
Abstract
Abstract
The treatment performance and microbial diversity of two parallel single-stage activated sludge bioreactors were investigated and compared. An innovative upflow microaerobic sludge bed (UMSB) bioreactor and an aerobic control system (ACS) were operated at dissolved oxygen levels of 0.5–0.9 and 2.5–4 mg/L, respectively. Under similar conditions, at a hydraulic retention time of 25 h, an organic loading rate of 1.4±0.1 kg COD/m3·d, and an influent total phosphate (TP) of 43±4 mg/L, the chemical oxygen demand (COD) removal efficiencies of the UMSB and the ACS were 94%±1% and 95%±1%, respectively, and no significant difference in performance was noted between the two systems (p=0.498). It was, however, noted that TP removal efficiency of the UMSB (57%±5%) was significantly higher compared with ACS (34%±6%) (p=0.00). Phylogenetic analysis indicated that bacteria in the UMSB and ACS were highly diverse and obviously different. It is likely that facultative anaerobes, microaerophiles, and aerobes were able to coexist in the UMSB. This suggests the possibility of constructing microecosystems that integrate anaerobic and aerobic niches in one reactor. In this study, we demonstrate that under oxygen-limited conditions, it is possible to construct a single-stage activated sludge bioreactor for removing organic substances and phosphorous simultaneously. These findings have important consequences for recognizing and supplementing the enhanced biological phosphorus removal process and for developing new energy-saving treatment technologies.
Introduction
S
Shen and Guiot (1996) investigated the impact of influent dissolved oxygen (DO) on the characteristics of anaerobic granules at various DO concentrations (0.5–8.1 mg/L) in 1- and 5-L laboratory-scale modified upflow anaerobic sludge bed-anaerobic/aerobic coupled reactors. When the granular sludge was operated under oxygen-limited conditions between 1 and 3 months, there was no detrimental impact of influent DO on the methanogens, and even when the influent DO concentrations were up to 8.1 mg/L, there was no inhibitory effect on acetogens and methanogens. These results show that the anaerobic/aerobic coupled reactor can be successfully operated under O2-limited conditions and can function as an ideal engineered ecosystem that integrates aerobic and anaerobic niches. Compared to the conventional aerobic process, microaerobic systems are more energy efficient. They require less energy for aeration and produce fewer biological solids.
Microaerobic technology has been getting more and more attention. Many researchers have been focusing on the performance of such a system, for example, low DO membrane bioreactor (MBR) (Chu et al., 2006), low DO sequencing batch reactor (SBR) (Díaz et al., 2010), and so on. In previous studies, Zheng et al. (2013) compared the difference of treatment performance and microbial diversity between the upflow microaerobic sludge bed (UMSB) and aerobic control system (ACS) for domestic wastewater treatment. In these studies, we compared the difference of treatment performance and microbial diversity between UMSB and ACS for high organic wastewater treatment. In the present study, we compare the microaerobic and aerobic technologies and also investigate cultivation-independent molecular approaches to study the phylogenetic relationships as well as the diversity of the microbial communities in a microaerobic bioreactor. Cloning and sequencing of polymerase chain reaction (PCR)-amplified 16S rRNA gene fragments have been successfully applied for the analysis of a bacterial community structure in a wide range of environmental samples (Calheiros et al., 2010; Tancsics et al., 2010). In our study, this approach was used to determine bacterial community structure and biodiversity in the UMSB and ACS.
Research into the operational characteristics of wastewater treatment facilities, operating at low DO levels, is of great importance in terms of energy saving and development of novel processes. If such a treatment process can be used to remove organic substances and nutrients, such a process could have important practical benefits. The main objective of this research is to compare the treatment performance of microaerobic and aerobic technology in two parallel single-stage activated sludge bioreactors and to analyze the microbial diversity of UMSB and ACS. It is expected that this study will provide some fundamental information for the recognition and application of microaerobic treatment processes.
Materials and Methods
Synthetic wastewater
Glucose was used as the sole organic source of synthetic wastewater. The synthetic wastewater with the following composition was used: glucose 1.5 g/L; (NH4)2SO4 0.3 g/L; KH2PO4 0.2 g/L; NaCl 0.05 g/L; CaCl2 0.015 g/L; MgSO4·7H2O 0.05 g/L; NaHCO3 0.15 g/L; tap water.
Reactor setup and operation
We used two parallel identical laboratory-scale column-type reactors, comprising a plastic feed tank and a lucite aerated column. The two reactors were provided with a hopper bottom with the following dimensions: length 90 cm, internal diameter 10 cm, and working volume of 6.3 L. The metering pump transferred wastewater from the feed tank to the bottom of the aerated column at a controlled rate. The effluent overflowed from the top of the aerated column and was subsequently discharged. Air was introduced through a porous stone diffuser at the bottom of the reactor.
A 2 g seed sludge was generated by mixing river sediment, field soil, and activated sludge and was placed in the aerated column containing 6.3 L of synthetic wastewater. The mixture was incubated in the batch mode at 25°C±1°C for 2 days to obtain enough biomass before continuous operation. After 2 days, the mixed liquor suspended solids (MLSS) at the bottom of UMSB and ACS were 16.2±1.1 and 9.6±1.5 mg/L, respectively. Then, the synthetic wastewater was prepared on a daily basis, after which it was continuously pumped into the aeration column. The system had a hydraulic retention time of 25 h and an organic loading rate (OLR) of 1.3–1.5 kg COD/m3·d. The DO level of the aeration column was controlled manually and maintained at 2.5–4 mg/L and 0.5–0.9 mg/L in the ACS and UMSB, respectively. The pH was adjusted to 7.0±0.5 by adding 1 M NaHCO3 or 1 M HCl. A heating rod was used to maintain the temperature of the reactor at 25°C±1°C. Water samples and in situ DO and pH measurements, from the upper part of the reactor, were taken on a daily basis.
Analytical methods
COD, total nitrogen (TN), total phosphate (TP), and MLSS were measured according to standard methods (APHA, 2005). DO was determined by a DO meter (HANNA HI 9145). pH was determined by a pH meter (HANNA HI 8124). The measurement sets were individually compared with one-way analysis of variance (ANOVA) using IBM SPSS statistics 20.0 and p<0.05 was considered a significant difference.
DNA extraction and PCR amplification
Wet sludge (0.5 g) was washed thrice, using phosphate-buffered saline (pH 7.0), followed by centrifugation at 4°C, 5,000 rpm for 5 min. The genomic DNA was then extracted using the method of Tsai and Olson (1991). Approximately 10 μg DNA g−1 of wet sludge was obtained using electrophoresis on 1% (w/v) agarose gel and visually compared with a molecular mass ladder. To acquire suitable amplicons, 10–100-fold dilutions of crude DNA were used as templates for subsequent PCR. To minimize PCR bias, three separate reactions were run for each sample and pooled. PCR amplification of the 16S rRNA gene fragments was carried out using the forward primer 27f (5′- AGA GTT TGA TCC TGG CTC AG-3′) and the reverse primer 1492r (5′- GGT TAC CTT GTT ACG ACT T-3′). The conditions for the PCR amplification were as follows: initial denaturation for 5 min at 95°C, followed by 30 cycles of denaturation for 1 min at 94°C, annealing for 1 min at 52°C, extension for 1.5 min at 72°C, and final extension for 8 min at 72°C. PCR products were purified using TaKaPa purification kits.
Cloning and sequencing of 16S rRNA gene
Cloning of amplified 16S rRNA gene fragments into the TA cloning vector PMD18-T was performed following the manufacturer's instructions. Then, DH5α chemically competent Escherichia coli was transformed with the plasmids according to the manufacturer's instructions. Transformants were selected by Luria-Bertani agar plates containing ampicillin (150 μg/mL). Cloned inserts were amplified from lysed colonies by PCR with plasmid vector-specific primers M13F (5′-GTA AAA CGA CGG CCA G-3′) and M13R (5′-CAG GAA ACA GCT ATG AC-3′). Clones were sequenced by Beijing Sunbiotech Co., Ltd.
Phylogenetic analysis
The operational taxonomic unit (OTU) was classified with distance-based OTU and richness determination (MOTHUR) (Schloss et al., 2009). One nucleotide sequence randomly selected from each OTU was BLAST searched against the Ribosomal Database Project (RDP, release 10.0) to identity the taxonomic group. Phylogenetic trees were constructed using MEGA version 4.0 by the Neighbor-Joining (NJ) algorithm, and the Jukes–Cantor distance estimation method with bootstrap analyses for 1,000 replicates was performed. The possible chimeras in the obtained sequences were checked using the BELLEROPHON before phylogenetic analysis (Huber et al., 2004).
Nucleotide sequence accession numbers
Particle 16S rRNA gene sequences that were determined have been deposited in the GenBank, nucleotide sequence databases under accession nos. KC179054-KC179078 and KC305851-KC305875.
Results and Discussions
Operation performance of UMSB and ACS
UMSB and ACS were operated for 45 days. Data on the pollutant concentration in the influent and the effluent, as well as the pollutant removal efficiency during the whole operation period, are presented in Figure 1. These results indicate that, although DO concentrations were very different in the UMSB and ACS, both of the systems indicated good performance in terms of COD removal. The influent COD was 1456±37 mg/L, after 10 days of acclimation, the effluent COD of the UMSB and ACS stabilized at low levels of 50.4–100.8 and 48.0–78.4 mg/L, respectively. The COD removal efficiency stabilized to levels of 94%±1% and 95%±1%, respectively, with no significant difference between these two systems (p=0.498>0.05). This indicates that there was no significant difference in the respiration rate of the organisms between the UMSB and the ACS, which is consistent with results reported by Hanaki et al. (1990).
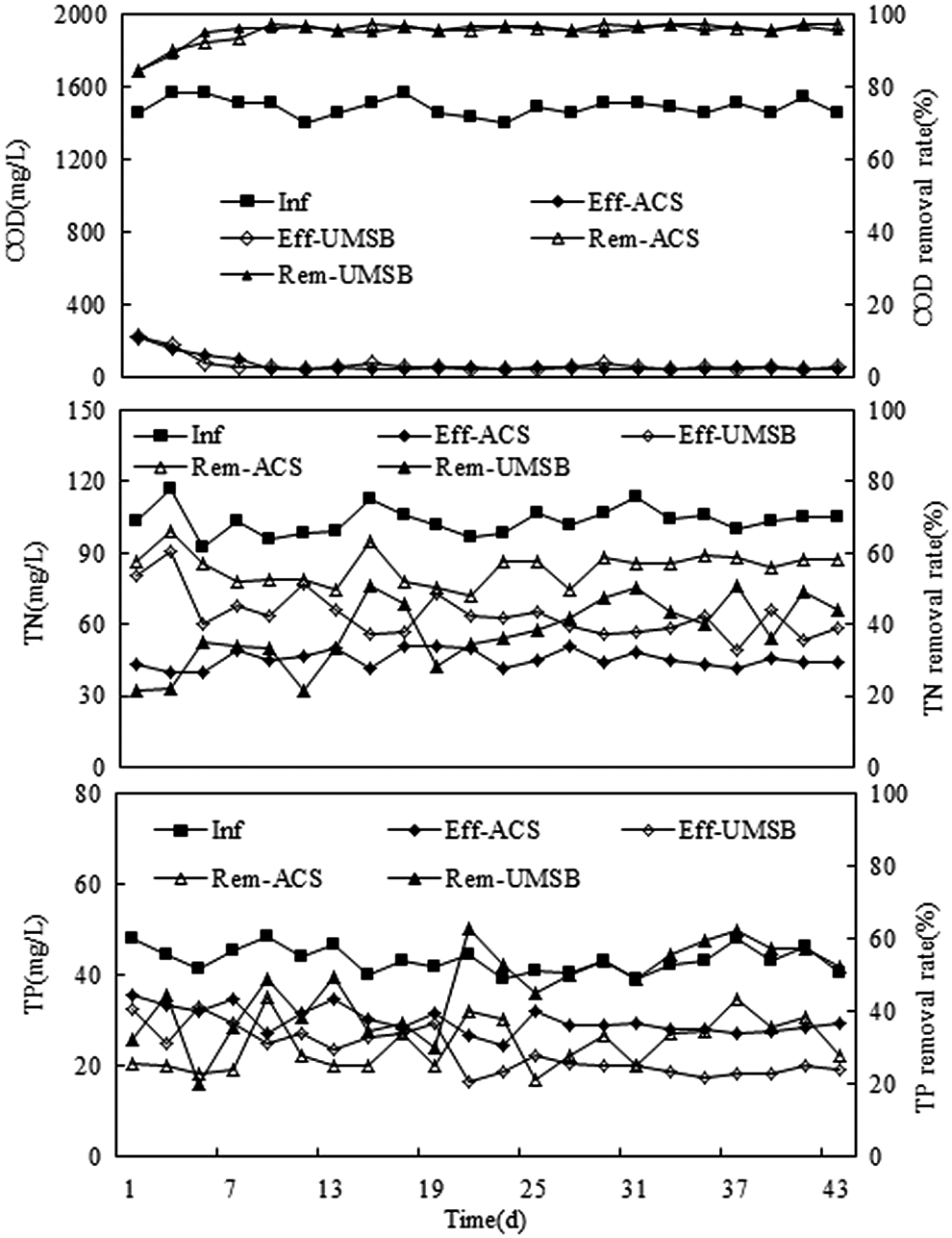
Performance of ACS and UMSB for synthetic wastewater. Inf, influent pollutant level; Eff, effluent pollutant level; Rem, removal rate; ACS, aerobic control system; UMSB, upflow microaerobic sludge bed.
He et al. (2009) also reported that organic substances could be significantly biodegraded in the reactor at DO in the range of 0.8–5 mg/L. Peng et al. (2001) studied simultaneous organic carbon and nitrogen removal in a sequencing batch reactor with a synthetic municipal wastewater feed while being maintained at a low DO level (of 0.8 mg/L). The reactor achieved high treatment capacities (with COD removal efficiency at 95% and OLR reaching 2.4 kg COD/m3·d). In the present study, in which both reactors have high COD removal efficiencies, the results were consistent with the findings listed above. The air flow rates of ACS and UMSB were found to be 36 and 15 L/h, respectively, indicating a 58% saving in aeration costs when using the latter system. It is, thus, clear that operating at low DO levels reduces energy consumption.
During the 45-day operating period, influent TN and TP concentrations were measured at 103±7 and 43±4 mg/L, respectively. After 10 days of acclimation, the effluent TN of ACS and UMSB was 44±3 and 56±8 mg/L, respectively, and the TN removal efficiency of the ACS was higher (56%±3%) compared with the UMSB (43%±6%). At the steady stage, TP concentrations in the effluents of ACS and UMSB were 28±3 and 18±2 mg/L, respectively. The TP removal efficiency of the UMSB was, thus, significantly higher (57%±5%) compared with ACS (34%±6%) (with p=0.00) (Fig. 1).
In recent decades, enhanced biological phosphorus removal (EBPR) in activated sludge systems has become a widely applied wastewater treatment process for the removal of P without the use of chemical precipitation. EBPR can be achieved through the activated sludge process by recirculating sludge through anaerobic and aerobic conditions (Oehmen et al., 2007). Conventional EBPR-based treatment can be extremely efficient, but its requirement for anaerobic pretreatment zones may be problematic when retrofitting existing activated sludge installations. Moreover, the EBPR process may display inconsistencies in performance, since for optimal results, relatively high concentrations of volatile fatty acids (VFAs) need to be present in the influent wastewater (Mullan et al., 2005). For these reasons, the availability of effective yet economical P removal strategies is thus of great importance to the water industry. Mullan et al. (2005) reported that a single-stage aerobic P removal system might have advantages over conventional EBPR technology in terms of (1) rate of throughput, (2) tolerance of high nitrate levels (since the anaerobic phase of EBPR is sensitive to nitrate), (3) reduced dependence upon wastewater strength, and (4) operation at low VFA concentrations. Ju et al. (2005) reported that under microaerobic conditions, the organisms might simultaneously perform aerobic and anaerobic respiration; the fermentation and the aerobic and anaerobic niches might coexist, therefore anaerobes and aerobes are able to survive in such systems. This means that the microaerobic system might function like the aerobic/anaerobic sequential process. In our research, the P removal efficiency of the UMSB was significantly higher compared with the ACS, thus indicating that the EBPR process could function in a single-stage activated sludge bioreactor operating under microaerobic conditions. Besides, in our study, the air flow rate of the UMSB was only 15 L/h, which was much less compared with the ACS (36 L/h), thus indicating that excessive aeration clearly negatively affects the biological phosphorous removal process, a phenomenon that is consistent with the finding of Brdjanovic et al. (1998). With the exception of phosphorus removal, the microaerobic treatment system could be a better alternative for the treatment of many pollutants requiring aerobic/anaerobic sequential treatment. It should also be noted that the UMSB uses less oxygen, which means that energy is saved, thus incurring lower initial costs and operating costs.
Fdz-Polanco et al. (2000) reported that higher COD concentrations limited severely the growth of nitrifying microorganisms being outcompeted; when the influent COD concentration increased to 400 mg/L, only 18% of N-NH4+ was oxidized. In the present studies, the influent COD concentration reached 1,000 mg/L; this condition was not suitable for nitrification and denitrification. Nitrogen source was mainly used in the microbial group.
Microbial diversity and phylogenetic analysis
To obtain more detailed information on the microbial community in the UMSB and ACS, we carried out PCR amplification on partial 16S rDNA fragments from the extracted DNA, using bacterial-specific primer sets. Sequences with ≥98% similarity were combined into 1 OTU using the MOUTHUR software. Thus, 25 OTUs were obtained from 33 UMSB clones and 26 OTUs were obtained from 37 ACS clones. In the case of UMSB, analyzed clones represented nine phyla (Fig. 2): γ-Proteobacteria (4 OTUs, 11 clones), β-Proteobacteria (7 OTUs, 8 clones), α-Proteobacteria (3 OTUs, 3 clones), ɛ-Proteobacteria (3 OTUs, 3 clones), Actinobacteria (2 OTUs, 2 clones), δ-Proteobacteria (2 OTUs, 2 clones), Bacteroidia (2 OTUs, 2 clones), Bacilli (1 OTU, 1 clone), and Flavobacteria (1 OTU, 1 clone). Eight phyla were obtained in ACS (Fig. 3): α-Proteobacteria (8 OTUs, 14 clones), γ-Proteobacteria (2 OTUs, 3 clones), β-Proteobacteria (4 OTUs, 9 clones), Actinobacteria (2 OTUs, 2 clones), δ-Proteobacteria (1 OTU, 1 clone), and Bacilli (2 OTUs, 2 clones). Six additional clones, belonging to the phyla Sphingobacteriia (4 OTUs, 4 clones) and TM7 (2 OTUs, 2 clones), were found in the ACS, which did not appear in UMSB. The phyla of ɛ-Proteobacteria, Bacteroidia, and Flavobacteria were not detected in ACS, but were found in UMSB.
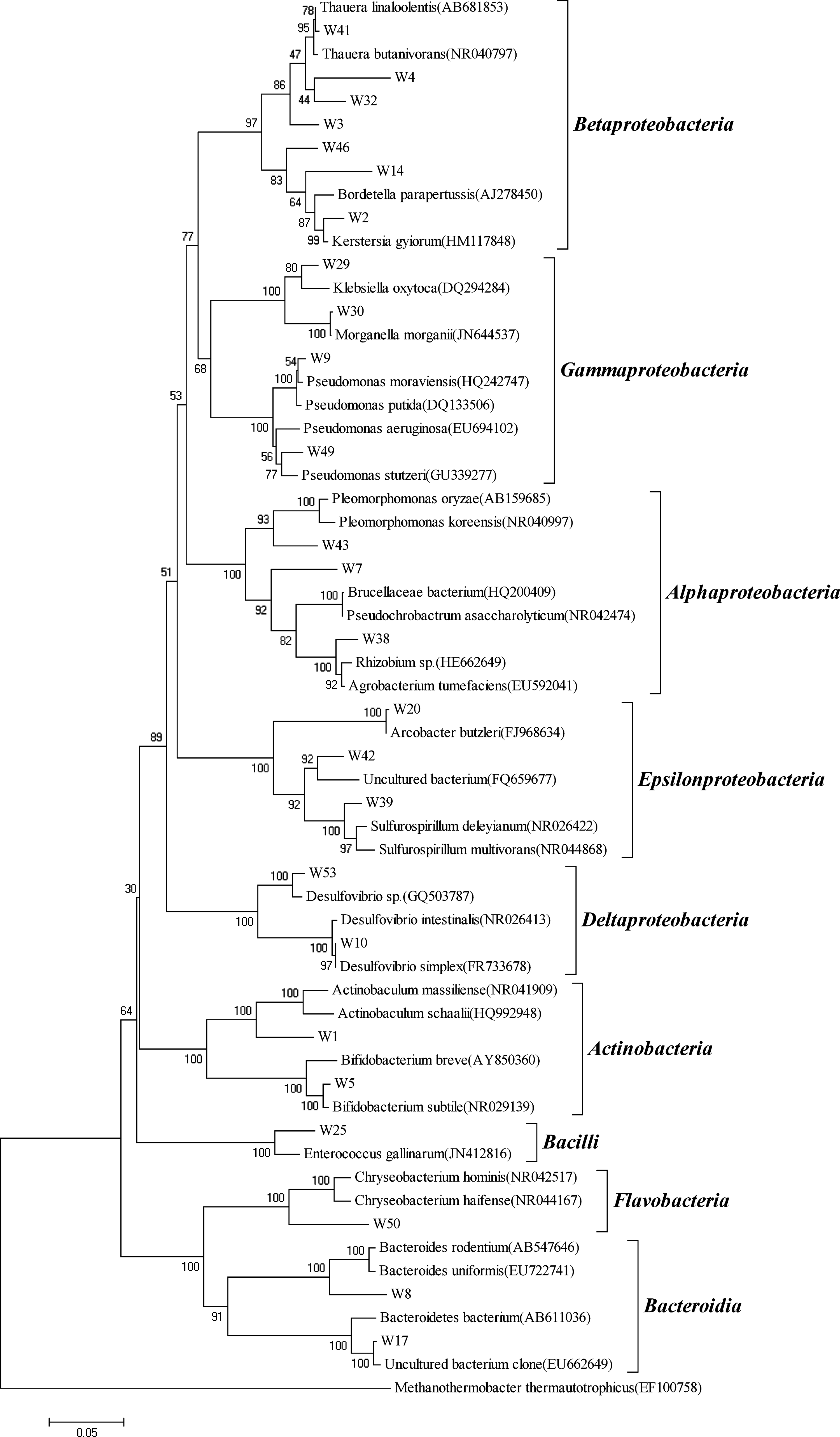
Phylogenetic tree constructed by the Neighbor-Joining (NJ) method and the phylogenetic positions of the clones isolated from UMSB. Bootstrap replication was 1,000. Scale bar estimated difference in nucleotide sequence positions.
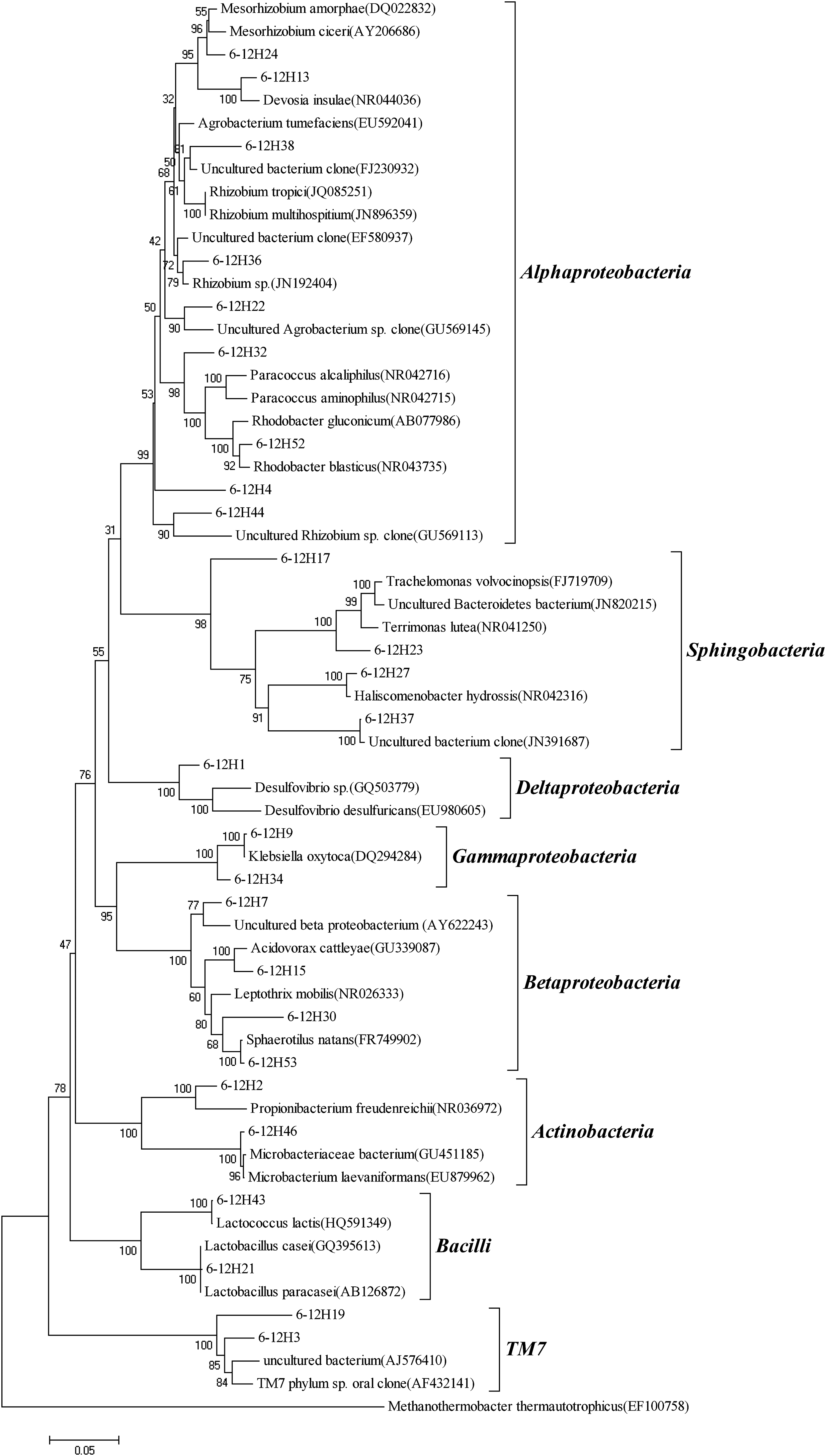
Phylogenetic tree constructed by the NJ method and showing the phylogenetic positions of the clones isolated from ACS. Bootstrap replication was 1,000. Scale bar estimated difference in nucleotide sequence positions.
The phylogenetic tree (Figs. 2 and 3) indicated that bacteria in the UMSB and ACS were highly diverse and obviously different from each other. In the UMSB, γ-Proteobacteria was the most abundant cluster, suggesting that the γ-Proteobacteria was the dominant group in the bacterial population. Sequences belonging to γ-Proteobacteria showed similarities to those of Morganella morganii, Klebsiella oxytoca, and Pseudomonas sp. Of these, M. morganii and K. oxytoca were facultative anaerobic organisms, and Pseudomonas sp. was an aerobic organism. Sequences in the class β-Proteobacteria indicated a relationship to aerobes Bordetella sp. and Thauera sp., and the sequences also showed a relationship to uncultured Candidatus Accumulibacter sp. (JN679133) and C. Accumulibacter sp. (JN679214), which are well-known phosphate-accumulating organisms (PAOs) (Oehmen et al., 2007). Sequences belonging to ɛ-Proteobacteria, Bacteroidia, and Flavobacteria were only detected in UMSB. Sequences belonging to ɛ-Proteobacteria showed relationships to Arcobacter butzleri and Sulfurospirillum deleyianum, which are microaerophilic organisms; Flavobacteria commonly appeared in EBPR processes (Zengin et al., 2011). The phylogenetic analysis results showed that facultative anaerobes, microaerophiles, and aerobes existed in the UMSB. The coexistence of these organisms suggests that microaerobic and aerobic niches might coexist in the UMSB, and that the UMSB might function in a similar way to an EBPR process.
Results illustrated in Figure 2 indicate that none of the sequences detected from the UMSB belonged to the known PAOs. There were numerous factors, for example, such as the application of different carbon sources and the ratios of organic carbon to P in the influent (Ahn and Speece, 2006; Zeng et al., 2006) that would affect the proliferation of PAOs. Numerous studies have indicated that a high COD/P ratio (e.g., >50 mg COD/mgP) in the wastewater feed is favorable for the growth of glycogen-accumulating organisms (GAOs) instead of PAOs. A low COD/P ratio (e.g., 10–20 mg COD/mg P) should be more favorable to the growth of PAOs (Mino et al., 1998). Mino et al. (1998) also noted that competition for the carbon source was the deciding factor affecting the predominance of particular organisms. Zengin et al. (2010) reported that continued glucose feeding favored preferential growth of GAOs over PAOs through the direct glycogen metabolism. In this research, glucose was the sole carbon source and the COD/P ratio in the influent was as high as 29–34 mg COD/mg P, which is not suitable for the growth of PAOs. Besides, the number of the clones is only 33 in UMSB; therefore, the number of the detected clones should be increased significantly or the high-throughput sequencing technique should be used to analyze the microbial diversity in future research work. These might be the reason why no known PAOs were detected from the UMSB. Jeon and Park (2000) reported that EBPR was accomplished by two kinds of microorganisms, a lactic acid-producing organism (LPO) and PAO, when glucose was supplied as a sole carbon source. The LPOs, which rapidly accumulate glucose as glycogen in the cell, are facultative bacteria that produce lactic acid to obtain energy for glycogen storage, which is a different process to that used by other typical acidogenic bacteria for energy production. LPOs could compete with other acidogenic bacteria, since LPOs rapidly accumulate glucose as glycogen before fermentation of glucose by acidogenic bacteria, to produce short-chain VFAs. In this study, there was one clone belonging to Bacilli, which may play the role of LPO.
Research on microaerobic technology often focuses on performance (Chu et al., 2006; Díaz et al., 2010). The present study may, therefore, represent the first report of cultivation-independent molecular approaches for elucidating the phylogenetic composition and the diversity of the microbial communities in a microaerobic bioreactor, compared to that found in a parallel aerobic bioreactor. Microaerobic technology requires advances concerning the identity and biodiversity of the microorganisms involved in these systems, as well as their biochemistry and metabolism. Beyond the phylogenetic analysis, the next challenge is to obtain more information about the genetics and gene expression of the microaerobic-related enzymes.
Conclusions
Results of this research indicate that it is possible to remove organic substances and phosphorous simultaneously in a single-stage activated sludge bioreactor under oxygen-limited conditions. The findings are important in terms of recognizing and supplementing the innovative energy-saving wastewater treatment system, especially the EBPR process. Operation at low DO means energy saving as well as lower initial costs and operating costs. Phylogenetic analysis showed that facultative anaerobes, microaerophiles, and aerobes were able to survive under oxygen-limited conditions. This suggests the possibility of constructing microecosystems, in which anaerobic and aerobic niches are integrated. The microaerobic treatment system might be a better alternative to the aerobic/anaerobic sequential processes. More work is necessary to understand the fundamental biochemical and microbiological mechanisms of microaerobic treatment systems.
Footnotes
Acknowledgment
Our acknowledgement goes to the State Key Laboratory of Water Environment Simulation, School of Environment, Beijing Normal University.
Author Disclosure Statement
No competing financial interests exist.