
Abstract
Urinary trypsin inhibitor (UTI) ulinastatin as a broad-spectrum protease inhibitor has been widely used to treat acute pancreatitis and shock and to improve the surgical outcome in the clinic. In the present study, we investigated the potential antihuman breast cancer effects of UTI and its combination with taxotere (TXT). Human primary breast cancer cells and breast cancer cell line MDA-MB-231 cells were treated with UTI with or without TXT, and invasion and metastasis ability of these cells were evaluated, respectively, by a transwell assay. Reverse transcription–polymerase chain reaction was used to detect fibroblast growth factor, vascular endothelial growth factor c, epidermal growth factor, epidermal growth factor receptor, transforming growth factor-β1, and protein kinase B/AKT. We also investigated the in vivo role of UTI by using a xenograft mouse model, and immunohistochemical assay was employed to show the expression of factors involved in either angiogenesis or the epithelial–mesenchymal transition (EMT). Our results showed that UTI inhibited invasion and metastasis in both primary and MDA-MB-231 cells both in vivo and in vitro. Especially, UTI presented the significant combined effects with TXT on these cells in terms of angiogenesis blocking and EMT inhibition. These results suggest that UTI and its combination with TXT present therapeutic potential against breast cancer and deserve further preclinical and clinical studies.
Introduction
Breast cancer is one of the most common malignancies among women. The incidence rate of breast cancer is increasing yearly, whereas the age of onset is decreasing. 1 Current treatments for breast cancer are mainly surgery and chemotherapy. However, the invasion and metastasis of tumor cells usually lead to poor prognosis for many patients. Penetration into the extracellular basement membrane is the premise of cancer cell metastasis, where a variety factors, including many proteases, play essential roles.
Epithelial–mesenchymal transition (EMT) refers to a biological process when the epithelial cells change into cells with mesenchymal phenotypes through complex mechanisms. 2 During EMT, the epithelial cells lose cell polarity and connections with the basement membrane that are characteristics of an epithelial cell phenotype and obtain a high invasion/metastasis potential, with the ability to degrade the extracellular matrix and assume mesenchymal phenotypes that are considered to be closely related with initiation and development of cancer. 3
Urinary trypsin inhibitor (ulinastatin, UTI) as an inhibitor of proteases can inhibit a broad-spectrum of proteases, including trypsin, chymotrypsin, plasmin, human leukocyte elastase, and hyaluronidase. It has antitumor metastasis and protective effects on patients treated with radiotherapy and chemotherapy, and has been widely used to treat acute pancreatitis and shock and to improve surgical outcome in the clinic.
Our previous studies demonstrated that UTI significantly inhibited the proliferation of the human breast cancer cell line MCF-7 in vitro and the growth of xenografts in nude mice in vivo by decreasing the expression of CXCR4 and MMP-9. In addition, UTI could enhance the antitumor effects of cyclophosphamide and docetaxel. 4,5
To find a more effective treatment for breast cancer, we intended to examine the additive effects of UTI and taxotere (TXT) on the proliferation of breast cancer MDA-MB-231 cells and tumor growth in nude mice as well. We have investigated the effects of UTI with or without TXT on the invasion and metastasis of breast cancer cells in vitro and xenografts in nude mice in vivo. Our results show that UTI and its combination with TXT can block both the invasion and metastasis of breast cancer cells by inhibiting the events of angiogenesis and the EMT.
Materials and Methods
Cell lines and animals
The human breast cancer cell line, MDA-MB-231 (ER−), was provided by the Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. Female BALB/c-nu/nu nude mice, aged 4–6 weeks old and weighing 17–21 g, were purchased from the Animal Research Center of the Chongqing Medical University.
Reagents and instruments
UTI (ulinastatin) was purchased from Techpool Biopharma Co., Ltd., and TXT was purchased from Sanofi-Aventis. The reverse transcription–polymerase chain reaction (RT-PCR) kit was purchased from Takara Shuzo Co., Ltd. Antibodies for vascular endothelial growth factor c (VEGF-c), basic fibroblast growth factor (bFGF), PAFR, epidermal growth factor (EGF), epidermal growth factor receptor (EGFR), transforming growth factor-β1 (TGF-β1), CD44, AKT, NM23, CK19, and S100A were purchased from Abcam. Mouse anti-human breast cancer-associated antigen CA15-3 monoclonal antibody was obtained from Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd. The cytoplasm of breast cancer cells may show CA15-3-positive staining. The FACSVantage™ SE Flow Cytometry System was from Becton Dickinson. The Gel Doc XR Image System was from Bio-Rad. The CX40 fluorescence microscope was from Olympus Corporation.
Cell culture and nude mouse breeding
A breast cancer sample was obtained from a female patient from the Department of Breast and Thyroid Surgery at the Second Affiliated Hospital of the Chongqing Medical University. The 72-year-old patient had not received any chemotherapeutic drugs and had no history of other major diseases. The results of pathological examination demonstrated that the tumor was invasive ductal carcinoma, and immunostaining of estrogen receptor (ER), progesterone receptor (PR), and oncoprotein cerB-2 was scored as follows: ER (+), PR (++), and CerbB-2 (−). The sample was obtained by radical mastectomy under sterile conditions and minced into ∼1-mm3 pieces within 2 hours after surgery. The small pieces of tissue were digested with 1% type 2 collagenase and hyaluronidase for 12 hours at 37°C. The supernatant was discarded, and the breast cancer cells were collected after centrifugation at 1000 rpm for 5 minutes. The concentration of cells was adjusted to 105 per milliliter with RPMI 1640+10% fetal bovine serum (FBS). Trypan blue staining was used to determine cell viability. Breast cancer cells were subcultured repeatedly to remove impurities and nonadherent cells. The human breast cancer cell line MDA-MB-231 was cultured in RPMI-1640 containing 10% FBS, 100 U/mL penicillin, and 100 mg/L streptomycin at 37°C, 5% CO2 in a humidified incubator. Cells in the logarithmic growth phase were trypsinized and used for following experiments. Mice were housed under specific pathogen-free conditions, with temperature ranging from 22°C to 25°C, and the humidity was 50%–65%. All the manipulations were performed under sterile conditions.
Immunofluorescence staining
Breast cancer cells were seeded at a concentration of 1×105 per well and allowed to grow on the coverslip to 80% confluency. The sections were fixed, permeabilized with 0.1% Triton X-100, washed with phosphate-buffered saline (PBS), and then exposed to anti-CA15-3 antibody (1:100) followed by fluorescein isothiocyanate-labeled secondary antibody (1:100). The slides were viewed at a magnification of×400 using an Olympus CX40 fluorescence microscope.
Drug administration
For cell-based assays, the four groups included the control group, the UTI group, the TXT group, and the UTI+TXT group. The concentration of UTI was 800 U/mL 3 and TXT was 3.7 μg/mL. All drugs were reconstituted <6 hours before administration. Control wells were treated with an equal volume of saline.
Animal experiment
The breast cancer cells were trypsinized, rinsed with PBS, and resuspended with a serum-free RPMI-1640 medium to the density of 1×107 cells per milliliter. The cell suspension in 0.2 mL was inoculated into the right fat pad of nude mice. In week 3 after inoculation, 28 of 45 mice with tumor volumes of ≥500 mm3 were randomly divided into the following four groups: UTI—7 mice received 1600 U per day of UTI for 20 consecutive days; TXT—7 mice were injected with 20 mg/kg TXT on days 1, 7, and 144; UTI+TXT—7 mice were treated with both UTI and TXT as mentioned above; and control—7 mice were injected with an equal volume of saline for 20 days. Three weeks after drug administration, all mice were sacrificed to collect specimens. Tumor volumes were calculated as 1/2×(L 2×D), where L (cm) was the maximum diameter of the tumor, and D (cm) was the minimum diameter. Tumor inhibition rates were calculated using the formula, 1−(ending tumor volume of the treatment group−starting tumor volume of the treatment group)/(ending tumor volume of the control group – starting tumor volume of the control group)×100%. q Value was calculated according to Jin's formula, which was used to determine the interaction of UTI and TXT. 6 Jin's formula is q=c/(a+b)−a×b, where a was the inhibition rate of the UTI group; b was the inhibition rate of the TXT group; and c was the inhibition rate of the UTI+TXT group. q>1.15 was considered as synergy; 1.15>q>0.85 was an additive effect; and q<0.85 represented an antagonistic effect.
A xenograft model of MDA-MB-231 was established in a similar manner while the cells were inoculated subcutaneously into the right armpits of the nude mice.
Cell invasion assay
Matrigel (300 μg/mL) was spread evenly on the membrane in transwell inserts, and the chamber was kept at 4°C overnight. Matrigel, 100 μL of a 2 mg/mL solution, was added into each upper chamber of a 24-well transwell plate, and the plate was incubated at 37°C for 8 hours; then, each well was washed with a serum-free medium for 30 minutes at 37°C. The primary breast cancer cells that were cultured for 24 hours in a serum-free medium were trypsinized and centrifuged. Cell pellets were resuspended with a medium containing 0.1% FBS to 2×104 cells per milliliter. Cells (100 μL) treated with/without drugs were added to the upper chamber of the transwell, and 200 μL medium containing 0.1% FBS+200 μL medium containing 10% FBS was added to the lower chamber. The plates were incubated at 37°C in a 5% CO2 incubator for 24 hours. Each well was washed twice with PBS, and the inner surface of the upper chamber was carefully wiped using a cotton swab. Invaded cells were fixed with 95% ethanol, stained with 4 g/L Trypan blue for 10 minutes, observed under the microscope, counted, and photographed. The inhibitory rate of invasion was calculated as follows: percent of inhibition=(1−invasion cell number of the experimental group/control invasive cell number) ×100.
Assessment of mRNA expression using RT-PCR
Cells were cultured in an RPMI-1640 medium containing 10% FBS in ∼5×105 per milliliter. Forty-eight (48) hours after the administration of the drugs, an RNAiso Plus Kit was used to extract the total RNA. Reverse transcription was performed with a Reverse Transcription Kit according to the manufacturer's instructions, following the assessment of the purity and concentration of the extract. The complementary DNA synthesized using RNA as the template was analyzed immediately and was used as a template for real-time PCR. β-Actin and GAPDH were employed as internal reference controls. All primers were designed by Primer premier 5.0 software and synthesized by the Sangon Biotech (Shanghai) Co., Ltd. Primers were as follows: forward primer of VEGF-c: 5′-GGAGCAGTTACGGTCTGTGT-3′, reverse primer: 5′-GTCCTTGAGTTGAGGTTGGC-3′; forward primer of bFGF: 5′-TCAAACCCTTCTCTGTACCC-3′, reverse primer: 5′-AAGCCATATTCATTCACCAT-3′; PAFR forward primer: 5′-AACGTCACTCGCTGCTTTGAG-3′, reverse primer: 5′-CACAGTTGGTGCTAAGGAGGC-3′; EGF forward primer: 5′-ATGAATGTGAGATGGGTGTC-3′, reverse primer: 5′-TCTTACGGAATAGTGGTGGT-3′; EGFR forward primer: 5′-CCAAGGCACGAGTAACAAGC-3′, reverse primer 5′-AGGGCAATGAGGACATAACCAG-3′; TGF-β1 forward primer 5′-CCGCAACAACGCAATCTATG-3′, reverse primer: 5′-GTCAGCAGCCGGTTACCAAG-3′; AKT forward primer: 5′-TCTTCCACCTGTCTCGTGAGC-3′, reverse primer 5′-AGCCCGAAGTCCGTTATCTT-3′; GAPDH forward primer 5′-ATCAACGGGAAACCCATCAC-3′, reverse primer 5′-CGCCAGTAGACTCCACGACAT-3′; β-Actin forward primer: 5′-CACCCGCGAGTACAACCTTC-3′, reverse primer: 5′-CCCATACCCACCATCACACC-3′. The PCR conditions consisted of 94°C for 30 seconds, 58°C for 30 seconds, and 72°C 1 minutes for 35 cycles. Five microliters of each amplification product was then subjected to electrophoresis on 1.8% agarose gels in the TAE buffer. Digital images were obtained and analyzed with the Gel Doc-XR System. The expression level of the target gene was evaluated as a ratio of the density of the target gene band to the internal reference gene band.
Immunohistochemistry
After the mice were euthanized, tumor tissues were fixed in formaldehyde, embedded in paraffin, and sectioned. The expression of VEGF-c, bFGF, EGF, EGFR, TGF-β1, CD44, AKT, NM23, CK19, and S100A was determined by an SP immunohistochemical method. Cells with a brown cytoplasm or brown/yellow granules were recognized as positive. Staining images were analyzed with Image Pro-plus 6.0 software (Media Cybernetics, Inc.) for integral optical density.
Statistical analysis
All data are expressed as mean±standard deviation. One-way analysis of variance was used to compare the differences between the groups by using Statistical Package for Social Sciences (SPSS) 18.0 software. p Values <0.05 were considered statistically significant.
Results
Purification of primary breast cancer cells
After serial passages for 2 weeks, the primary breast cancer cells had grown and proliferated stably. Cancer cells were spindle-shaped and adherent, and mingled with a small amount of polygonal-shaped stromal cells and minimal cell debris. The differential adhesion method was repeatedly performed to remove stromal cells and fibroblasts. The cell viability was more than 90% as shown by trypan blue staining. The cytoplasm of breast cancer cells showed CA15-3-positive staining (Fig. 1).
Primary cells stained with anti-CA15-3 (400×).
Inhibition of breast cancer cell invasion by UTI and TXT
Compared with the control group, the invasion of primary breast cancer cells was significantly inhibited in either the UTI or the TXT group; however, the inhibition in the UTI+TXT group was the highest (p<0.05) (Fig. 2A). Similarly, the invasion ability of MDA-MB-231 cells was significantly lower after treatment (p<0.05) (Fig. 2B and Supplementary Table S1; Supplementary Data are available online at
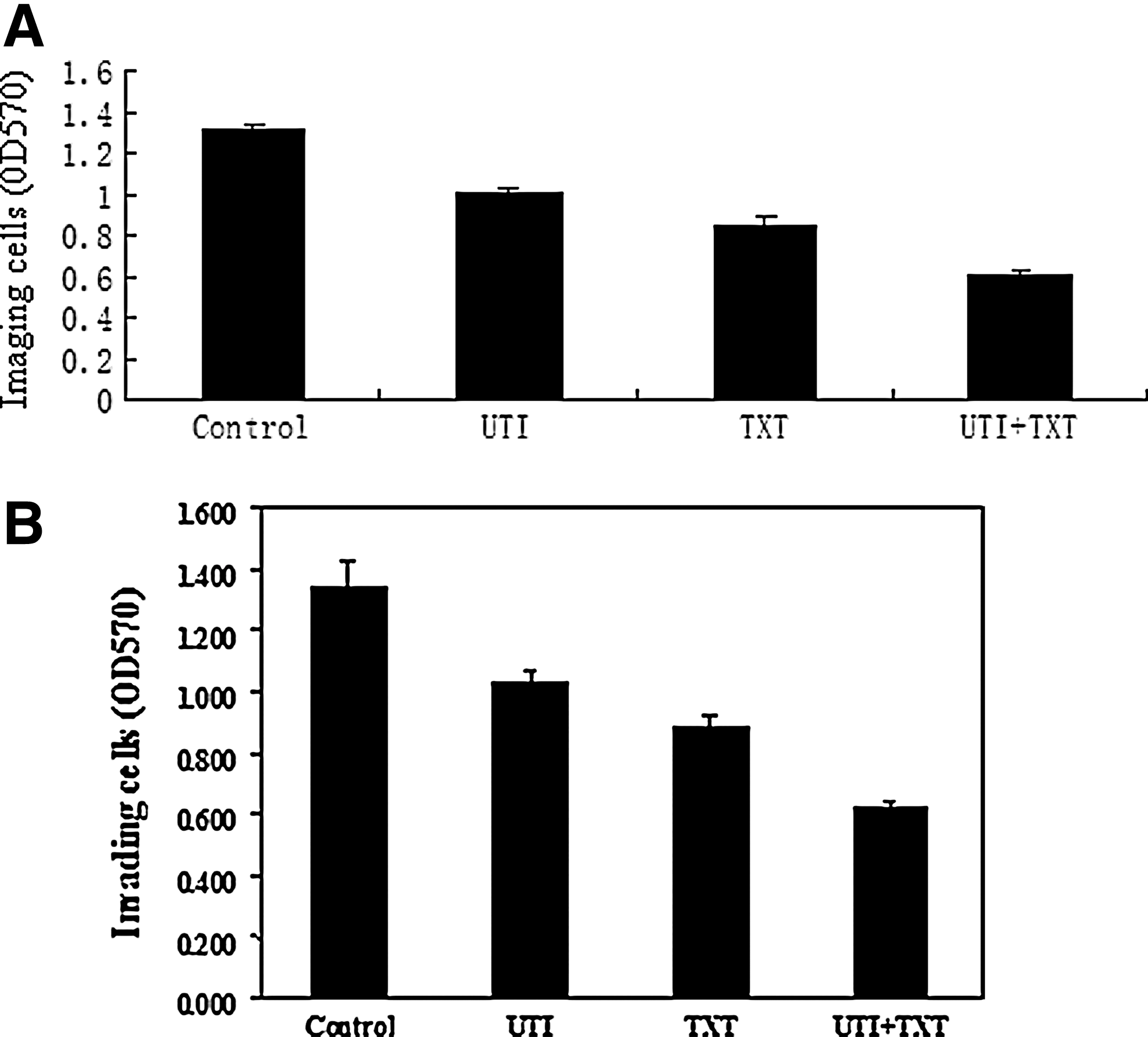
Effect of UTI and/or TXT on invasion ability of the
RT-PCR detection of the gene expression pattern in the breast cancer cells treated by UTI and TXT
To determine the effects of drugs on the gene expression pattern, RT-PCR was performed to determine mRNA levels of relative genes in the breast cancer cells. Compared with the control group, treatment with UTI and/or TXT for 48 hours led to reduced expression levels of the VEGF-c, bFGF, PAFR, EGF, EGFR, TGF-β1, and AKT genes in primary breast cancer cells, which was statistically significant (Fig. 3A–G and Supplementary Table S2). Inhibition of gene expression in the UTI+TXT group was more obvious than in the TXT and UTI groups (p<0.05). UTI, TXT, and UTI+TXT significantly inhibited mRNA expression of VEGF-c and bFGF in MDA-MB-231 cells, compared with the control group (p<0.05) (Fig. 3H, I and Supplementary Table S3).

Effects of UTI and/or TXT on the mRNA expression of
UTI and/or TXT inhibited the growth of breast cancer xenografts in nude mice
Two mice died during the experiment (one from the control group and the other from the UTI group), which may be due to rapid tumor growth, excessive consumption, and cachexia. Compared with the control group, UTI treatment did not significantly affect the tumor volume, but delayed the growth of tumors (p<0.05). Eleven days after drug administration, tumor volumes of the TXT and UTI+TXT groups were diminished in a time-dependent manner, and the inhibition rate of the UTI+TXT group was higher than that of the TXT group (p<0.05). According to Jin's formula, q equaled 1.088 in the UTI+TXT group, suggesting an additive effect of UTI and TXT on inhibition of xenografts in nude mice (Fig. 4A and Supplementary Table S4). Growth curves of MDA-MB-231 xenografts in nude mice were similar with those of the primary breast cancer model (Fig. 4B).

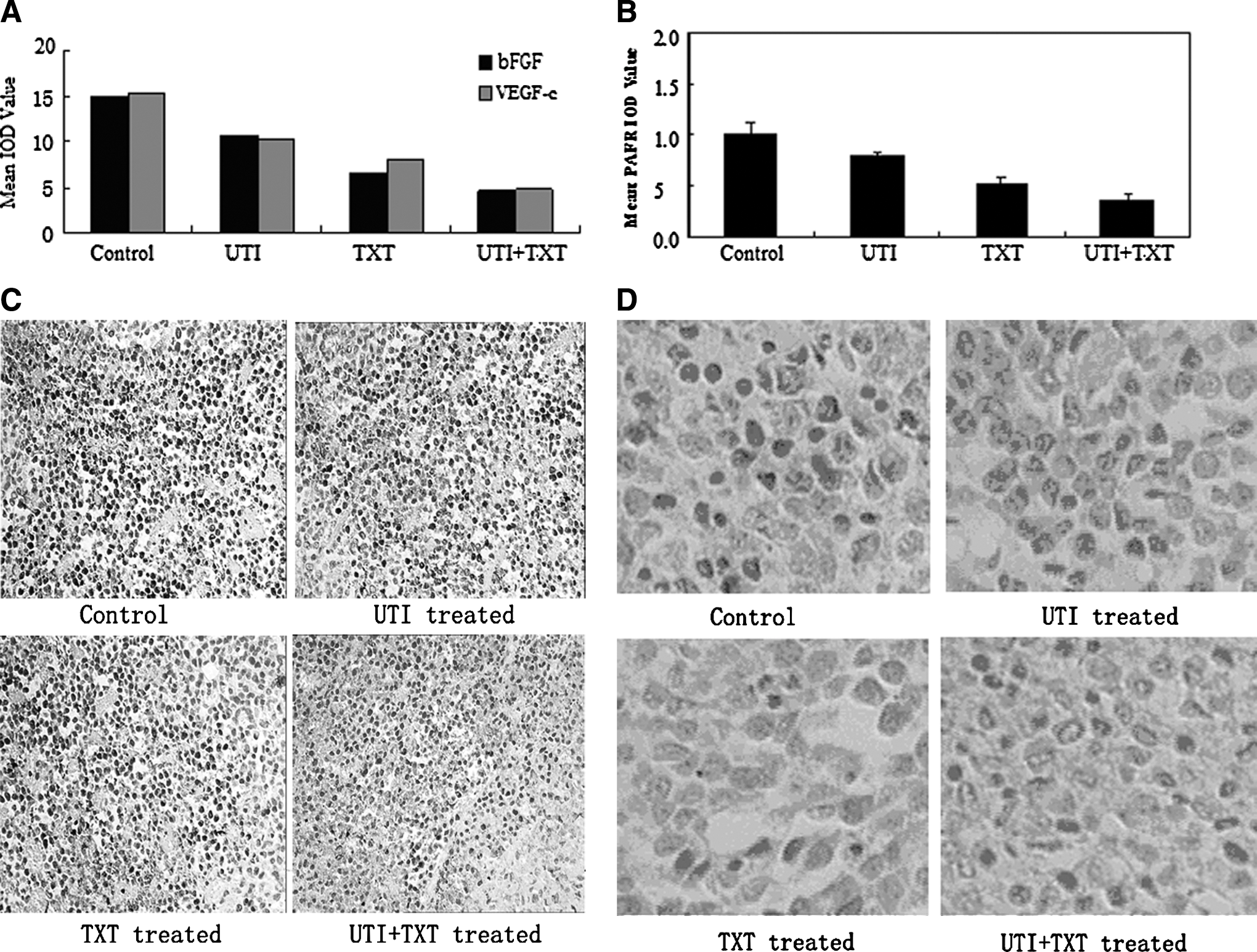
Expression of bFGF, VEGF-c, PAFR, EGFR, TGF-β1, CD44, AKT, CK19, S100A4, and nm23 protein in xenografts
The results of the immunohistochemical assay showed that, compared with control treatment, UTI and/or TXT inhibited the expression of bFGF, VEGF-c, EGF, EGFR, TGF-β1, AKT, CK19, and CD44 in tumors, and the UTI+TXT treatment exhibiting the strongest inhibition of these markers (Fig. 5A, C, Supplementary Tables S5 and S6). The comparison between every two groups was statistically significant (p<0.05). In addition, UTI and/or TXT increased the expression of nm23 protein, and the expression of nm23 was highest in the UTI+TXT group. Every two groups were statistically significant (p<0.05) (Fig. 5B, 5D).
Discussion
Invasion and metastasis have been recognized as the most important biological features of breast cancer. Many extremely complicated regulatory factors have been identified to be involved in the development of breast cancer, which makes the treatment complicated. The goal of the study was to explore the angiogenesis and EMT mechanisms in UTI- and TXT-treated breast cancer cells in regard to invasion and metastasis, and eventually find the corresponding optimal therapeutic measures.
Tumor metastasis is a complex process, and many factors are involved, including oncogenes, tumor suppressors, angiogenesis factors, and EMT factors. nm23 is a metastasis suppressor gene that is engaged in nucleoside triphosphate metabolism in vivo and plays a regulatory role in cell differentiation and signal transduction. In addition, nm23 was a metastatic marker of breast cancer and was valuable in predicting metastatic potential, which is hard to determine by conventional pathology. 1 Furthermore, the nm23 gene was negatively correlated with TNM staging and positively correlated with the survival of patients, and nm23 can serve as a favorable prognostic factor that will provide a more comprehensive assessment of prognosis if it is combined with other cancer indicators. 7
CD44, an adhesion molecule and a member of integrated membrane glycoproteins, is involved in cell–cell interactions, cell adhesion, and migration. It interacts with a variety of molecules in the extracellular matrix, as well as with cytoskeletal proteins involved in the formation of cellular pseudopod and cell migration. 8 CD44 in cancer cells enables the anchoring of the cells to the extracellular matrix and promotes their adhesion and movement, which facilitates the distant metastasis of the cancer cells. 9
VEGF-c is a prolymphangiogenesis factor that is closely related with lymphangiogenesis and the lymph node metastasis of breast cancer. 10 –12 The invasion ability of the breast cancer cell line, MCF-7, is significantly inhibited after knockdown of VEGF-c. 13 In contrast, recombinant human VEGF-c promotes the invasion of the HeLa human cervical cancer cell line in a dose-dependent manner. 14 bFGF is a potent broad-spectrum mitogen that is related to the invasion of breast cancer cells and angiogenesis. 15 MDA-MB-231 cells release FGFs (aFGF and bFGF), which react with tumor cells and promote tumor angiogenesis, in an autocrine manner. 16
PAFR is a G protein-coupled receptor that plays an important role in the adhesion of endothelial cells, promoting metastasis and blood vessel formation. 17 WEB2086 (a PAFR antagonist) inhibits proliferation and invasion of breast cancer cells, which may be related to inhibition of metastasis, angiogenesis, and reduction of microvessel density in tumors. 18 –20
EGFR, encoded by the proto-oncogene c-erbB1, belongs to the human epidermal growth factor receptor (HER) family, with other family members, HER2, HER3, and HER4. 21 The activation of the EGFR-signaling pathway activates ras and raf, which phosphorylate c-fos and c-jun, thereby increasing activity of AP-1. AP-1 translocates to the binding sites in the promoter region of VEGF and IL-8, which induces their translation and stimulates angiogenesis. 22,23
Interestingly, using breast cancer xenografts as a model, we found that the expression of CD44, VEGF-c, PAFR, and EGFR was significantly decreased, whereas nm23 expression was dramatically increased after UTI+TXT treatment, which may partially explain the antitumor metastatic role of UTI+TXT.
TGF-β1 plays a key role during EMT-induced tumorigenesis. 24 TGF-β may increase the number of EGFRs on the cell surface to promote EMT. 25 TGF-β was significantly decreased in breast cancer xenografts after UTI+TXT treatment that could also partially explain the antibreast cancer metastatic role of UTI+TXT.
Taken together, we found that UTI+TXT significantly inhibited the invasion and metastasis of primary breast cancer and MDA-MB-231 cells both in vivo and in vitro. Compared with the control group, the expression of the VEGF-c, EGFR, TGF-β1, and AKT genes in breast cancer cells was decreased in either the UTI or the TXT treatment group and even lower in the UTI+TXT treatment group. Tumor growth was delayed in the UTI treatment group, and the tumor volumes of the TXT treatment group and the UTI+TXT treatment group reduced gradually (p<0.05). Both UTI and TXT inhibited the expression of bFGF, VEGF-c, PAFR, EGFR, AKT, TGF-β1, CK19, and S100A4 in breast cancer xenografts. The combination of both drugs was stronger than either drug alone under the conditions tested, which highlights the synergy effects of the two drugs. We propose that UTI enhances the anticancer effects of TXT through these possible mechanisms as follows: (1) UTI downregulated the expression of EGFR, AKT, and TGF-β1, inhibiting the expression of bFGF and VEGF-c, thus reducing lymphangiogenesis, angiogenesis, and tumor metastasis; (2) UTI controlled invasion and metastasis of tumor cells through downregulating TGF-β1, disrupting the interaction of EGF and EGFR, reducing expression of neural cadherin S100A4, and preventing EMT process; and (3) UTI inhibited metastasis by affecting the migration of cancer cells via the reduction of the TGF-β1, PAFR, and CD44 levels. In conclusion, our findings suggest that UTI could be applied as an adjuvant treatment of the invasiveness and metastasis of breast cancer.
Footnotes
Acknowledgment
This work is supported by a Scientific and technological project in the Chongqing grant (CSTC, 2008AC5082).
Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.