in triplicate. **p<0.005.
Discussion
The enhancement effect of radiosensitivity by tetrandrine on human esophageal cancer cell line TE1 has not been previously studied. The present study aimed to investigate the effects of tetrandrine on the radiosensitivity of TE1 cells and its potential mechanism. The results indicated that significant concentration- and time-dependent tetrandrine cytotoxicity was demonstrated for human esophageal cancer cell line TE1. The dose-enhancement ratio of combined treatment with tetrandrine and radiation (1.62 and 16.71, respectively, when dose of irradiation at D 0 and D q) was markedly increased than that of tetrandrine treatment alone. Further, it was shown that the expression of cell cycle protein cyclin B1 was markedly increased after combined treatment with tetrandrine and irradiation than that of treatment with tetrandrine or irradiation alone.
Previous studies reported that tetrandrine exerts antifibrotic effects in DMN-induced fibrotic rats. Tetrandrine also inhibited fibrogenic responses of HSC-T6 cells to TGF-β1 and TNF-α. 23 Other authors observed that administration of tetrandrine in bile duct-ligated rats can reduce the hepatic hydroxyproline content, AST, ALT, and alkaline phosphatase level. 11 Tetrandrine exerts antitumor effects, possibly by suppression of cell proliferation, 12,24 –26 angiogenesis, 25 and multidrug resistance to other anticancer drugs and promotion of apoptosis and radiation sensitivity. 17,18,20,25 Chen et al. found that tetrandrine has cytotoxic effects and elicits a concentration- and time-dependent inhibition of glioma cell proliferation, with an LC50 in the micromolar range. 22 This LC50 value is comparable with previously reported LC50 values for tetrandrine against breast cancer, lung cancer, neuroblastoma, Burkitt's lymphoma, hepatoma, and leukemia. 16 –20,27,28 Recently, tetrandrine has been shown to induce apoptosis in HSC 29 and nasopharyngeal carcinoma cell line CNE. 21 There are no previous reports on the effects of tetrandrine on human esophageal cancer cell proliferation.
To the best of the present study authors' knowledge, few of studies have shown the radiosensitization effect of tetrandrine. 21,27,28 Chen et al. found that tetrandrine had a time- and concentration-dependent cytotoxic effect. Dose-enhancement ratio for RT-Tet and Tet-RT was increased, and it was higher for RT-Tet. 27 The research for mechanism revealed that irradiation caused accumulation in the G2/M phase of the cell cycle, whereas tetrandrine caused G0/G1 accumulation. Compared with irradiation alone, combined irradiation and tetrandrine increased the G0/G1 fraction and decreased the G2/M accumulation, whereas combined tetrandrine and irradiation led to no reduction in radiation-induced G2/M accumulation. Sun et al. 21 tested tetrandrine-induced apoptosis and radiosensitivity in nasopharyngeal carcinoma cell line CNE and investigated the possible mechanisms. They found that tetrandrine could induce cell apoptosis. Further, it was shown that the level of Bcl-2 mRNA decreased and Bax mRNA increased after addition of tetrandrine by using reverse transcription–polymerase chain reaction. Irradiation-induced G2 arrest was abrogated by treatment with tetrandrine, as detected by flow cytometry and mitotic index. The accumulation of cyclin B1 protein and the suppression of Cdc2 tyrosine-15 and Cdc25C serine-216 phosphorylation were detected in irradiated cells treated with tetrandrine using western blot analysis. Taken together, these results showed that tetrandrine can induce apoptosis and abrogate irradiation-induced G2 arrest in CNE cells.
The present study investigated the enhancement effects of radiosensitivity and its potential mechanism by tetrandrine on the human esophageal cancer cell line TE1. The results demonstrate that tetrandrine inhibits the growth of human esophageal cancer cells and efficiently enhances the cytotoxic effect of irradiation. The mechanism by which tetrandrine enhances the cytotoxic effect of irradiation may be by increasing the expression of cyclin B1 protein, abrogating radiation-induced G2 phase arrest, and promoting TE1 cells from the G2 phase into the M phase. These results are comparable with those previously reported. 21,27,28
In summary, the present study data suggest that tetrandrine has a great potential in the treatment esophageal cancer, and more benefit could be obtained by combined treatment of tetrandrine and irradiation. However, these results are preliminary. The optimal schedule of combining RT with tetrandrine has to be yet determined. Further studies for mechanism are necessary. In vivo studies are needed before making any recommendation about the use of tetrandrine in the treatment of esophageal cancer.
Footnotes
Acknowledgments
This work was supported by grants from the Training Program of Young Scientists of Changzhou (CQ2008006) and the Guiding Project for Social Development of Changzhou (CS2008920).
Disclosure Statement
The authors declare that they have no competing interests.