
Abstract
Abstract
Biomarkers are useful for evaluating exposure and toxic effects of chemicals on invertebrates. Earthworms (Eisenia fetida) were exposed for 28 days to three phthalic acid esters (PAEs), dimethyl phthalate (DMP), diethyl phthalate (DEP), and dibutyl phthalate (DBP), in artificial soil to examine the potential of several biomarkers as indicators of environmental pollution. Survival, growth, sperm count, cocoon production and hatchability, and the lysosomal membrane stability, measured by neutral red retention time (NRR-time), were measured in the earthworm after exposure. DMP, DEP, and DBP showed significant toxicity to earthworm growth, and a 10% reduction in biomass occurred at soil concentrations of DMP 3.48 mg/kg, DEP 5.63 mg/kg, and DBP 4.42 mg/kg. LC50 values of DMP and DEP were 1,560.120 and 1,516.186 mg/kg, respectively, after 14 days of exposure. The LC50 value of DBP could not be determined. The most sensitive biological parameters were reproduction (cocoon production and hatchability) and NRR-time. The number of cocoons was reduced at concentrations above DMP 35 mg/kg, DEP 25 mg/kg, and DBP 20 mg/kg, and cocoons had reduced hatching success at concentrations above 1.5 mg/kg. NRR-time was reduced significantly at exposure concentrations above DMP 250 mg/kg, DBP 150 mg/kg, and DEP 200 mg/kg. These results indicated that soils contaminated with phthalic acid esters can harm or eliminate earthworm populations. Cocoon production rate, hatchability, and NRR-time can be used as biomarkers for monitoring levels of PAE contamination in soils.
Introduction
P
Soil acts as a natural sink for pollutants, which can often transfer to vegetation and enter food chains (Cai et al., 2008). In Chinese soils, polycyclic aromatic hydrocarbons and PAEs are the most abundant semivolatile organic chemicals (Cai et al., 2008; Mackintosh et al., 2004). Along with urban expansion, land use in suburban areas has changed from a single wheat/rice-based mode to a multiform mode. As a result, more organic pollutants, including PAEs, are introduced into the soils from the frequently used agricultural plastic films and other chemicals. PAEs in soil then pose a risk to human health through food chain transfer (Chen et al., 2008; Sun et al., 2013).
Many studies have documented the toxic effect of PAEs on humans and environmental health, especially their endocrine disruption effects (Chen et al., 2011; Wittassak et al., 2011), but little is known about possible effects of PAEs on earthworms. Earthworms are ubiquitously distributed in soils and play a major role in the breakdown of organic matter and the release of nutrients as they transfer decomposed material from the soil surface to subsurface layers (Edwards and Bohlen, 1996). Earthworms are in direct contact with soil pore water and consequently with the bioavailable contaminants in the soil (Chang et al., 2002). Earthworms have been widely used in ecotoxicological assessment of soil (OECD, 1984) and are excellent indicators of soil ecosystem health (Kuhle, 1983). Biomarkers can be defined as any biological responses to an environmental chemical at the individual level or below, demonstrating a departure from the normal status (Walker et al., 2001). Biomarkers have been widely used to examine exposure in laboratory studies with various earthworm species (Spurgeon et al., 2000). Monitoring the growth and development of juvenile Eisenia fetida in contaminated soils provides assessment of contaminant effects. In a polluted environment, changes to individual energy budgets will occur as the organism expends energy resisting the contaminant by avoidance exclusion, removal, or complexing. This additional energy requirement will decrease the scope for growth of the exposed animals, reducing not only growth but also differentiation, cocoon production, and ultimately a number of important life history characteristics, such as time to sexual maturity and generation time (Donker et al., 1993).
In this study, we examined the sublethal toxicity of three phthalic acid esters (DMP, DEP, and DBP) on earthworms by analyzing changes in the survival, growth, sperm count, cocoon production, and hatchability of earthworms in microcosms. We evaluated the usefulness of the biomarker neutral red retention time (NRR-time) as a tool for predicting the effect of PAEs on earthworms and to provide a basis for the study of environmental toxicology in soil ecosystems.
Materials and Methods
Materials and reagents
All the chemicals and solvents used for extraction and gas chromatography–mass spectrometer (GC–MS) analysis were of pesticide residue analysis grade (dichloromethane was supplied by Acros, Avondale, PA). DMP, DEP, and DBP were supplied by Dr. Ehrenstorfer (Augsburg, Germany). All the solutions were used within 6 months of receipt. Sample treatment and data analysis were done as in a previous study (Wang et al., 2006; Zeng et al., 2008; Zeng et al., 2009).
Sublethal toxicity tests with E. fetida
Adult E. fetida (300–600 mg wet weight, purchased from a commercial breeder in Tianjin, China) were acclimatized in clean artificial soil made in accordance with the ISO 11268 (ISO 11268-1, 1993, and ISO 11268-2, 1998) for 14 days. The initial concentrations of DMP and DEP in the soil could not be determined. Tests were conducted using a laboratory culture of the species E. fetida following a modified procedure originally described by Løkke and van Gestel (1998). The residual concentrations of DMP, DEP, and DBP in the soil and bioaccumulation in earthworms at various doses were assessed in preliminary experiments that generated the concentration–response curves for E. fetida. Based on these curves, we selected initial concentrations of DMP, DEP, and DBP to study sublethal toxicity. DMP, DEP, and DBP were dissolved in acetone, and the entire soil batch for a treatment concentration amended with the solution in a ratio of 250 mL acetone solution per 1,000 g of dry soil. The treated soil was left under a fumehead overnight (8 h) to let acetone evaporate before adding the worms. Water was then mixed thoroughly into the soil, and the treated soil was divided into individual replicates. Five hundred grams of dry soil was used per replicate. E. fetida were exposed to the selected concentrations. Four replicates of each concentration and untreated controls were prepared, each containing 10 earthworms. In each container, 28–30 g (wet weight) finely ground manure from nonmedicated cattle was added on the surface of soil as a food source once every 2 weeks. Test containers, cylindrical plexiglass units with inner diameter 6 cm and height 5.5 cm, were kept for 28 days at 20 ± 1°C with 12-h light–12-h dark cycle. At the end of the test, the soil was searched for surviving earthworms. The earthworms were washed, dried, counted, and weighed. The cocoons were collected by water washing in 2-mm mesh, counted, and maintained in Petri dishes with wet filter paper at 20 ± 1°C for additional 28 days to allow hatching. The cocoons were monitored regularly once every 7 days for more than 28 days to evaluate reproductive success.
Growth inhibition
Three initial concentrations of DMP, DEP, and DBP were selected for the test. Earthworms were fed by applying 0.5 g of dry cow manure per worm to the soil surface once every 7 days. The same procedure but with acetone alone was used as a control. After 2, 7, 14, and 28 days of exposure, the earthworms were removed from the substrate, counted, washed in distilled water, dried on paper towels, and weighed. The weights of the earthworms in each dose group after various exposure periods were used to compute growth inhibition as follows:
where GI n is the growth inhibition for dose group n, W0 is the weight on day 0, and Wt is the weight after t day of exposure.
NRR-time and sperm count
NRR-time was measured using the method described by Weeks and Svendsen (1996). Samples of coelomic fluid were collected by inserting a hypodermic needle directly into the coelomic cavity posterior to the clitellum of the worm. Coelomic fluid (20–50 μL) and an equal volume (50 μL) of physiological earthworm Ringer solution were drawn into a hypodermic syringe. The working neutral red concentration (80 μg·m/L) was renewed every hour during the measuring process. The coelomic fluid was placed onto a clean slide and stained with an equal volume of neutral red solution. Each slide was scanned for 2 min at 5-min intervals until 50% of cells were red. This time was recorded as the NRR-time.
Sperm count was determined using the method of Cikutovic (1993). Sperm count and body mass were determined for earthworms after 14 and 28 days of exposure. Five earthworms were removed from each treatment and control. Two counts per earthworm were made. After weighing, earthworms were transferred to dissecting dishes. Seminal vesicles and testes were removed and macerated in 1 mL PBS (pH 7.4). By teasing apart these organs, the sperm was released into the fluid, and sperm was counted. The sperm number corrected by the worm weight (in milligrams) was taken to represent the sperm production of each earthworm. Results were expressed as total sperm counts per milligram for each worm.
Data analysis
DMP, DEP, and DBP were determined with an Agilent 6890N/5975B GC–MS (Agilent Technologies, Avondale, PA), operating in the electron impact and selective ion monitoring mode, with an HP-5MS capillary column (30 m × 0.25 mm and 0.25 μm film thickness) used. The transfer line and the ion source temperature were maintained at 280°C and 150°C, respectively. The column temperature program was initiated at 100°C for 1.0 min, increased to 300°C at a rate of 8°C/min, and held for 3.0 min. The extracts (1.0 μL) were injected into GC–MS in splitless mode with an inlet temperature of 250°C.
Study data were analyzed using SPSS 13.0 (SPSS, Chicago, IL) and Origin 7.0 (OriginLab, Northampton, MA). The critical value for statistical significance was p < 0.05 (SPSS). Estimation of the no-observed-effect concentration (NOEC) and the lowest-observed-effect concentration (LOEC) was done by multiple comparisons using Dunnett's procedure at the 5% significance level. Calculation of effect concentrations at 10% (EC10) and 50% (EC50) for reproduction, growth, and cocoon variability with bootstrapping intervals was done using the ICP approach (Norberg-King, 1993).
Results
Adult survival and growth, sperm count, cocoon production, and hatchability
Figure 1 shows the relationship between E. fetida exposure to different concentrations of DMP and DEP for 7 or 14 days and mortality. LC50 values of DMP and DEP were 1,560.120 and 1,516.186 mg/kg, respectively, after 14 days, but the LC50 value of DBP could not be determined. Figure 2 shows the growth suppression on E. fetida exposed to DMP, DEP, or DBP alone. The influence of compounds on E. fetida was expressed as a growth inhibition (GIn) of E. fetida compared to the control treatment. Earthworm growth decreased with increasing concentrations of DMP, DEP, and DBP. The mean earthworm biomass was significantly reduced (p < 0.05) when exposed to DMP 35 mg/kg, DEP 25 mg/kg, and DBP 20 mg/kg or more. A 10% change in biomass (10% effective concentration: EC10) was estimated at soil concentrations of DMP 3.48 mg/kg, DEP 5.63 mg/kg, and DBP 4.42 mg/kg.
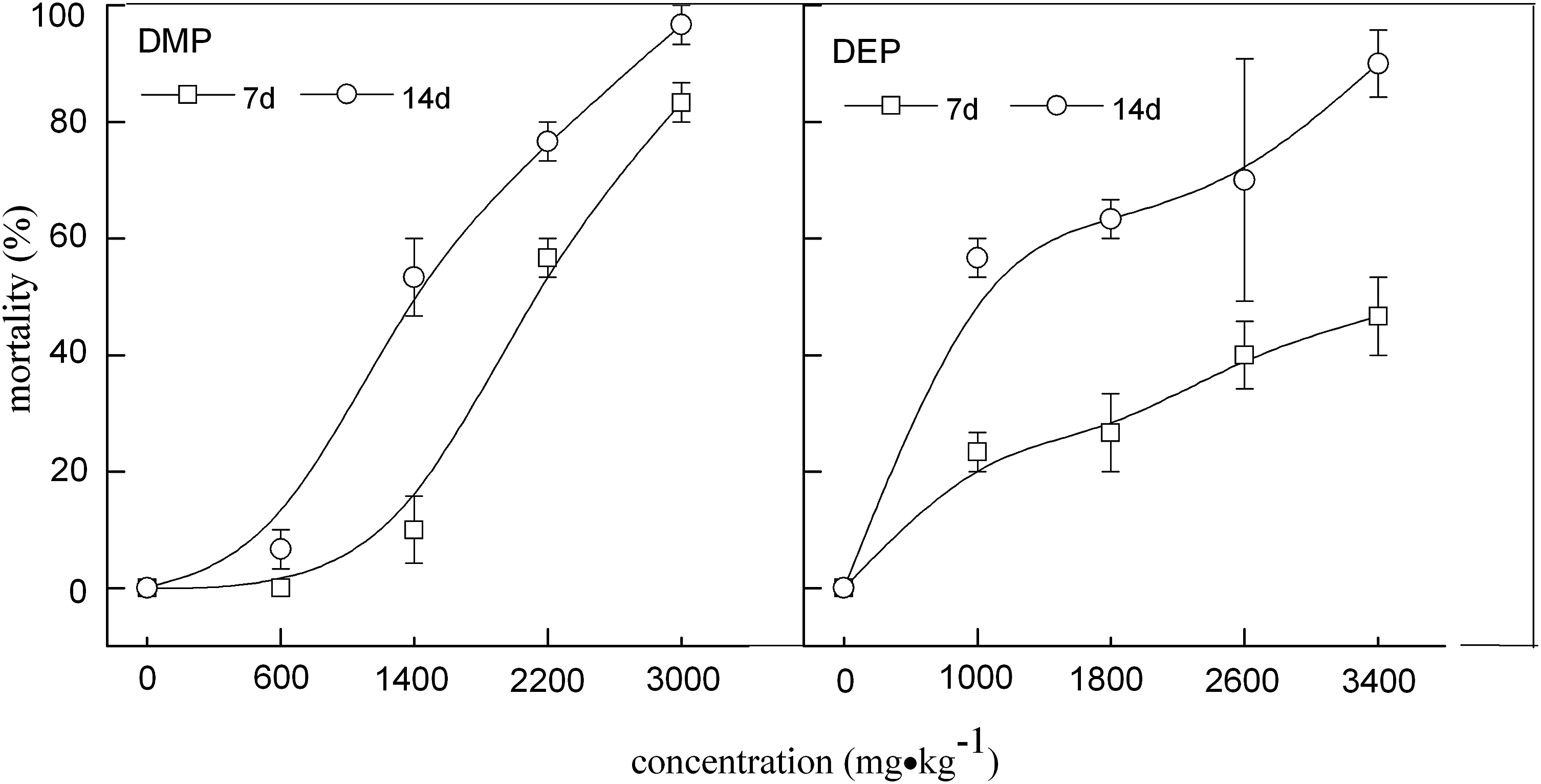
Relationship between mortality of Eisenia fetida and their exposure to different concentrations of DMP and DEP after 7 or 14 days of exposure. Error bars indicate standard deviation. DEP, diethyl phthalate; DMP, dimethyl phthalate.

Relationship between growth inhibition rate of E. fetida and their exposure to different concentrations of DMP, DEP, and DBP after 2, 7, 14, and 28 days of exposure (*p < 0.05, **p < 0.01, ***p < 0.001). DBP, dibutyl phthalate.
After 28 days of exposure to different concentrations of DMP, DEP, and DBP alone, the number of sperm per milligram of body mass did not change significantly (Table 1). However, in worms exposed to DMP, DEP, and DBP, cocoon production was reduced with increasing concentrations (Table 1). No cocoons were produced at the LC50 test concentrations (DMP 1,560 mg/kg, DEP 1,516 mg/kg, and DBP 1,000 mg/kg). The LOECs for cocoon production were DMP 20 mg/kg, DEP 15 mg/kg, and DBP 10 mg/kg (Table 2). The mean cocoon weight ranged from 8.55 to 12.17 mg and showed no significant change within the tested soil concentrations. The number of surviving juveniles showed a clear dose–response relationship with concentration. The highest number of hatchlings was found in the control group, and significantly fewer juveniles were found at test concentrations above DMP 35 mg/kg, DEP 25 mg/kg, and DBP 20 mg/kg or more (Table 2). The mean hatching successes of cocoons in the control group were 81.13% compared to <50% at DMP 250 mg/kg and DBP 150 mg/kg and <40% at DEP 200 mg/kg (Table 1).
DEP, diethyl phthalate; DMP, dimethyl phthalate.
Based on 95% significance levels.
Confidence intervals enclosed are bootstrapping values (equivalent to a 95% significance level).
Attention is drawn to the fact that EC10 values are extrapolated beyond the lowest test concentration.
LOEC, lowest-observed-effect concentration; NOEC, no-observed-effect concentration; NRR-time, neutral red retention time.
Effect of DMP, DEP, and DBP on NRR-time
Figure 3 shows the NRR-time (in minutes) for earthworms exposed to different concentrations of DMP, DEP, and DBP for 7, 14, and 28 days under laboratory conditions. The NRR-time for earthworms was reduced with increasing concentrations of DMP, DEP, and DBP. The NRR-time for the control groups was >80 min, while the NRR-time for earthworms exposed to DMP 720 mg/kg, DEP 630 mg/kg, and DBP 540 mg/kg or more was <60 min. NRR-times were significantly affected by the exposure of PAEs at all concentrations. The EC10 and EC50 for the NRR-time were extrapolated to DMP 3.72 and 14.68 mg/kg, DEP 5.65 and 17.64 mg/kg, and DBP 5.25 and 15.41 mg/kg, respectively. The NOEC values were less than DMP 20 mg/kg, DEP 15 mg/kg, and DBP 10 mg/kg (Table 2). The effects on membrane stability following DMP, DEP, and DBP exposure were highly correlated with the number of cocoons produced (DMP r2 = 0.947, DEP r2 = 0.898, and DBP r2 = 0.918) (data not shown).
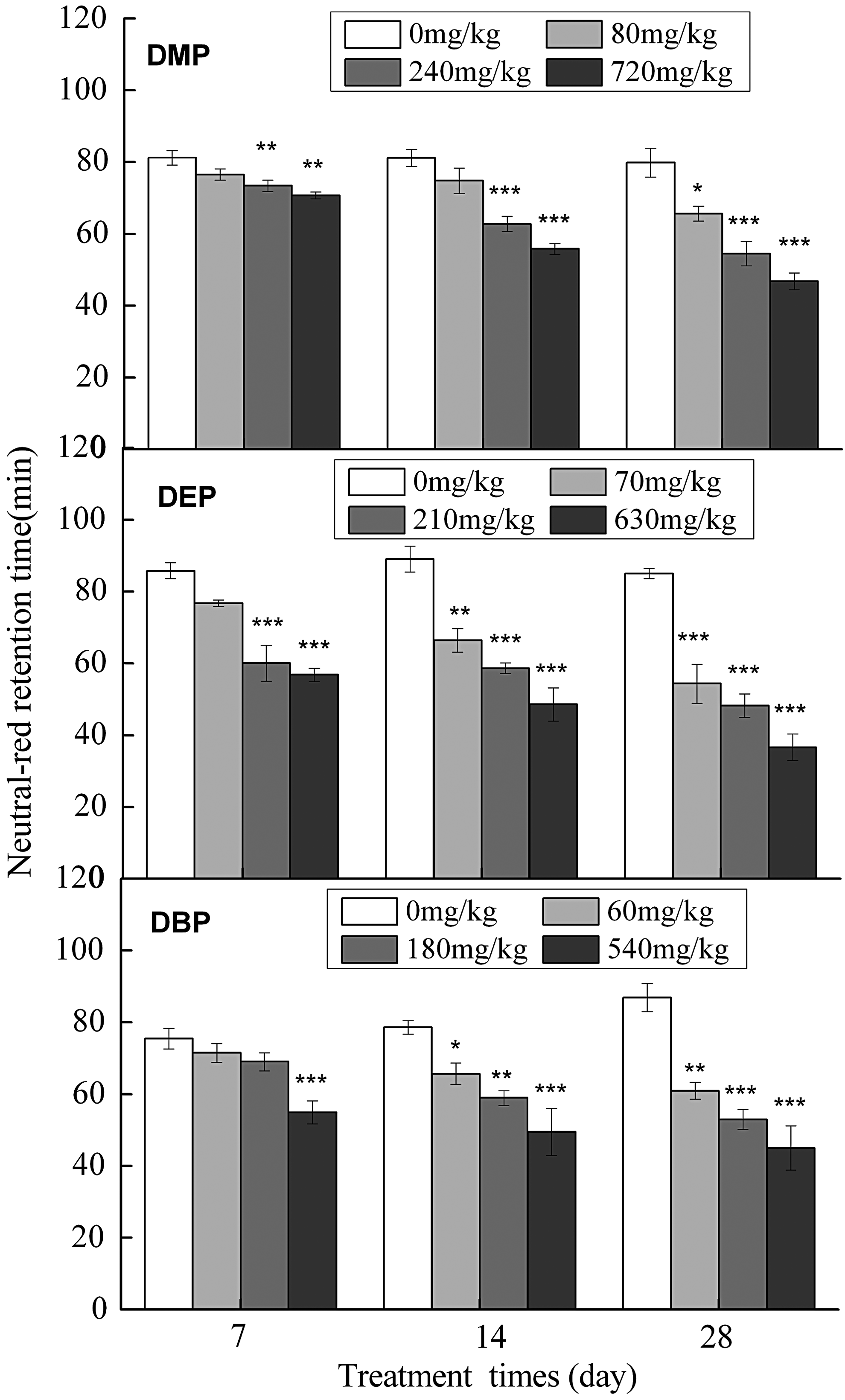
Neutral red retention times (minutes) for earthworm (E. fetida) exposed to different concentrations of DMP, DEP, and DBP for 7, 14, and 28 days under laboratory conditions (statistical significance vs. control group: *p < 0.05, **p < 0.01, ***p < 0.001).
Discussion
This is the first report of PAE toxicity to earthworms. Cocoon production, hatching, and juvenile growth rate and sexual development are important earthworm life history parameters since they can modify field population dynamics. Our results indicate that juvenile E. fetida are more sensitive to PAE-contaminated soils than mature worms as significantly increased mortality was found in the most heavily contaminated soils. This was similar to the report of Donker et al. (1993). High levels of a pollutant can alter growth by both direct and indirect effects. Growth can be impaired by direct toxic effects on worms or by changes in the energy budget as an individual attempts to prevent accumulation in sensitive tissues. Disruption of the energy budget in the worms exposed to contaminated soils could also cause differences in sexual maturation, cocoon production, and hatchability. Energy is required for maintenance, growth, differentiation to reach maturity, and egg production.
Previous studies demonstrated that cocoon production rates were particularly sensitive during the early period of reproduction. However, our study showed that the DMP, DEP, and DBP exposure did not depress sperm counts after 28 days of exposure, but there was a reduction in cocoon production with increasing concentrations. The number of cocoons was reduced at concentrations above DMP 35 mg/kg, DEP 25 mg/kg, and DBP 20 mg/kg, and cocoons exhibited a reduced hatching success at concentrations above 1.5 mg/kg. The number of surviving juveniles was inversely related to the exposure concentration. The highest number of hatchlings was found in the control group, and significantly fewer juveniles were found at all test concentrations above DMP 35 mg/kg, DEP 25 mg/kg, and DBP 20 mg/kg or more. Mean cocoon size increased slightly with exposure to contaminated soil. It is not clear if this relationship is a response of earthworms to the presence of high metal levels or if cocoons are larger due to the decrease in cocoon production rates in the heavily contaminated soils.
High PAE levels in soils used for vegetable production have been reported in China, such as Shenzhen in Guangdong and Handan in Henan (Cai et al., 2005). These were all related to the use of plastic film mulch. In film-covered soils, PAE contents were 208% higher than those for vegetable soils where no film was used. In farmland soils, DMP, DEP, DEHP, and DOP concentrations in film-covered soils were 9.7%, 26.9%, 13.8%, and 587% higher, respectively, than in soils where film was not used (Kong et al., 2012). E. fetida does not occur naturally in the soils of this region; however, this earthworm is a suitable model for predicting the effects of PAEs on populations of other earthworm species.
It could be argued that the plastic film used in Chinese agricultural soils should be immediately banned or gradually replaced considering the resulting PAE pollution. Chen et al. (2011) showed that vegetable production soils using plastic mulch for long periods had relatively higher PAE content. Within the sample areas, both vegetable fields and noncultivated fields were used to cultivate vegetables, and the levels of DMP and DEHP were similar. These observations suggest that the PAEs are at least partly derived from human activity.
Treated biosolid waste sludge is commonly used for soil conditioning (biosolids represent half of European sludge production), and the resulting accumulation of persistent toxic organic compounds, as PAEs, in the soil poses a threat to ecosystems and human health. The use of sewage sludge in agriculture can produce human exposure during its application or through the resulting introduction of these compounds into the food chain. There is an urgent need to ensure that sludge is free from contaminants before use (Amir et al., 2005). Some PAEs and their metabolites have possible endocrine disruption effects, such as interference with reproductive physiology in mammals (Harris et al., 1997). Potential health effects in humans are reduced fertility and postnatal development alterations (Ema et al., 2008). For instance, an in vivo study on rats demonstrated adverse effects of BBP on testes and sperm production (Sharpe et al., 1995). Another multigenerational study investigated the reproductive toxicity of dibutyl phthalate (DBP) in rats (Harris et al., 1997). There are many publications related to the impact of PAEs on human health but few reports on the lethal effects of PAEs on earthworm populations. More research is needed to clarify the toxicity of these pollutants.
The present toxicity study did not follow the international standard test protocols (OECD or ISO) for sublethal effects on earthworms. We used a natural agricultural soil with a lower organic matter content that differed from the OECD standard soil. In this agricultural soil, the absorption of PAEs might be higher, and hence, the toxicity is higher compared to a similar test with OECD soil. The quantity and the quality of organic matter play a significant role in the sorption of organic chemicals to soil particles (Huang et al., 2003). We showed sublethal effects of three PAEs (DMP, DEP, and DBP) on adults at significantly lower levels than those causing mortality. The most sensitive endpoints were growth and hatchability of cocoons (Table 2).
Neutral red staining is an effective technique for evaluating cell toxicity. It has been used to test the cytotoxicity of pharmaceuticals, surfactants, food additives, pesticides, solvents, complex chemical mixtures, and a variety of chemical agents using cells from mammals, humans, and fish as targets. PAEs are chloride channel inhibitors, and they may have direct effects on membrane stability. The biomarker NRR-time assay may be useful for determining earthworm toxicity. We found that the lysosomal membrane stability was reduced with increasing concentrations of DMP, DEP, and DBP, and there was a clear dose–response relationship. Membrane stability alteration following DMP, DEP, and DBP exposure was highly correlated with the reproduction of earthworms as was also observed by Wu et al. (2012) using phenanthrene and pyrene. Earthworms were under physiological stress at a field dose of DMP 80 mg/kg and DEP and DBP 60 mg/kg, and NRR-time provided a sensitive predictor of earthworm stress related to PAE exposure. Relatively large variation between different individuals at the same exposure concentration was, however, observed in some cases. The NRR-time assay technique is lengthy and must be performed on fresh and living cells, so only a limited number of samples can be analyzed. Future studies with the NRR-time assay would benefit using more test samples to decrease the variability. Reproduction is a crucial component of earthworm population dynamics, so NRR-time could be used to estimate toxic effects on field populations of earthworms. However, the natural fluctuation in the NRR response and the sensitivity to stress factors, such as pH, soil type, temperature, and drought, should be further investigated before the biomarker is applied as a field-based monitoring tool, although many of these have been observed to be of little importance.
The initial concentrations of PAEs in soils from the Hangzhou Bay region are likely to exceed the highest observed effect concentration of 4.2 μg/g found in this study, so PAEs could potentially affect earthworm populations. Therefore, the immediate risk of PAEs to soil and soil-dwelling invertebrates is probable. More detailed field and laboratory studies are needed to determine the specific environmental risks of PAEs. From the assessment of the field populations of E. fetida, this species appears to exhibit greater tolerance to PAEs than other earthworm species. Avoidance behavior of the worms could be explained through their ability to sense PAEs, thus exhibiting a survival advantage in the environment. This phenomenon has been shown with the nematode Caenorhabditis elegans, which exhibits avoidance behavior mediated through multiple neural pathways (Sambongi et al., 1999). Our study of E. fetida and their avoidance of PAE-contaminated soil provide further evidence that PAE residues are toxic.
Conclusion
Earthworm populations were adversely affected following exposure to PAEs at environmentally relevant concentrations. The earthworm neutral red retention assay was sufficiently sensitive for measuring the sublethal effects of PAEs. Data from juvenile development experiments should be considered when proposing a safe environmental concentration for a pollutant. The cocoon production rate and hatchability and the NRR-time could be useful as novel bioassays for monitoring the levels of PAE contamination in soil ecosystems.
Footnotes
Acknowledgments
This study was supported by the Natural Science Foundation of Zhejiang Province, China (grant nos. LQ14C170002 and LY13B070008). We would also like to thank professor Weihong Zhong for the helpful discussions and the good working conditions.
Author Disclosure Statement
No competing financial interests exist.