
Abstract
Abstract
To illustrate the control of harmful cyanobacterial growth and the removal of nutritients from fresh water, three submerged macrophytes were grown in the raw water of Guishui Lake. Lindernia rotundifolia, Hygrophila stricta, and Cryptocoryne crispatula were grown together in situ to assess their effectiveness in nutrient removal in microcosms. Results revealed the inhibitory effects of these species on cyanobacterial growth. In addition, water quality in the planted microcosms showed improvement when compared to the water quality of the unplanted microcosm. At all treatments studied, the chemical oxygen demand in the planted microcosms was lower than that in the unplanted microcosms, and the removal rate of all the nitrogen and phosphate in the planted microcosms was better than that of the microcosm without plants. Our study offers a useful algal control method for the lakes or reservoirs that suffer from harmful cyanobacterial blooms.
Introduction
Evidence suggests that the worldwide increase in HABs is linked to increased nutrient loading (Heisler et al., 2008) While the strong relationship between phosphorus loading and HCBs in freshwater ecosystems has been observed for years (Burkholder, 2002; Wilson et al., 2005), more links between nutrient input and the amount of HABs have been discovered recently (Khan and Ansari 2005; Glibert and Burkholder, 2006). Glibert et al. (2005) have noted that over the past several decades the biomass of algae cells has increased with the increase in nitrate loading. However, a statistical correlation does not establish a causal link, and this relationship suggests that eutrophication may be a causative agent of change (Glibert et al., 2006a; Glibert et al., 2006b). Some research has shown that extensive blooms are strongly related to nitrogen-rich agricultural runoff in water. A week after elevated ammonium levels were reported in Washington state inland waterways, an intense bloom of the toxigenic diatom Pseudonitzschia pseudodelicatissima was found (Trainer et al., 2007). Multiple examples suggest that the management of nutrient inputs to the watershed can lead to a significant reduction in HABs.
While it is clear that steps must be taken to significantly reduce the negative impact of high nutrient inputs on bodies of water, the need to maintain required rates of agricultural productivity precludes the possibility of stopping the use of fertilizers altogether. Given this, an ecologically friendly system for reducing inputs and/or treating point and nonpoint source runoffs is desirable. Aquatic submerged species can utilize large amounts of nitrogen (N) and phosphorus (P), thus removing them from the water. Many researchers have investigated the N and P removal capacities of different aquatic plants (Ahern et al., 2008; Fraser et al., 2004; Maltais-Landry et al., 2009). Bindu et al. (2008) have demonstrated that subsurface flow systems planted with Colocasia esculenta can decrease the nitrate and phosphate content of wastewater in addition to removing organic matter. They can also improve the quality of treated water and inhibited blooms (Mjelde and Faafeng, 1997). Wu et al.(2010b) reported that decreases in the levels of nutrients can control cyanobacterial blooms.
In earlier work by our group, three aquatic submerged plants, Lindernia rotundifolia, Hygrophila stricta, and Cryptocoryne crispatula, were selected from 12 plant varieties and removed N and/or P from water effectively (Zhong et al., 2008). In other research, we studied the effect of the input density of plants (plant/water, g/L) on the purifying efficiency of eutrophic lake water (Wang et al., 2009). The result demonstrated the fittest input density of plants, which had the highest purifying efficiency, could be useful to the removal of N and P. Building on our prior work, the purpose of this research is to illustrate the control of harmful cyanobacterial growth (HCG) and the removal of nutritients from the raw water by using submerged macrophytes.
Materials and Methods
Study site
The study site (Fig. 1) was located in Guishui Lake, which can be found in Yanqing County, up river of the Guanting Reservoir, and northwest of Beijing, China. Guishui River runs through the town of Yanqing. The banks of Guishui River are the main agricultural areas of the county.
Study site.
Design of experiment
Three microcosms (0.8 m×0.8 m×1.5 m each) were located in nearshore water of Guishui Lake. The water depth was ∼3 m. Details of the microcosm design are illustrated in Fig. 2. Two microcosms were planted with submerged macrophytes and one (the control) remained without plants. To avoid waves overtopping the microcosms, we buoyed the microcosms above the surface of water 30 cm. Water quality was monitored in these microcosms for 28 days, from July 14, 2008, to August 10, 2008. During the experiment, the average water temperature was 28°C.
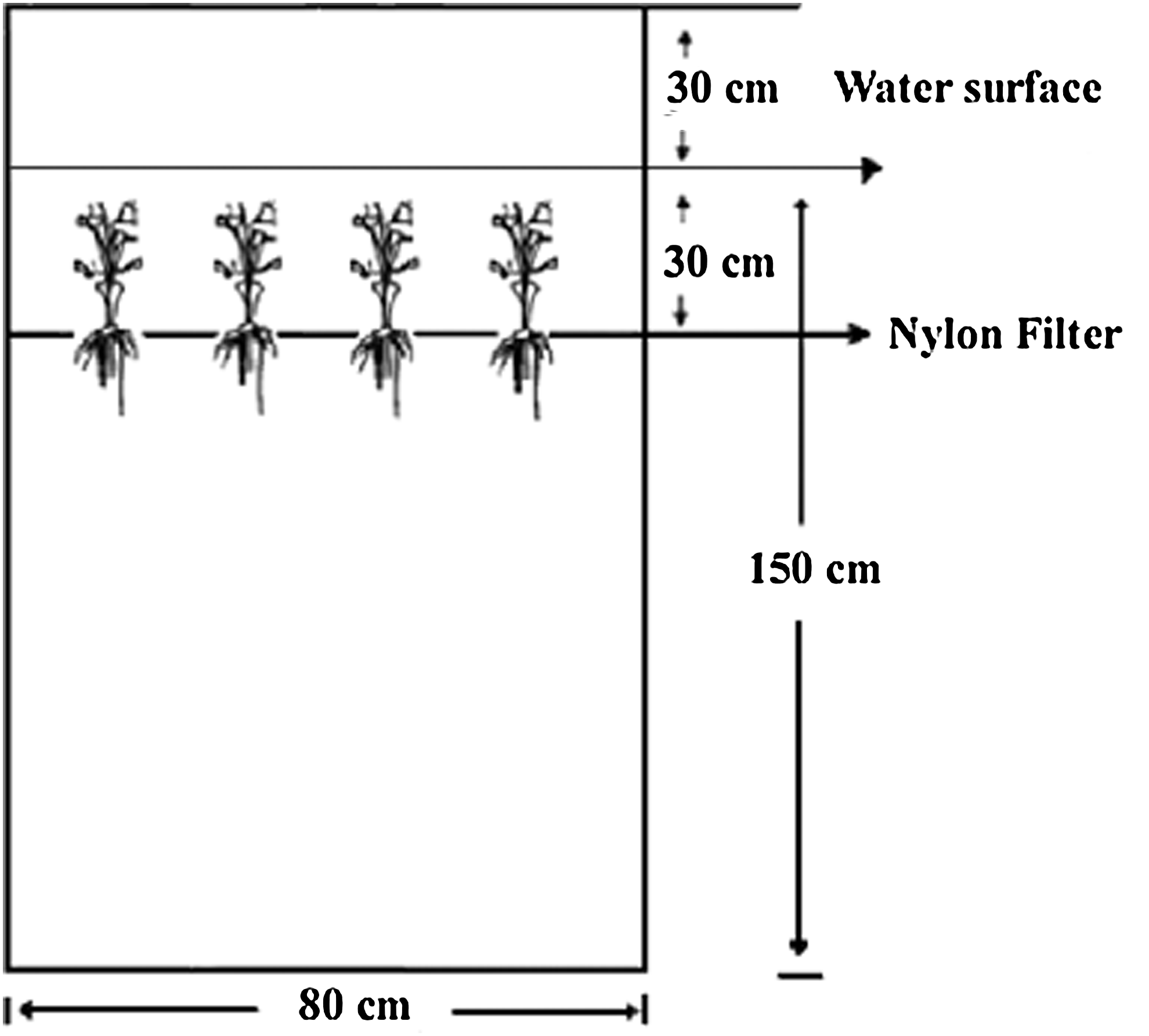
View of the experimental microcosm.
L. rotundifolia, H. stricta, and C. crispatula, (<25 cm in length) were selected for use and then planted according to the method described by Wang et al. (2009). The input density of the plants in each of the three microcosms was 480 g L. rotundifolia, 432 g H. stricta, and 528 g C. crispatula. The submerged macrophytes were kept at 5 cm below the water surface.
Analysis of water
Starting from hour 0, water samples were analyzed periodically for pH, temperature, cell count, chemical oxygen demand (COD), total nitrogen (TN), ammonium nitrogen (NH4+), nitrate nitrogen (NO3−), nitrite nitrogen (NO2−), and dissolved phosphate phosphorus (PO43−). These were determined according to Vaillant et al. (2003). Turbidity was monitored using a spectrophotometer, and the calibration was performed using deionised water and a liquid turbidity standard of 500 FTU (formazin turbidity units).
Results
Reduction of algal biomass and turbidity
The difference between the control microcosm and the microcosms planted with L. rotundifolia, H. stricta, and C. crispatula could be observed by day 6, when an algal bloom appeared in the control microcosm but not in the planted microcosm (see Fig. 3). In addition, the water in the planted microcosm was clearer than that in the control.

Pictures of microcosm experiments at Guishui Lake on day 6.
The variations of algal biomass are shown in Fig. 4. The biomass of single cell cyanobacterial species was counted under a microscope. The algal (cyanobacterial) biomass of the planted microcosm was lower than that of the control. During the first four days, a significant reduction in algal biomass was observed in both the planted and unplanted microcosms, from 1.8×106 cells/mL to 0.6×106 cells/mL and 1.8×106 cells/mL to 0.95×106 cells/mL, respectively. The algal biomass in the planted microcosm decreased consistently until the end of the experiment. From day 6 to day 28, the algal biomass in the planted microcosm contained 0.4×106 cells/mL, with slight fluctuations in number. In contrast, the algal biomass in the control microcosm rapidly increased to 1.5×106 cells/mL from day 4 to day 9, and algal bloom occurred after day 6 (Fig. 4). In the planted microcosm, algal bloom was inhibited during the entire period of the experiment.
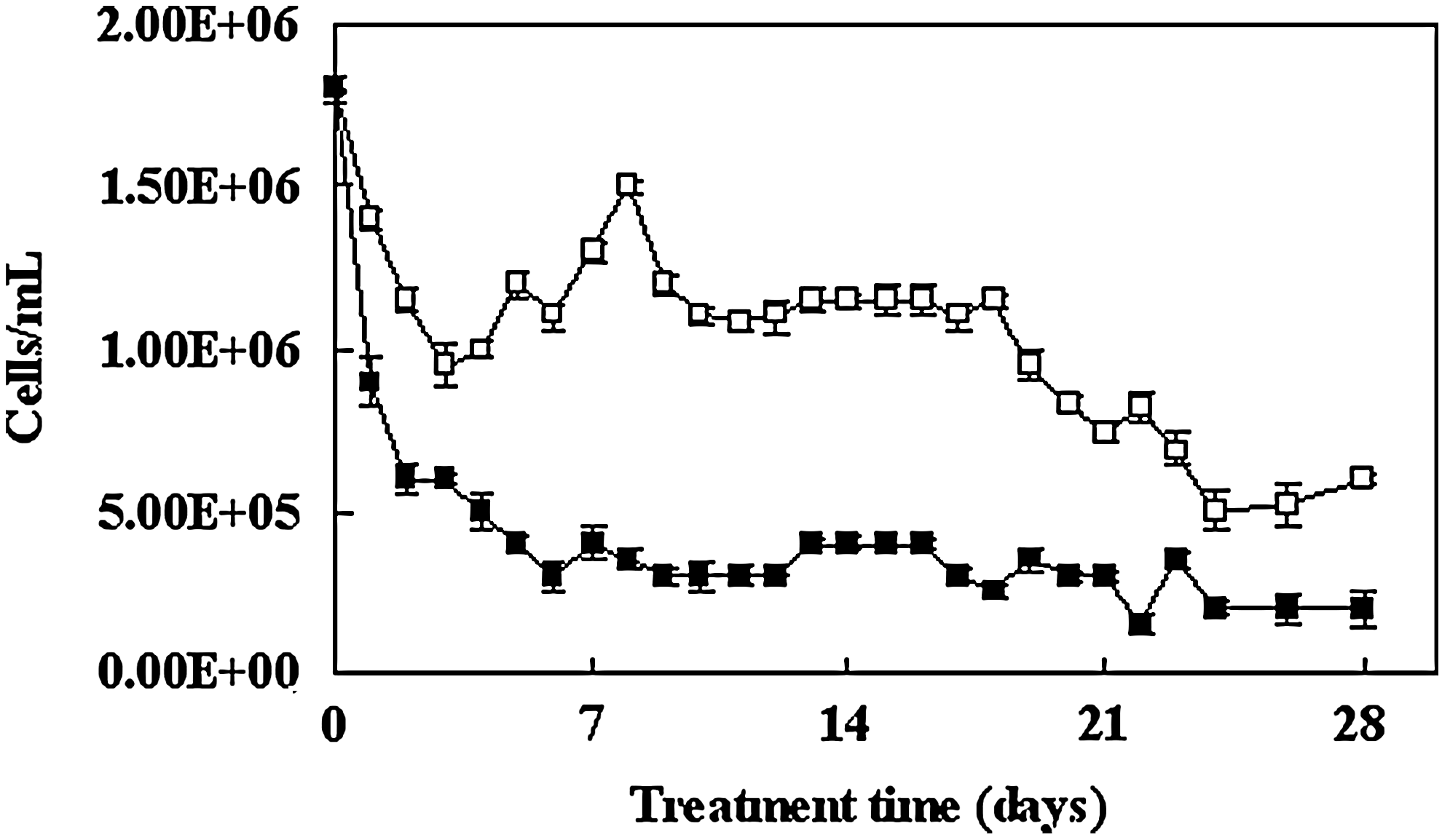
Cyanobacterial cell count without plant microcosms (□) and with plant microcosms (■). Bars represent standard error of the mean (n=3).
As shown in Fig. 5, when the treatment time progressed from day 5 to day 28, turbidity in both planted and control microcosms declined. However, the turbidity of the water in the planted microcosms decreased more rapidly than the turbidity of the water in the control microcosm. After 28 days of treatment, the turbidity in the control microcosm had decreased gradually from 48.2 FTU to 25.7 FTU, while the turbidity in the planted microcosms had decreased from 48.8 FTU to 11.9 FTU. The final turbidity removal of microcosms was over 46% for the control microcosm and 75% for the planted microcosms.

Effects of macrophytes on turibidity. Microcosm without plants (□); microcosms with plants (■). Bars represent standard error of the mean (n=3).
COD removal
Figure 6 illustrates the removal of organic matter, represented by the COD concentration, over the whole experimental period. The results show that COD concentrations were reduced. COD concentrations were lower in the planted microcosms than in the control at all treatment dates, which indicates that the plants had a positive effect on water quality. A reduction of 81.0% COD, with a well-controlled growth of algal bloom as well, was obtained by the planted microcosms in nine days. Water quality was also partially improved (Fig. 5), and the transparency of the water was increased (Fig. 3). It is possible that the decrease in COD is related to the reduction of algal biomass, which would indicate that the algae contributed most to the high COD concentrations when algal blooms occur.
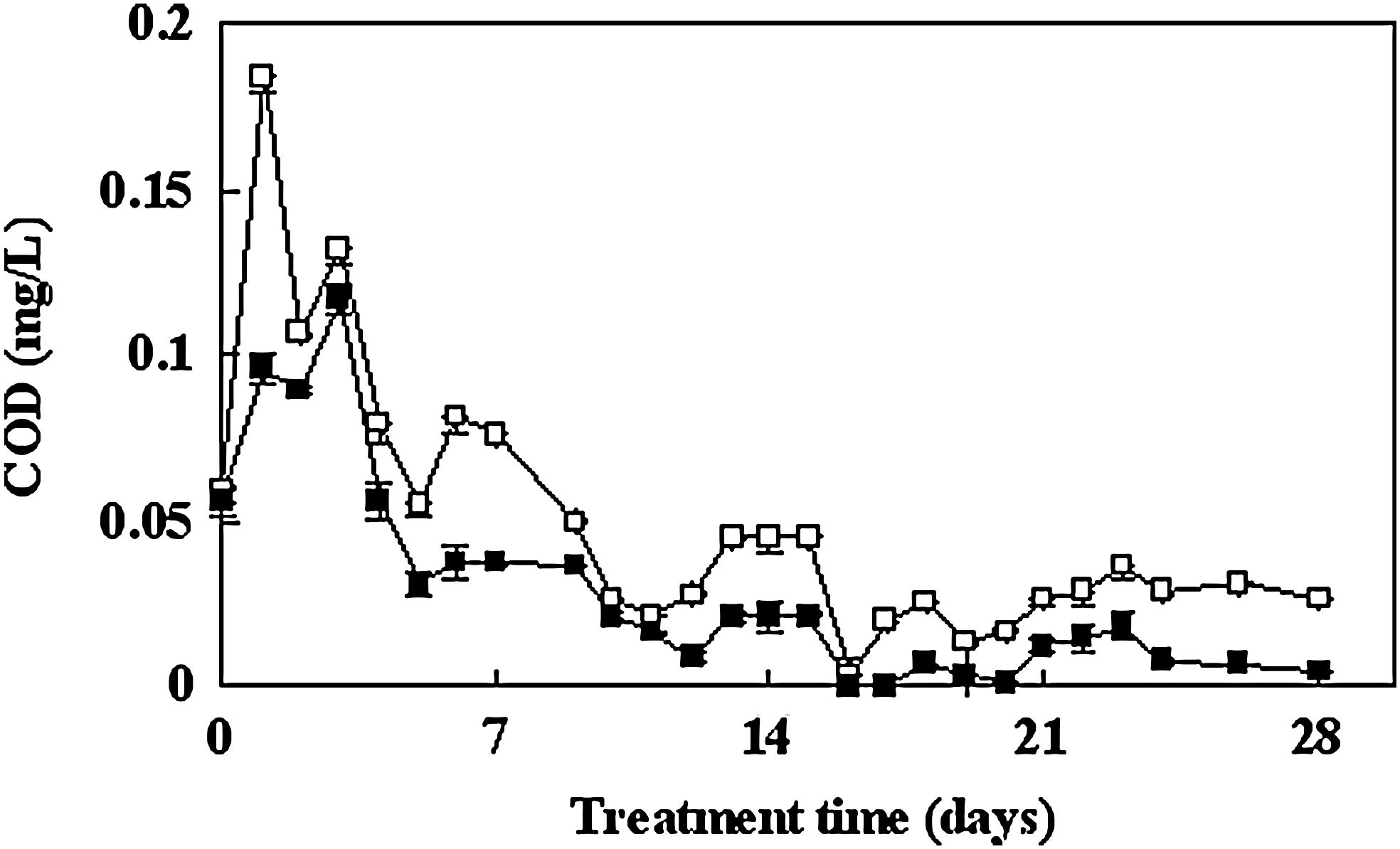
Effects of macrophytes on the concentration of COD. Microcosm without plants (□); microcosms with plants (■). Bars represent standard error of the mean (n=3).
Nutrient removal
The performance of each microcosm was compared for nutrient removal efficiencies. Results from the variation of NH4+, NO2−, NO3−, TN, and PO43− concentrations during the whole experiment are presented in Fig. 7 and Fig. 8. The microcosms were stable after 15 days. At all treatments dates, more nutrient removal was observed in the planted microcosms than in the control microcosm. When the overall removal of nutrients is considered, a faster and more significant removal of nutrients occurred during the early part of the experimental period, that is, before day 10.

Effects of macrophytes on the removal of N from water. Microcosm without plants (□); microcosms with plants (■).
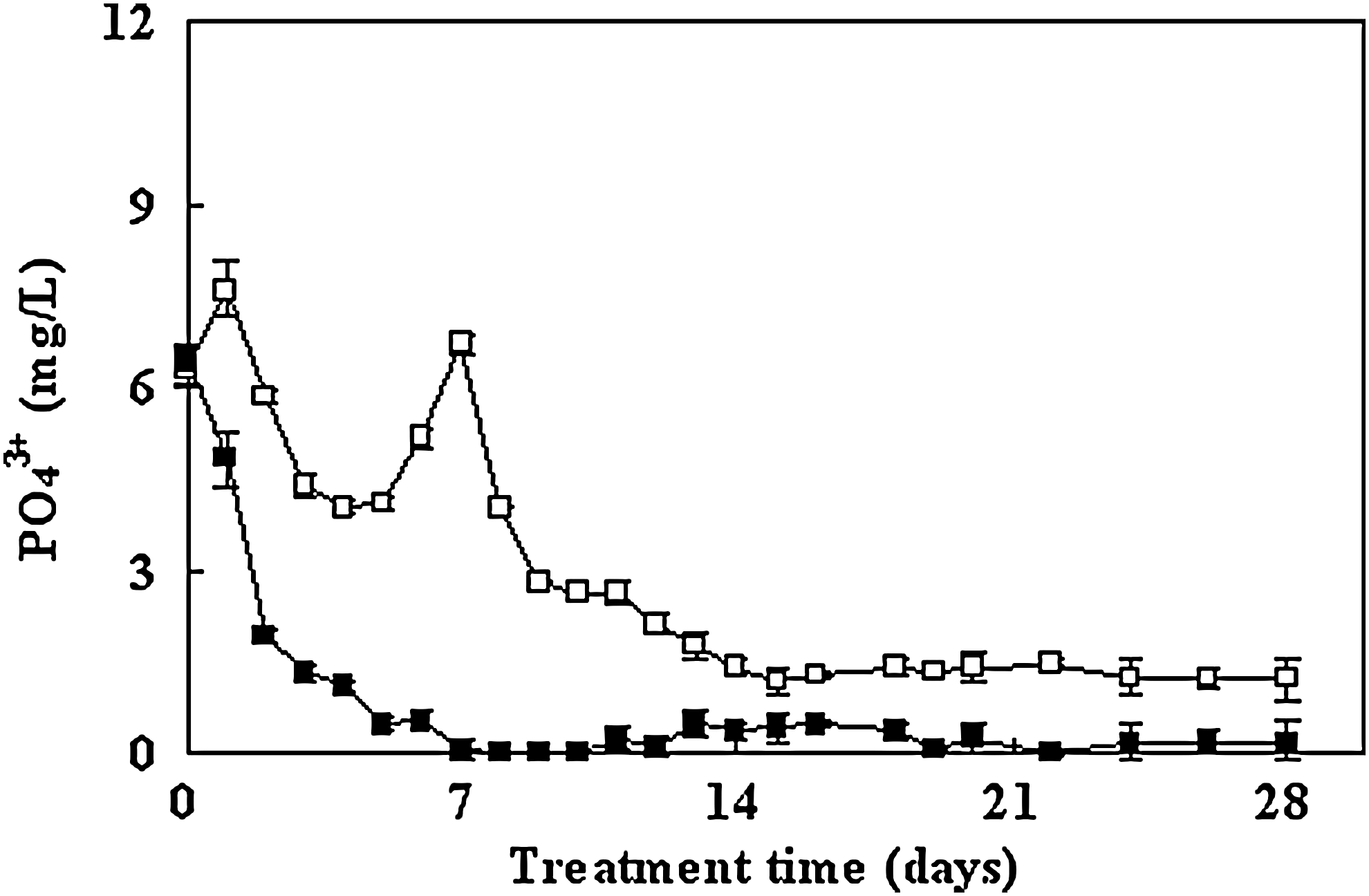
Effects of macrophytes on the removal of phosphate from water. Microcosm without plants (□); microcosms with plants (■). Bars represent standard error of the mean (n=3).
All components of the N removal rate in the planted microcosms were higher than they were in the control. The general trend was for N levels to decrease over time. Comparisons of NO3− and NH4+ removal among the species treatments and the control and across the sampling dates are shown in Figs. 7A and 7B. A rapid decrease in NO3− and NH4+ concentrations was observed on day 2 and day 4 in the planted microcosm. Furthermore, the NO3− and NH4+ concentrations of the planted microcosms took only 9 and 4 days, respectively, to reach a lower value, while the unplanted microcosm took nearly 15 days to do so. The unplanted microcosm also showed consistent and significantly higher NO2− and TN levels at all post-treatment dates (Figs. 7C and 7D). The NO2− and TN concentrations of the planted microcosms reached lower values after 10 and 6 days, respectively, while the unplanted microcosm took nearly 15 days to reach lower concentrations.
Figure 8 shows PO43− levels in the planted and unplanted microcosms across the sampling dates. The data on P removal showed a decrease in the level present, but it was not completely eliminated from any microcosm. The unplanted microcosm showed consistent and significantly higher P levels at all post-treatment dates than the planted microcosms. These results indicate that the planted microcosms had better and more rapid removal of PO43− than the unplanted microcosm. During the first 6 days, a rapid reduction in the concentration of P was observed in the planted microcosms. In these microcosm, the amount of P decreased from 6.42 mg/L to 0.48 mg/L, while in the control microcosm, the amount of P decreased from 6.31 to 4.08 mg/L.
Discussion
Microcosms are considered realistic environmental conditions and highly ecologically of representative of in situ experiments (Bloesch et al., 1988; Caquet et al., 1996; Sugiura, 1996). Microcosm experiments are implemented under complex, near-natural conditions to avoid the limitations of laboratory experiments. The size of microcosm is closely related to the purpose of the investigation. Since the aim of our experiment was to study harmful cyanobacterial growth in the epilimnion, we adjusted the depth of our microcosm to approximately that of the natural ecosystem. However, the size of our microcosm (∼1 m3) may be a problem for studying the structure and function of lake ecosystems.
In some diverse aquatic ecosystems, macrophytes have been successfully used to reduce N and P availability and to inhibit potential cyanobacterial blooms (Cassandra et al., 2006). Some researchers have proposed inhibiting cyanobacterial blooms using allelochemicals and nutrient removal by submerged macrophytes to maintain clear-water states in shallow lakes (Mulderij et al., 2005; Erhard and Gross, 2006).
Our results support earlier findings that plants in water systems can rapidly decrease N and P to levels significantly lower than those in water systems without plants, thus improving water quality. The two parallel planted microcosms had highly similar results. By day 6, algal blooms were effectively inhibited. In both the control microcosm and planted microcosms, turbidity declined obviously when the algal biomass decreased. These observations imply that turbidity can be attributed, to a large extent, to the concentration of algal cells.
The changes of COD were in accord with the biomass (Figs. 4 and 6). A lower algal biomass would induce a lower COD. Our result showed that both the biomass and COD in the planted microcosms were lower than in control microcosm during the entire treatment time. At least one study (Hu et al. 2008b) has reported that high oxygen demand results in higher levels of COD in nutrient-enriched waters. Literature also indicates that the oxygen available in such treatment systems would first be used by the microorganisms for the oxidation of organic matter. Only after COD was satisfied would the oxygen be spared for the nitrification process (Mbuligwe, 2004).
The conversion rates of NH4+ to NO2− and NO2− to NO3− could be higher in the planted microcosms, which may explain why NH4+ and NO2− were lower in planted microcosms than in the control microcosm. The N removal from planted microcosms observed in our experiment (Fig. 7) may be due to the increased plant uptake process rather than microbial denitrification. In the control microcosm, the N removal depended solely on microbial nitrification and denitrification processes.
The reduction of nutrient levels was in agreement with a previous study (Maltais-Landry et al., 2009) that found N levels in water were significantly decreased after planting. This may be because plants have a large biomass and provide a large surface area for the uptake of nutrients and ions. The plant roots provide an additional surface area for microbial colonization. The oxygen transport through the roots facilitates the growth of nitrifying bacteria, which help in the conversion of other forms of N, such as ammoniacal nitrogen (NH3), into NO3− which is either taken up by plants or microbe denitrified. Ghaly et al. (2005) reported that plants prefer NO3− to NH4+. This ultimately results in the removal of NO3− from wastewater (Maltais-Landry et al., 2009).
Gloger et al. (1995) attributed the reduction in NH4+ to direct plant uptake and nitrification. This may explain the higher removal efficiency of NO3− and NH4+ in the planted microcosms compared to that in the unplanted control (Fig. 7A and 7B), likely by enhancing nitrification via oxygen delivery through the rhizosphere. A review by Brix (1997) noted that macrophytes have the ability not only to bind high amounts of nutrients within their system but also to create an environment conducive to decreasing nutrients.
As shown in Fig. 8, a lower amount of PO43− was observed in the planted microcosms during the treatment period. The concentration of the phosphate in the microcosms was lower because the plants absorbed a certain amount of phosphate. P is considered to be a major growth-limiting nutrient in aquatic systems (Wu, 2010b). A comparison of planted and unplanted microcosms shows that the bloom was inhibited because of the lower P concentration of the planted microcosms on day 6. Since P is essential for all metabolic processes, it is a limiting nutrient for cyanobacteria. The excess of P in lake water explains the almost total loss of N from the water column in both unplanted and planted microcosms but the retention of P (Figs. 7 and 8).
Although in our microcosm experiment, the plants could remove N and P efficiently and control algae growth, this method is not suitable for a large area, such as a whole lake. However, in a lake, it is the nearshore water that is usually heavily polluted, and algal blooms often occur there. Given this, we suggest building a planted filter zone, such as a wetland, in nearshore water and water input locations to hold the water within that area to reduce significantly the high nutrient load caused by algal bloom.
Conclusion
The data from the present study demonstrate the importance and efficiency of a remediation engineered model that uses macrophytes to reduce nutrient concentrations and inhibit cyanobacterial growth. The water quality in our planted microcosms was obviously improved through planting. Our study offers a useful algal control method for lakes or reservoirs that suffer from harmful cyanobacterial blooms.
Footnotes
Acknowledgments
The study was supported by the National Natural Science Foundation of China (Grant No. 21007003) and Fundamental Research Funds for the Central Universities FRF-BR-09-009A and CNPC Innovation Fund (2009D-5006-04-02).
Author Disclosure Statement
No competing financial interests exist.