
Abstract
Abstract
Benzo[a]pyrene (BaP) is a carcinogenic polycyclic aromatic hydrocarbon (PAH) that is not known to be a bacterial growth substrate. Organisms capable of cometabolizing BaP in complex field-contaminated systems have not previously been identified. We evaluated BaP mineralization by a bacterial community from a bioreactor treating PAH-contaminated soil during coincubation with or after pre-enrichment on various PAHs as growth substrates. Pyrosequence libraries of 16S rRNA genes were used to identify bacteria that were enriched on the added growth substrate as a means of associating specific organisms with BaP mineralization. Coincubating the bioreactor-treated soil with naphthalene, phenanthrene, or pyrene inhibited BaP mineralization, whereas pre-enriching the soil on the same three PAHs enhanced BaP mineralization. Combined, these results suggest that bacteria in the bioreactor community that are capable of growing on naphthalene, phenanthrene, and/or pyrene can metabolize BaP, with coincubation competitively inhibiting BaP metabolism. Anthracene, fluoranthene, and benz[a]anthracene had little effect on BaP mineralization compared to incubations without an added growth substrate under either coincubation or pre-enrichment conditions. Substantial increases in relative abundance after pre-enrichment with phenanthrene, naphthalene, or pyrene, but not the other PAHs, suggest that members of the genera Cupriavidus and Luteimonas may have been associated with BaP mineralization.
Introduction
B
Bioremediation is a primary strategy for reducing PAH contamination in soil (USEPA Office of Solid Waste and Emergency Response, 2010), but the efficacy of bioremediation may be limited by its ability to meet cleanup standards for the carcinogenic PAHs at a given site. Many microorganisms can grow on PAHs containing two, three, or four rings, but no microorganisms are known to grow on PAHs containing five or more rings. Although there have been no reports of bacteria capable of utilizing BaP as a sole carbon and energy source, several bacteria can oxidize BaP when a suitable cosubstrate is present (Juhasz et al., 1997; Boonchan et al., 2000; Marcoux et al., 2000; Dries and Smets, 2002; Zhao and Wong, 2010; Ping et al., 2014) or after growth on a suitable enrichment substrate (Juhasz et al., 1997; Aitken et al., 1998; Chen and Aitken, 1999; Moody et al., 2004; Rentz et al., 2008).
We previously used DNA-based stable isotope probing (SIP) to describe the bacterial guild able to grow on naphthalene, phenanthrene, anthracene, pyrene, fluoranthene, and benz[a]anthracene in a PAH-contaminated soil obtained from an MGP site (Jones et al., 2011a). In the present study, we evaluated the ability of the bacterial community in a bioreactor used to treat the MGP soil to mineralize BaP during coincubation or after pre-enrichment with each of these six PAHs. The primary objective was to identify those PAH growth substrates associated with BaP metabolism. We hypothesized that any organism capable of cometabolizing BaP would mineralize BaP if it grew in response to pre-enrichment with a PAH growth substrate. We also hypothesized that BaP mineralization would be competitively inhibited in the simultaneous presence (coincubation) of a PAH substrate that could serve as a growth substrate for an organism otherwise capable of cometabolizing BaP. Pyrosequencing of 16S rRNA genes from the communities enriched on the various PAHs was conducted to identify those organisms that increased in relative abundance in response to the growth substrates associated with BaP mineralization. Identifying the organisms associated with BaP metabolism in complex systems is a first step toward elucidating the genetic determinants of BaP metabolism and toward developing bioremediation strategies to improve the removal of BaP and other carcinogenic PAHs.
Materials and Methods
Growth substrates and chemical reagents
Naphthalene, phenanthrene, pyrene, fluoranthene, and unlabeled benzo[a]pyrene were obtained from Sigma-Aldrich (St. Louis, MO), anthracene was obtained from Eastman Kodak (Rochester, NY), and benz[a]anthracene was obtained from Acros Organics (Fair Lawn, NJ). [7,10-14C]Benzo[a]pyrene (68 mCi/mmol) was obtained from GE Healthcare UK Limited (Buckinghamshire, United Kingdom). All other reagents were of the highest purity available. All solvents were molecular biology or high-pressure liquid chromatography (HPLC) grade.
Bioreactor conditions and maintenance
PAH-contaminated soil was obtained from the site of a former MGP in Salisbury, North Carolina and processed as described elsewhere (Richardson and Aitken, 2011). Briefly, large objects were removed by hand; the soil was sieved through a 10-mm wire screen, blended, and sieved again through #6 mesh (3.35 mm) before storage in the dark at 4°C. The processed soil (64% sand, 30% silt, 6% clay, 15% moisture, pH=7.6) was treated in a bench-scale, semi-continuous, aerobic, slurry-phase bioreactor with a 7-day treatment cycle as described previously (Singleton et al., 2011); 20% of the treated soil slurry was replaced every 7 days with untreated (processed) soil in a buffer containing 5 mM potassium phosphate (pH 7.5) supplemented with 5 mM NH4NO3 (reactor buffer), resulting in a solids retention time of 35 days. At the time these experiments were done, the bioreactor had been operating for over 18 months.
Mineralization experiments
Effects of coincubation and pre-enrichment with two-, three-, and four-ring PAHs on BaP mineralization were tested using the bioreactor-treated soil. The capacity for BaP mineralization was followed in a short-term (24-h) assay in which incubation flasks were amended with 14C-labeled BaP. For the coincubation experiment, triplicate 125-mL Erlenmeyer flasks were spiked with 625 μg of an unlabeled PAH in acetone and 60,000 dpm of radiolabeled BaP in methanol (the corresponding amount of labeled BaP added was 0.09 μg/flask, or 3 μg/L). After evaporation of the carrier solvents, 5 mL of treated soil slurry from the bioreactor, 25 mL of reactor buffer, and a CO2 trap (Singleton et al., 2008) were added to each flask. The flasks were capped with foil-covered, Teflon-lined screw caps and agitated on an orbital shaker (150 rpm) in the dark and at room temperature (21–23°C) for 24 h. The CO2 trap was analyzed by liquid scintillation counting.
For the pre-enrichment experiment, triplicate flasks prepared as described above for the coincubation experiment were spiked with a growth substrate without BaP. In a preliminary experiment, 7 days was determined to be adequate for significant removal of each growth substrate (Supplementary Fig. S1). In separate clean flasks, 60,000 dpm of radiolabeled BaP in methanol was added and the methanol allowed to evaporate. Slurries from the 7-day incubations were transferred to the clean flasks containing the radiolabeled BaP, and a CO2 trap was added to each flask. The flasks were capped and shaken for 24 h, then each CO2 trap was analyzed by liquid scintillation counting. Triplicate flasks with no growth substrate added were also prepared for the coincubation and pre-enrichment experiments to serve as controls. Triplicate flasks with no substrate added, but amended with 85% phosphoric acid to pH <2 were prepared as inhibited controls. Data from each experiment were compared using a two-tailed Student's t-test assuming unequal variance.
To measure recovery of 14C in each flask, after removing the CO2 trap the flasks containing radiolabeled BaP were acidified with 200 μL of 85% phosphoric acid, a new CO2 trap was placed in the flask, and the flasks were agitated for 3 h. Soil slurry (1 mL) from each flask was then mixed with 1 mL of ethyl acetate in separate 15-mL conical-bottom centrifuge tubes. The tubes were vortexed for 1 min and centrifuged for 5 min. The CO2 trap and an aliquot of the ethyl acetate were analyzed by liquid scintillation counting. Radiocarbon recovery was determined by summing the activity measured in the initial (24-h incubation) CO2 trap, the subsequent 3-h CO2 trap, and the soil slurry. There was no difference in 14C recovery between flasks with or without added growth substrate compared to live and acid-inhibited flasks without added growth substrate (data not shown). Recovery of 14C in CO2 traps in acid-inhibited flasks from either coincubation or pre-enrichment conditions was <0.2% (data not shown).
Concentrations of each PAH in the bioreactor slurry and incubation flasks were determined by HPLC as described elsewhere (Richardson et al., 2011; Hu et al., 2012).
Microbial community analysis
We considered the possibility that, although it was added at a very low concentration, the BaP added in the mineralization experiments might influence the microbial community. Therefore, for each PAH growth substrate (and the controls with no substrate added), a set of duplicate flasks was set up as described for the mineralization experiments, except that the BaP was added in unlabeled form (0.09 μg/flask, or 3 μg/L). For the pre-enrichment condition, two sets of duplicate flasks were prepared. In one set, after the 7-day preincubation, an additional incubation for 24 h was conducted in the presence of unlabeled BaP, and in the second set the additional incubation was conducted in the absence of BaP. From each flask, 2 mL of soil slurry was removed at the end of the 24-h additional incubation period and centrifuged. DNA was extracted from the soil pellet using the FastDNA® Spin Kit for Soil (MP Biomedicals, Solon, OH) according to the accompanying instructions, except that DNA was eluted with Tris-EDTA buffer (TE, pH=8.0). Aliquots of DNA from each replicate extraction were pooled. Each pooled sample was polymerase chain reaction (PCR)-amplified in triplicate using a unique pair of barcoded primers targeting region 27F to 338R of the 16S rRNA gene. Each primer consisted of an eight-base barcode (Supplementary Table S1) and a two-base spacer (TC for the forward primer, CA for the reverse primer) (Hamady et al., 2008) followed by 27F (AGAGTTTGATCCTGGCTCAG) (Lane, 1991) or 338R (TGCTGCCTCCCGTAGGAGT) (Suzuki and Giovannoni, 1996). For DNA from the pre-enrichment condition with phenanthrene in the presence of BaP, three sets of triplicate PCR reactions were performed to evaluate the reproducibility of pyrosequencing.
Each set of triplicate PCR products was pooled, and the amplicon was recovered in TE using a QIAquick PCR Purification Kit (Qiagen, Valencia, CA) with the microcentrifuge protocol. The DNA concentration in each cleaned amplicon was quantified with a NanoDrop 3300 fluorospectrometer (NanoDrop Products, Wilmington, DE) using the Quant-iT PicoGreen dsDNA Assay Kit (Invitrogen, Eugene, OR). An aliquot containing 7.6 ng of DNA from each sample was submitted to the UNC-Chapel Hill High-Throughput Sequencing Facility (HTSF) for sequencing adapter ligation and multiplex pyrosequencing using the Life Sciences 454 Genome Sequencer FLX Titanium platform (Roche Diagnostics Corp., Branford, CT).
Analysis of pyrosequence libraries
Processing and clustering analysis of the sequences was conducted using Mothur software and tools (Schloss et al., 2009). Sequences were parsed into libraries based on the presence of the eight-base barcode. All sequence reads were quality checked; primer dimers, poor quality reads, or reads less than 200 bases in length were removed. Sequences passing the quality filters were trimmed at 350 bases. Sequences retained following the quality filtering process were aligned using the SILVA reference database (Pruesse et al., 2007). Uchime (Edgar et al., 2011) was used to remove chimeric sequences from the data set. Before processing, there were 237,788 total sequences; after eliminating sequences that were too short, of poor quality, or without detected barcodes, the 24 libraries collectively contained 92,794 sequences.
The Mothur-formatted Ribosomal Database Project (RDP) reference set (http://rdp.cme.msu.edu) was used to classify the aligned sequences with an 80% cutoff. Sequences were clustered into operational taxonomic units (OTUs) with a cutoff of 97% similarity and a confidence threshold of 80%. Because anthracene group 1 and pyrene group 2 bacteria (Jones et al., 2011a) would not be captured in the RDP reference set, a local BLAST search using the blastn application within BLAST+ (Camacho et al., 2009) was used to search for sequences in the pyrosequencing data related to those groups. Other groups of interest were identified based on the classification data obtained from Mothur. Trees generated in Mothur were used as the input for UniFrac analyses (Lozupone et al., 2006). The category mapping variables used as input for these analyses and a description of each library are in Table 1.
ANT, anthracene; BaA, benz[a]anthracene; BaP. benzo[a]pyrene; BRS, bioreactor slurry (inoculum for all experiments); Co, coincubation; FLA, fluoranthene; Pre, pre-enrichment; NAP, naphthalene; None, no exogenous substrate; PHE, phenanthrene; PYR, pyrene.
Results
BaP mineralization
The bioreactor-treated soil mineralized 15% of BaP over a 24-h incubation without any additional substrate (Fig. 1), indicating that the microbial community in the reactor contained organisms able to metabolize BaP. The addition of naphthalene, phenanthrene, and pyrene inhibited BaP mineralization over a 24-h coincubation compared to the control in which no exogenous substrate was added (i.e., incubation with any of these cosubstrates led to significantly less mineralization than in the control; p<0.01) (Fig. 1). Coincubation with anthracene, fluoranthene, or benz[a]anthracene had no effect (p>0.1).
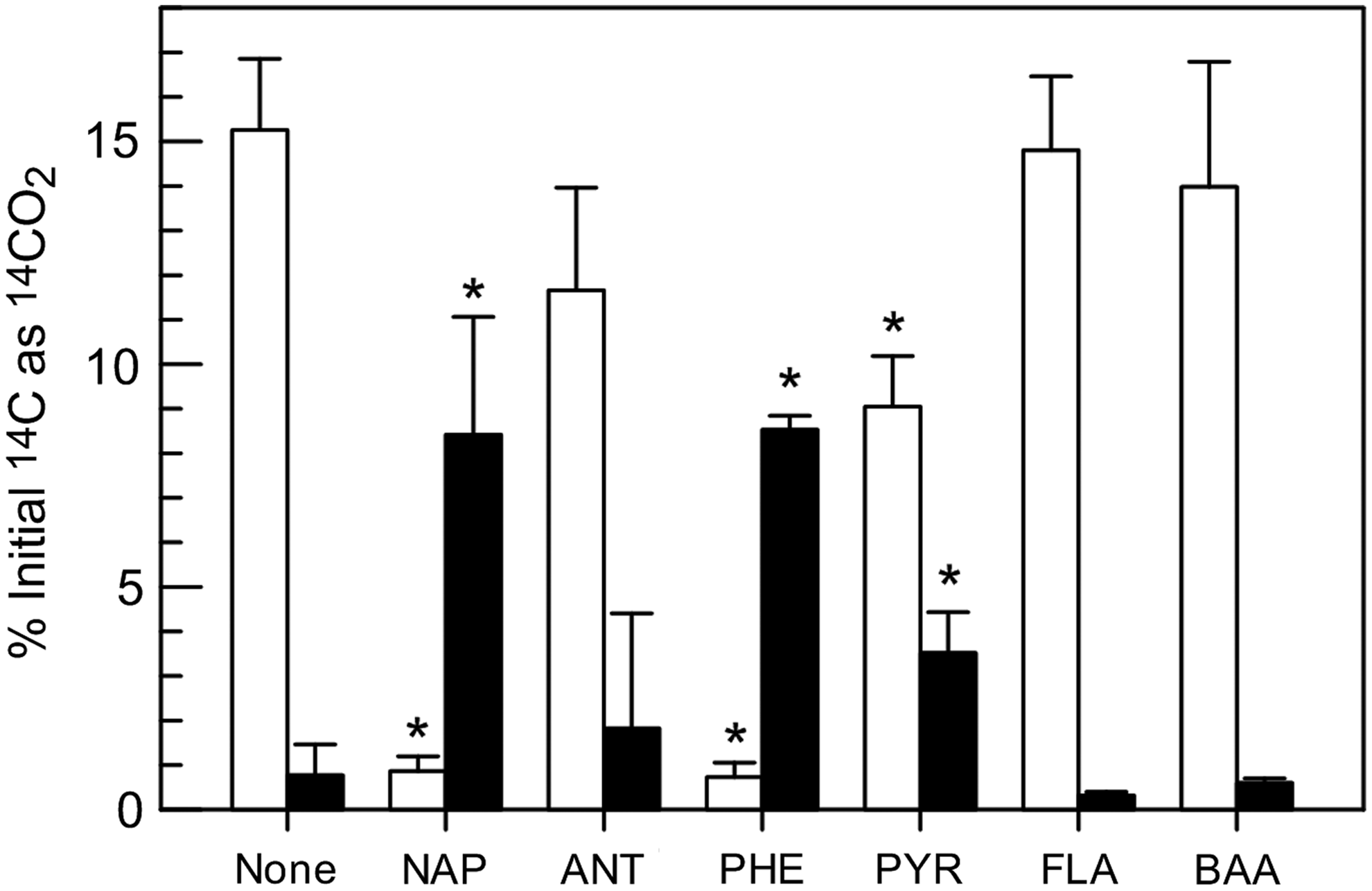
Initial 14C evolved as 14CO2 after 24 h in the presence of 14C-BaP (mean and standard deviation, n=3). Open and closed bars are data from coincubation and pre-enrichment experiments, respectively, with the indicated growth substrate or in the absence of an added growth substrate (None). Data represent means and standard deviations of triplicate flasks for each condition. Asterisks indicate that the value is significantly different from the respective incubation without exogenous substrate. Abbreviations are as in Table 1.
To evaluate the effect of pre-enrichment with a PAH growth substrate, the microbial community in the bioreactor-treated soil slurry was incubated in the presence of a PAH growth substrate for 7 days, then BaP mineralization was tested over an additional 24 h (residual concentrations of each PAH after the 7-day incubation period are shown in Supplementary Fig. S2). Pre-enrichment with naphthalene, phenanthrene, or pyrene enhanced BaP mineralization (p<0.05) compared to incubation for 7 days in the absence of exogenous substrate (Fig. 1). The other PAH growth substrates had no effect (p>0.1).
Analysis of pyrosequencing libraries
The libraries and corresponding incubation conditions evaluated are summarized in Table 1. Supplementary Table S1 summarizes the number of sequences and OTUs, and the richness and diversity estimates for each library. The coincubation libraries were generally richer and more diverse than the pre-enrichment libraries, which is consistent with the extended period of incubation in the presence of a dominant growth substrate in the pre-enrichment experiments. The pairwise UniFrac significance test (weighted to account for differences in the number of sequences recovered) determined that there was no significant difference among the coincubation libraries (p>0.1) or between any coincubation library and the bioreactor slurry inoculum library (p>0.1). There was also no significant difference in the distance between pairs of pre-enrichment libraries generated after an additional 24 h of incubation with or without BaP (p>0.1), suggesting that the presence of a trace concentration of BaP (3 μg/L) had no effect on the overall bacterial community. DNA from the bioreactor slurry pre-enriched on phenanthrene and then amended with BaP was used to generate triplicate libraries (designated PHB1, PHB2, and PHB3; Table 1) to illustrate the reproducibility of the pyrosequencing results. Despite the difference in the number of sequences recovered from each replicate (Supplementary Table S1), the pairwise UniFrac significance test (weighted) determined that there was no significant difference among these libraries (p>0.1).
Principal coordinates analysis of the pyrosequencing libraries (weighted and normalized) accounted for 62% of the variation among the libraries, which could be grouped into distinct clusters (Fig. 2). Bacterial communities resulting from coincubation (cluster 1 and the library from coincubation with phenanthrene, CPHB) differed from those resulting from pre-enrichment (clusters 3 and 4). Although not indicated by the pairwise UniFrac significance test, principal coordinates analysis suggested that the library from coincubation with phenanthrene (CPHB) was different from the other coincubation libraries (Fig. 2). The libraries from preincubation experiments in which a PAH growth substrate was not added (PNOB and PNON) were dissimilar from all other pre-enrichment libraries. They were also different from the library from coincubation in the absence of exogenous growth substrate (CNOB), suggesting that the longer incubation period (8 days in preincubation vs. 24 h in coincubation) led to a shift in the community. The libraries from pre-enrichment with benz[a]anthracene, either with (PBAB) or without (PBAA) BaP addition, were also different from all other pre-enrichment libraries (Fig. 2). These libraries were most similar to the libraries from preincubation in the absence of exogenous growth substrate (PNOB and PNON), which may have resulted from less growth on benz[a]anthracene compared to the other PAHs. The relationships among bacterial communities from the various incubation conditions illustrated in Figure 2 are supported by cluster dendrogram analysis (Supplementary Fig. S3).
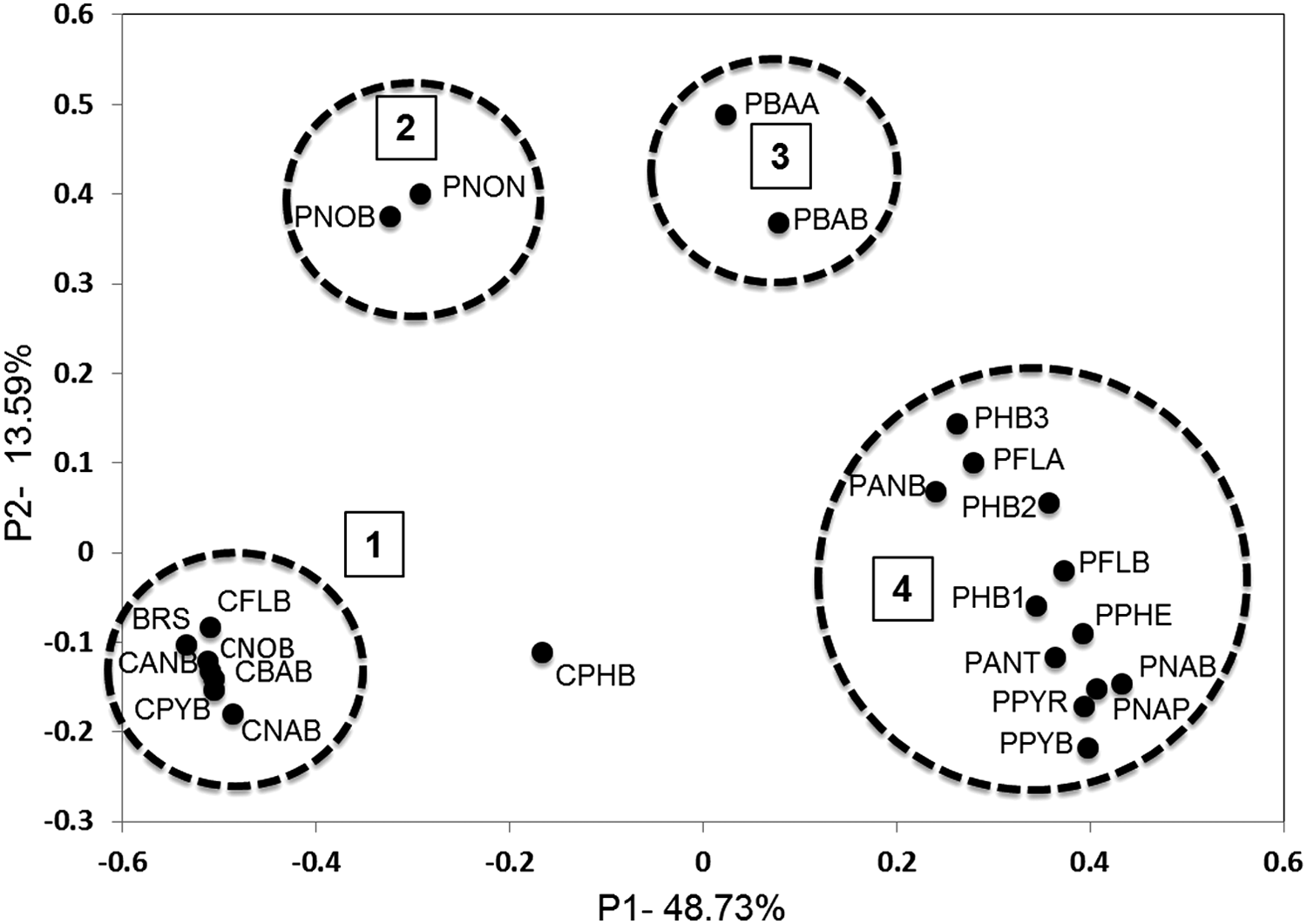
Weighted and normalized principal coordinates analysis of pyrosequence libraries. Clusters of libraries referred to in the text are circled and numbered. Libraries are defined in Table 1. Clusters beginning with the letter C are from coincubation experiments and with the letter P are from the pre-enrichment experiments; BRS, bioreactor slurry.
Taxonomic representation in pyrosequence libraries
The major bacterial taxa represented in each library are summarized in Supplementary Figure S4. Proteobacteria accounted for the majority of sequences recovered from each library. Relative abundances of 16S rRNA gene sequences representing all of the bacterial genera and groups associated with growth on the six PAH substrates in the untreated soil by SIP (Jones et al., 2011a), as well as all other genera and groups representing at least 1% of the sequences in any library, are provided in Supplementary Table S2 for the pre-enrichment conditions and Supplementary Table S3 for the coincubation conditions.
Differences among the clusters shown in Figure 2 are manifested as differences among the major taxa (Table 2). Clusters 1 and 4 differed greatly in the representation of Alpha- and Betaproteobacteria, whereas clusters 2 and 3 differed in the representation of Alphaproteobacteria and Acidobacteria. Library CPHB, which did not align with any cluster, differed from cluster 1 in the representation of Betaproteobacteria (due to substantial enrichment of Acidovorax; Supplementary Table S3) and from cluster 4 in the abundance of Alphaproteobacteria and Acidobacteria sequences.
Cluster designations are as in Figure 2. Except for CPHB (n=1), values are mean±standard deviation (cluster 1, n=7; cluster 4, n=12) or mean and range (clusters 2 and 3, n=2).
—, not detected in the library or any library in the cluster.
Discussion
Although BaP is known to be mineralized by bacteria, it is difficult to determine which organisms are responsible for BaP mineralization in complex field-contaminated systems because no organisms have been isolated with BaP as a sole carbon source. Molecular techniques such as SIP can detect only those organisms capable of assimilating carbon during growth on a given substrate (Radajewski et al., 2000), not organisms that cometabolize an organic compound in the absence of growth. On the assumption that BaP is removed from a contaminated system primarily through cometabolism, our approach was to identify those PAH growth substrates that could influence the mineralization of BaP by organisms in a bioreactor treating soil from a field-contaminated site. These organisms would already have been enriched in the bioreactor as a result of growth on PAHs and other carbon sources in the soil.
BaP mineralization in the bioreactor-treated soil slurry was inhibited in the presence of added naphthalene, phenanthrene, and pyrene compared to incubation in the absence of added substrate over a 24-h period (Fig. 1). Competitive inhibition of PAH metabolism has been observed in a number of cases when compounds were incubated together, and occurs when more than one substrate is metabolized by the same enzymes (Stringfellow and Aitken, 1995; Shuttleworth and Cerniglia, 1996; Kanaly and Bartha, 1999). Coincubation of anthracene, benz[a]anthracene, or fluoranthene with BaP did not inhibit BaP mineralization, suggesting that organisms capable of growing on or metabolizing these substrates either do not mineralize BaP or do so through pathways that are independent of the pathway(s) for metabolism of the other PAHs.
Incubation of the bioreactor soil slurry in the absence of exogenous substrate for 7 days (the preincubation condition with no added substrate) led to almost a complete loss of BaP mineralization activity compared to the treated soil removed directly from the bioreactor (the coincubation condition with no added substrate; Fig. 1). This loss of activity could have resulted from the decay of organisms associated with BaP mineralization under substrate-limited conditions, a decline in the metabolic capacity to mineralize BaP in one or more of those organisms, or both. Pre-enrichment with naphthalene, phenanthrene, or pyrene as a growth substrate restored BaP mineralization activity, whereas pre-enrichment with anthracene, benz[a]anthracene, and fluoranthene did not.
Association of bacterial groups with BaP mineralization
The combined results of the coincubation and pre-enrichment experiments strongly suggest that one or more organism(s) capable of growing on naphthalene, phenanthrene, and/or pyrene are able to mineralize BaP, whereas bacteria that grew on anthracene, benz[a]anthracene, or fluoranthene are not. Accordingly, we anticipated that these differences would be manifested in the pyrosequencing libraries from pre-enrichment with the various PAH growth substrates. Although the libraries from pre-enrichment with all of the growth substrates other than benz[a]anthracene were grossly similar (Fig. 2 and Supplementary Fig. S4), we examined these libraries for differences in representation of individual sequences between preincubation with an added growth substrate and preincubation without an added growth substrate (Supplementary Table S2). For those sequences that increased in relative abundance in one or more libraries in which a growth substrate was added, we looked for much greater increases in response to naphthalene, phenanthrene, and/or pyrene than in response to anthracene, benz[a]anthracene, or fluoranthene.
The bacterial genera that were at least 3% of the total sequences in any library from pre-enrichment with naphthalene, phenanthrene and/or pyrene are summarized in Figure 3. Relative abundances from incubations without BaP amendment for an additional 24 h are shown along with relative abundances from incubations with BaP amendment to illustrate the reproducibility (or not) for a given pre-enrichment substrate. We believe that substantial differences between incubations amended with BaP and those without BaP (e.g., Cupriavidus in response to pre-enrichment with phenanthrene) are more likely to have resulted from experimental variability at the individual sequence level than from an effect of BaP per se, as the BaP concentration was very low (3 μg/L). With the exception of Acidovorax, all of the genera shown in Figure 3 were 1% or less of the sequences in the bioreactor slurry used as inoculum for the pre-enrichment experiments.
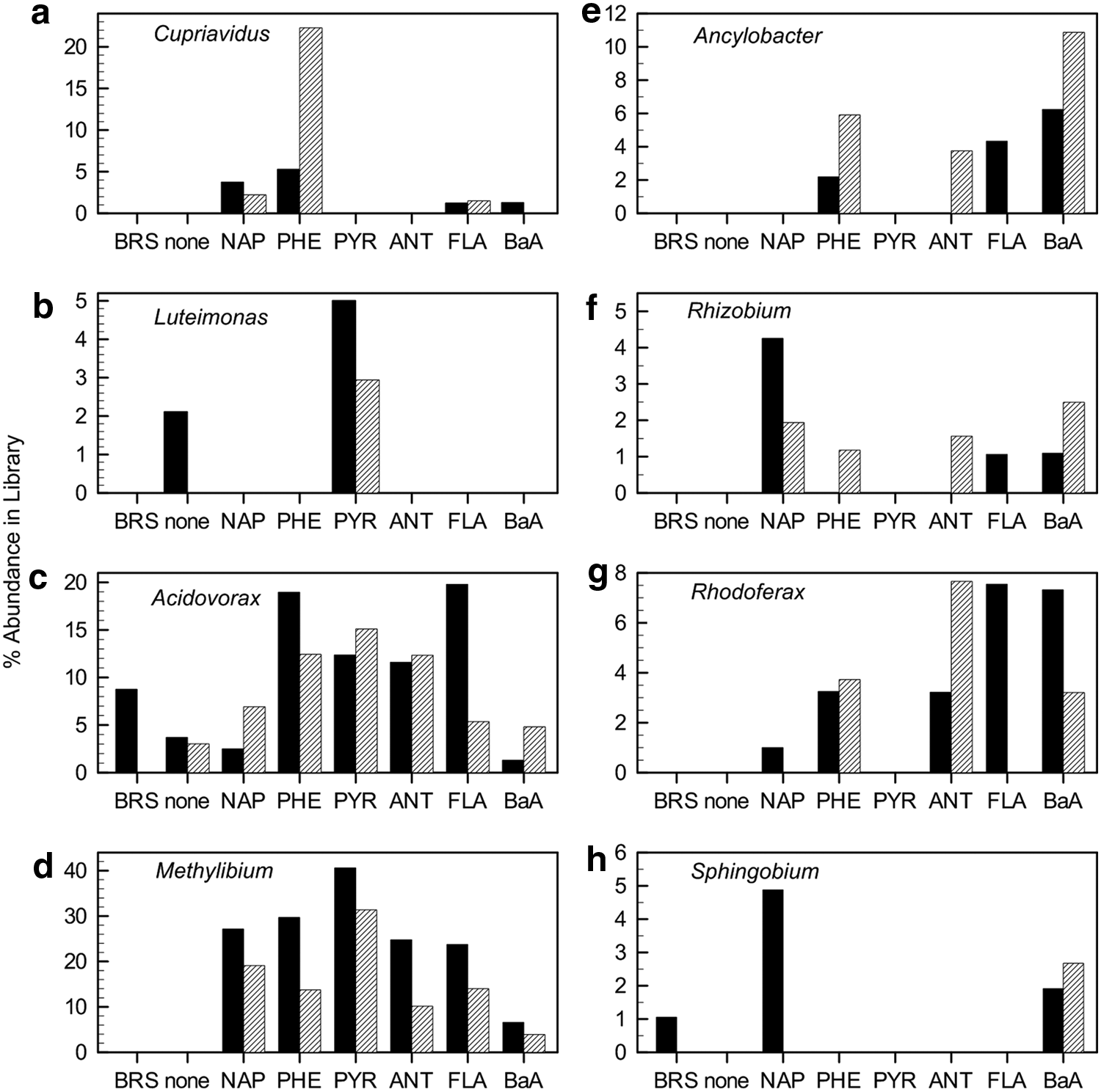
Responses of selected bacterial genera (as % of the respective pyrosequence library) to preincubation with each of the polycyclic aromatic hydrocarbon (PAH) substrates or in the absence of added substrate (None) in comparison to the abundance in the inoculum (BRS); abbreviations are as in Table 1. Solid bars represent data from incubations in the absence of BaP and hatched bars represent incubations in the presence of BaP. No bar is shown if the genus was not in the library or was present at <1%.
Of the eight genera whose relative abundances increased substantially in response to naphthalene, phenanthrene, and/or pyrene compared to the incubations with no added substrate (Fig. 3), most of these also increased in response to at least one of the other growth substrates. For example, Acidovorax and Methylibium sequences increased substantially in response to all of the PAHs. The data in Figure 3 suggest that these organisms, as well as those associated with the genera Ancylobacter, Rhizobium, and Rhodoferax, have a broad response to PAHs, but not likely an ability to cometabolize BaP. The response of sequences representing the genus Sphingobium (Fig. 3) is ambiguous; there was a substantial increase in relative abundance after pre-enrichment with naphthalene in the absence of unlabeled BaP amendment, but not in its presence. Sequences associated with Sphingobium also increased to between 2% and 3% of the libraries after pre-enrichment with benz[a]anthracene, suggesting that it responded to one of the substrates not associated with the enhancement of BaP mineralization.
One genus, Cupriavidus, increased to more than 20% of the sequences after pre-enrichment with phenanthrene (amended with unlabeled BaP for an additional 24 h) and to a lesser extent in response to naphthalene (2–4% of the naphthalene libraries). Although sequences associated with Cupriavidus also increased slightly (1.5% or less) in response to fluoranthene and benz[a]anthracene, the increases were not as substantial as with phenanthrene or naphthalene. We, therefore, suggest that one or more members of the genus Cupriavidus are likely candidates for association with BaP mineralization. Sequences associated with the genus Luteimonas increased in relative abundance (3–5%) after incubation with pyrene, whereas these sequences remained <1% of the total sequences for all of the other PAH substrates; therefore, members of Luteimonas may be associated with BaP mineralization as well.
The genus Cupriavidus, formerly Ralstonia and Wautersia (Vandamme and Coenye, 2004), is a member of the family Burkholderiaceae that contains many known PAH-degrading genera. Cupriavidus spp. have been isolated from petroleum-contaminated soil (Jurelevicius et al., 2010), and they have been associated with growth on phenanthrene (Widada et al., 2002) and the degradation of substituted aromatic compounds (Pérez-Pantoja et al., 2008). We recovered similar sequences from SIP experiments with salicylate and naphthalene in a different PAH-contaminated soil (Singleton et al., 2005; Powell et al., 2008), and they were later associated with naphthalene mineralization and growth in response to the addition of nonionic surfactants to the bioreactor treating that soil (Zhu et al., 2010).
A Luteimonas isolate was recently described after being recovered from hydrocarbon-contaminated soil from an industrial site (Zhang et al., 2010), and another isolate was recovered from a biofilter treating waste gas containing furan (Finkmann et al., 2000); otherwise, this genus has not previously been associated explicitly with PAH degradation. Many of the other groups enriched under the preincubation conditions have been associated with hydrocarbon degradation, but not necessarily PAH degradation (Jeon et al., 2004; Kasai et al., 2006; Popp et al., 2006; Han et al., 2009; Saavedra et al., 2010).
Role of SIP-identified PAH degraders
Based on the rationale described above, we conclude that none of the PAH degraders identified by SIP of the untreated soil (Jones et al., 2011a, summarized in Supplementary Table S2) appears to be associated with BaP mineralization in the bioreactor community, even though the pre-enrichment experiments performed in this study mimicked the incubations performed in the previous SIP experiments. The bacterial groups resulting from pre-enrichment experiments with the bioreactor-treated soil slurry are mostly quite different from those resulting from SIP experiments with untreated soil. For example, anthracene group 1, the dominant anthracene-degrading group identified by SIP with anthracene in the untreated soil (Jones et al., 2011b), was not enriched after incubating the bioreactor slurry with anthracene for 7 days (Supplementary Table S2).
Among the major PAH degraders identified by SIP of the untreated soil, only Acidovorax sequences were found to be present at >1% in the bioreactor slurry used as inoculum in this study (Supplementary Table S2). Members of Acidovorax were the predominant degraders of phenanthrene in the earlier SIP study on untreated soil (Jones et al., 2011a), which is consistent with their enrichment to 38% of the library from coincubation with phenanthrene (Supplementary Table S3) and 15% of the library from pre-enrichment with phenanthrene (Supplementary Table S2). Since Acidovorax sequences also increased in abundance in response to all the other PAHs relative to preincubation with no added substrate, we conclude that Acidovorax spp. in the bioreactor were not associated with BaP mineralization. Sequences associated with Thiobacillus were also abundant (11%) in the bioreactor slurry library (Supplementary Table S2), but Thiobacillus were observed only as a singleton sequence in the clone library from SIP on benz[a]anthracene (Supporting Information in Jones et al., 2011a). They also were not enriched under any pre-enrichment condition (Supplementary Table S2), suggesting that they were not a predominant degrader of any PAH investigated in this study.
Summary
The approach used in this study to identify PAH growth substrates capable of supporting BaP cometabolism can complement efforts to isolate organisms capable of cometabolizing BaP. Our collective work on SIP of PAH-degrading bacteria in field-contaminated systems (Singleton et al., 2005, 2006, 2007; Jones et al., 2008, 2011a, 2011b) has indicated that a number of organisms associated with PAH degradation in these systems have not previously been isolated or characterized. Therefore, enrichment techniques such as we employed in this study can represent a first step toward elucidating physiological features of important PAH degraders. Although the methods in this study provide only indirect evidence to associate one or more organisms with BaP mineralization, we are not aware of another cultivation-independent approach to identifying organisms that metabolize hydrocarbons that do not serve as carbon or energy sources. In turn, identifying bacteria capable of specific compound degradation in a complex system and associating those bacteria with other metabolic abilities can help to target isolation efforts. Ultimately, experimentation with bacterial isolates will lead to definitive evidence of the metabolic activity suggested by molecular results.
Footnotes
Acknowledgments
This work was supported by the National Institute of Environmental Health Sciences (grant 5 P42 ES005948). The authors thank Jesse Walsh and Xiaojun Guan (UNC HTSF) for performing the pyrosequencing and processing the pyrosequencing data files, respectively.
Author Disclosure Statement
All authors confirm that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.