
Abstract
Abstract
This study investigated the influence of light availability and water composition on biofilm growth. To this end, the formation of an epipsammic biofilm on riverbed sediments was monitored during 21 days. Experiments were carried out in two specifically designed experimental channels: channel 1 fed with river water and channel 2 with nutrient-enriched input. Each channel was divided into three sections receiving different light intensities. Total and biologically active organic carbon (BAOC), chlorophyll a and b, total carotenoids, soluble carbohydrates (SC), proteins, phosphatase, and bioavailable phosphorous were determined in sediment samples. In channel 1, fed with river water, a positive effect of light availability on chlorophyll a, total carotenoids, total and BAOC, crude proteins, and phosphatase activity was observed throughout the experiment. In channel 2, addition of the nutrients increased the concentrations of chlorophyll a, SC, and proteins, in comparison with the sections receiving the same light in channel 1. These properties also increased with light availability. This study demonstrated that epipsammic biofilm formation in mesocosm conditions depends on light availability and overlying water composition. Designed experimental fluvial channels fed only with river water can be employed to obtain epipsammic biofilm for use in environmental and biotechnological experiments, thus avoiding the affectation of competitor aninons or high ionic strength due to the presence of supplementary nutrients.
Introduction
T
Biofilm biomass and composition may vary in response to environmental factors. Irradiance, temperature, water flow, and nutritional status constitute environmental key factors for the growth of phototrophic biofilms, and this is demonstrated by their effects on photosynthetic and respiratory activity, and on the production of extracellular polymeric substances (EPS) (Zippel et al., 2007). In riverine systems, Villeneuve et al. (2011) found that bacterial and algal densities were highly dependent on seasonal factors (temperature and light availability) and chemical water quality. As for light availability, it is known that photosynthetically active radiation is essential for photosynthesis to store energy and reductive capacity for primary production (Zippel et al., 2007). Consequently, a positive effect of light availability on biofilm algal growth has been described by Guasch and Sabater (1994) and by Hill (1996), among others. As for nutritional status, it has been observed that an increase in inorganic nutrients (mainly nitrate and phosphate) favors algal growth as proved by increased algal biomass and chlorophyll concentrations (Perrin et al., 1987; Borchardt, 1996; Dodds et al., 1997, 2002; Tank and Dodds, 2003). Previous work by Devesa-Rey et al. (2009) showed that phytopigment concentration in the surface sediments along the Anllóns River watercourse was directly related to the concentrations of organic matter (OM), total N, total phosphorous (P), and available P in sediments.
Mesocosm experiments conducted in experimental channels make it possible to carry out ecosystem-level research under controlled conditions (Roussel et al., 2007). Mesocosm studies have greater ecological relevance than small-scale laboratory experiments, although less experimental control and replication (Clements and Neuman, 2002). In recent years, experimental fluvial channels have been used to study the growth of freshwater and marine phototrophic biofilms, using microscope slides as a substratum and characterized by gravimetry, microscopy, taxonomy, molecular biology, and chemical analysis (Zippel et al., 2007), to investigate the effect of biofilms on the chemical processes in sediments (Woodruff et al., 1999; Gainswin et al., 2006; Serra et al., 2009a), to determine the influence of environmental conditions and toxic substances (metals, herbicides, and bactericides) on the biofilm (Guasch et al., 2004; Guasch et al., 2007; Navarro et al., 2008; Ricart et al., 2009; Serra et al., 2009b; Ricart et al., 2010; Serra et al., 2010; Bowes et al., 2012; Bonnineau et al., 2013; Proia et al., 2013), to study the interaction between toxicants and grazers (Muñoz et al., 2001; Real et al., 2003; López-Doval et al., 2010), and also to evaluate the effect of biofilms on sediment stability (Stone et al., 2011) and on the toxicological effects of resuspended pollutants (Brinkmann et al., 2010). However, the use of experimental indoor fluvial channels to study the growth of biofilms developed on sediments is scarce and mainly focused on the study of the effects of biofilms on chemical processes in sediments, as in Woodruff et al. (1999) and Gainswin et al. (2006).
Chemical and biochemical methods can be used to follow the growth of the epipsammic biofilm. Given that algae cells, together with the EPS matrix, comprise the majority of freshwater biofilm biomass in good lighting conditions—bacterial biomass is usually lower—(Romaní, 2010), phytopigments and EPS concentrations may be suitable parameters to evaluate the development of biofilms. Chlorophylls, together with carotenoids, are the main photosynthetic pigments, whereas chlorophyll-a can be found in all photosynthetic algae, chlorophyll-b can only be found in green algae and Euglenophytes (Leavitt and Hodgson, 2001). In turn, the EPS matrix is a crucial structural parameter for biofilm integrity, stability, and architecture (Hamilton, 1987; Romaní et al., 2004). Polysaccharides are the main components of the EPS matrix (Sutherland, 2001) and can make up between 50% and 90% of their total OM (Denkhaus et al., 2007). Proteins, nucleic acids, (phospho)lipids, and humic substances have also been identified in biofilms (Flemming and Wingender, 2003). Sutherland et al. (1998), studying the effect of biofilms on the erodibility of sublittoral sediments, stated that soluble carbohydrates (SC; also called colloidal carbohydrates) are considered a measure of EPS or mucopolysaccharides secreted by diatoms. However, the analysis of total carbohydrates by the classical phenol-sulfuric acid assay (Dubois et al., 1956) includes intracellular, extracellular, and particle-bound carbohydrate material (Underwood et al., 1995) and does not provide a good estimation of EPS.
This study was conducted to (a) evaluate the influence of light availability and trophic state on the development of biofilms; (b) test the suitability of experimental channels to grow biofilms on fluvial sediments, which may be used to perform subsequent environmental and biotechnological studies; and (c) investigate whether biofilms can grow in channels fed with river water, without the addition of supplementary nutrients. The latter is a relevant point because subsequent studies, such as contaminant adsorption, may be affected by the presence of competitor anions or high ionic strength.
Biofilms growing directly on sediments reproduces the conditions of river ecosystems and appears appropriate to be used in mesocosm studies with environmental or biotechnological purposes. The information from this study will also increase the knowledge regarding biofilm development in natural streams, its variation under different light regimes, and the influence of the overlying water composition, which may be affected by hydrological conditions and soil use in the basin.
Material and Methods
General description of riverbed sediments
Riverbed sediments employed for this study were collected in the Anllóns River (NW Spain), a rural catchment of 516 km2 with a history of agricultural, forestry, and cattle rearing activities. During the Roman Empire, gold mining became an active industry that continued until the 20th century. Various studies have been conducted on the composition of the Anllóns River sediments (Devesa-Rey et al., 2008; 2010a; 2010b; Iglesias et al., 2011), and on the development of biofilms along the river course (Penalta-Rodríguez et al., 2008; Devesa-Rey et al., 2009; Sanmartín et al., 2011). Bacillariophyceae (diatoms) comprise up to 98% of the autotrophic population at the sampling stations, whereas the heterotrophic population evaluated by the most probable number (MPN) reaches values of up to 1.2 106 MPN g−1 (Devesa-Rey et al., 2010c).
Bed sediment was collected over metamorphic rocks (peridotite, serpentinite, and piroxenites) at Ponte de Eguas (43° 13′ 24.46″ N 8° 45′ 44.61″ W), located 8 km downstream from the town of Carballo (with a population of over 25,000). A complex sample was taken with a small plastic shovel from the top 4 cm, picking sediment at various points at this site. The 4 cm deep layer is considered the active layer under the water–sediment interface, suffering physical and biochemical exchanges between sediments and water. This is also the depth where biofilms generally develop (Moreno and Niell, 2004). The sediment sample was transported under a water layer in thermally insulated airtight plastic boxes to avoid oxidation. Once in the laboratory, the sample was stored wet at 4°C until deposited in the experimental channels forming a layer of about 3–4 cm. The main characteristics of the sediment are as follows: 86.2% sand fraction, 7.1% silt fraction, and 6.7% clay fraction; 13.9 g/kg OM; 629.6 mg/kg total nitrogen (N); 471.9 mg/kg total P; pH 5.5 (sediment:water ratio 1:2.5); and 51.9 g/kg total Fe.
Experimental design for mesocosm study
Biofilm formation was monitored in two experimental indoor fluvial channels, which were 130 cm long, 13 cm wide, and 10 cm high (Fig. 1a). The lighting system consisted of five lamps (Mazda Fluor Lumière du Jour C9 TF 65 85 W; Philips, Amsterdan, Holland). Each channel was divided into three sections (1, 2, and 3), with different light intensities (40, 30, and 20 μmol photon m−2/s, respectively), obtained by shadowing with one and two sheets of semi-translucent paper laid over the sections 2 and 3, respectively (Fig. 1b). Light intensity was measured by a portable lux meter (photo-radiometer) Model HD 2302.0 (Delta Ohm, Padova, Italy). Experiments were run with a 16:8 h light:dark cycle.
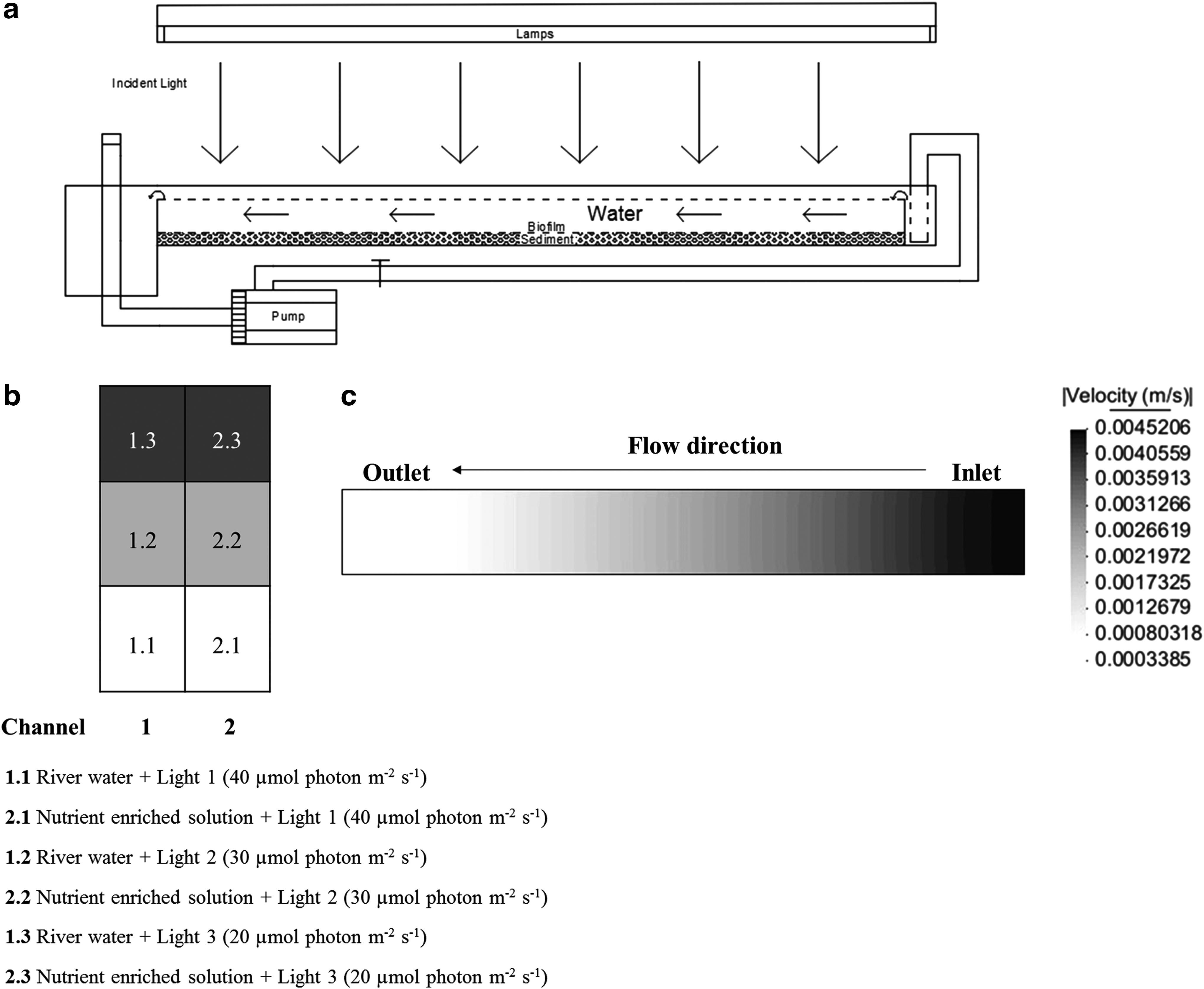
Experiments were carried out with laminar flow achieved using an individual pump system model 1260–210 (Eheim, Deizisau, Germany), with a maximum capacity of 40 L/min. The velocity profiles along the experimental channels have been modeled employing IBER v.2.3 (Bladé et al., 2014), a two-dimensional depth-averaged mathematical model for the simulation of free surface flow in rivers, estuaries, and channels developed by the Water and Environmental Engineering Group, GEAMA (University of A Coruña) and the Flumen Institute (UPC-CIMNE) (Fig. 1c). Selected Manning's Roughness Coefficients for bottom surface (sand/clay) and for channel walls (steel) were 0.023 and 0.011, respectively. Max. water velocity reached was 4.5 10–3 m/s and Reynolds number max. was 178.1. Channel 1 was fed with river water (pH = 7.47, electrical conductivity (EC) = 105 μS/cm, total alkalinity = 12.5 mg CaCO3 L−1, N = 1.98 mg/L, TOC = 1.25 mg/L, Cl− = 10.69 mg/L, NO3− = 7.32 mg/L, PO43− = 0.10 mg/L, SO42− = 5.80 mg/L, Na+ = 6.77 mg/L, K+ = 0.81 mg/L, Ca2+ = 7.41 mg/L, Mg2+ = 2.02 mg/L, Al3+<2.0 μg/L, Fe3+<2.0 μg/L) and channel 2 with the following nutrient input: (pH = 8.41, EC = 28.80 mS/cm, N = 5.69 g/L, TOC = 4.32 mg/L, Cl− = 15.17 mg/L, NO3− = 26.40 g/L, PO43− = 1.90 g/L, Na+ = 9.67 g/L) prepared in distilled water, according to the composition previously optimized by Devesa-Rey et al. (2010c). Evaporative water loss (6% of the circulating water per day), previously determined following the increase in concentration of 1 mM sodium chloride solutions incorporated to the experimental fluvial channels and corroborated by theoretical calculation by Stefan law integrated developed by Holman (1999), was corrected daily to keep constant the volume of the overlying water.
Biofilm development was monitored during 21 days. Every 3–4 days, approximately 50 g of the superficial (∼0.5 cm) sediment was collected by taking samples at various points in each section. Aliquots were centrifuged at 2,000 rpm to discard pore water and the solid sample used to determine phosphatase activity. The remainder of the samples were freeze-dried and stored in polyethylene bottles for later analyses. Water content (% w/w) was determined to calculate the results on a dry weight basis. All the determinations were carried out in triplicate.
Analytical procedures
Total organic carbon (OC) was determined by wet oxidation with K2Cr2O7 and H2SO4, followed by titration with Mohr's salt, according to the procedure proposed by Sauerlandt and modified by Guitián and Carballas (1976), using an automatic titration system. Biologically active OC (BAOC) was determined as per Weill et al. (2003), based on the oxidation of the OM with 0.2 M KMnO4.
Total N was determined by wet digestion with H2SO4, following the Kjeldahl method as described in Guitián and Carballas (1976). Crude protein (PROT) contents were indirectly determined from total N content using an N-to-protein conversion multiplier. In this case, the classic factor of 6.25 (Jones, 1931) may not be appropriate, because it could overestimate the actual protein content (Ezeagu et al., 2002) to include nonprotein nitrogenaceous substances such as pigments, nucleic acids, free amino acids, and inorganic nitrogen (Barbarino and Lourenço, 2005). Hence, a factor of 4.78, proposed by Lourenço et al. (2004) for 12 marine microalgae species in different growth phases, was applied in this study.
Phytopigments, including chlorophyll-a (Chl a), chlorophyll-b (Chl b), and total carotenoids (CAR), were solubilized after 46 min of extraction with dimethylsulfoxide, using an extractant:sediment ratio of 3.6 mL/g at 57°C, following the method optimized by Devesa et al. (2007). In the extracts, Chl a, Chl b, and CAR were determined spectrophotometrically (UV Visible Spectrophotometer; Cary 100 Conc., Varian, Inc., Palo Alto, CA) following the methodology proposed by Wellburn (1994). Equations 1–3 were used to determine the concentrations of Chl a and b, and CAR, in μg/mL.
where Chl a and Chl b represent the concentration of chlorophyll-a and chlorophyll-b respectively, and CAR represents the concentration of total carotenoids, which comprises the oxidized forms, xantophylls, and reduced forms, namely the carotenes. A665.1, A649.1, and A480 represent the absorbance of the extracts at 665.1, 649.1, and 480 nm, respectively. Each sample was submitted to several extractions until no phytopigments were quantified in the extracts, and the concentrations obtained were added to obtain the total concentration.
SC were extracted in water following the method described by Underwood et al. (1995) for the measurement of microbial carbohydrate exopolymers from intertidal estuarine sediments, and subsequently quantified in the aqueous extracts by the traditional phenol-sulfuric acid method (Dubois et al., 1956). D(+) glucose solutions were used as standards, carbohydrate absorbance was measured at 485 nm (UV Visible Spectrophotometer; Cary 100 Conc. Varian, Inc., Palo Alto, California) and the results are given in milligram glucose equivalents per gram of DW sediment.
Activity of the enzyme acid phosphatase (APHOS) was determined using p-nitrophenyl phosphate as a substrate, according to the procedure described by Dick et al. (1996). The intensity of the yellow color due to p-nitrophenol in the filtrate was measured using a UV-Vis spectrophotometer at 410 nm and the results were expressed as milligram p-nitrophenol per kilogram of DW sediment. Bioavailable P (BioP) was extracted with NaHCO3 0.5 N (Olsen and Sommers, 1982), followed by colorimetric determination with molybdenum blue, as described by Murphy and Riley (1962).
Results and Discussion
Influence of light availability on freshwater biofilm growth
Figure 2 shows the time evolution of OC (a) and BAOC (c) contents in channel 1, supplemented with river water. Different light intensity applied to each section (40, 30, and 20 μmol photon m−2/s for sections 1, 2 and 3, respectively) are indicated by the gray scale, where the darkest gray indicates the least illuminated section. In section 1, receiving the highest light intensity, OC increased from an initial value of about 0.70%, to 1.56% at day 11. Then, a subsequent decline until day 21 (1.09%) was observed. In sections 2 and 3, the maximum OC contents were reached at day 18 (1.32% and 1.28% respectively). Sections 1 and 3 (the most and least illuminated, respectively) always showed differences between their OC contents, proving the influence of light availability on biofilm growth. BAOC showed less variation and did not have a clear maximum throughout the experiment. Its values ranged between 405.29 and 724.01 mg/kg, representing between 3.4% and 11.1% of OC. This active fraction of the OM has not been accurately defined in aquatic ecosystems. Some authors described BAOC as a mixture of simple carbohydrates, proteins, and fatty acids (Fabiano and Danovaro, 1994; Fabiano et al., 1995), or as the fraction of OM that is bioavailable to benthic organisms (Polymenakou et al., 2007). In any case, it has been observed that BAOC is closely related to substrate-induced respiration, basal respiration, microbial biomass, and SC, which are all evidence of biological activity (Weill et al., 2003). Thus, the highest BAOC contents observed in the most illuminated section 1 can be suggestive of a positive influence of light availability on biofilm growth.
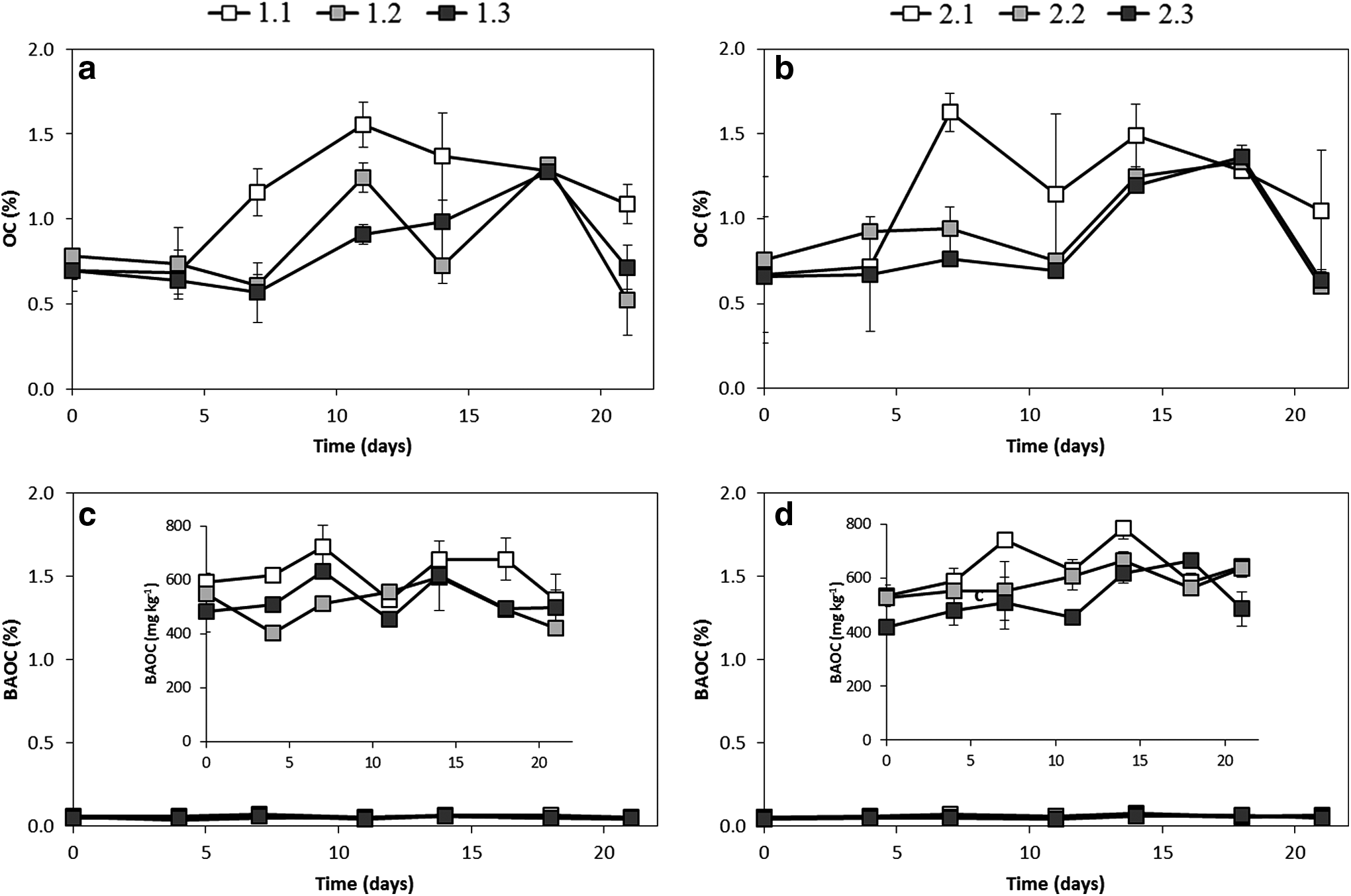
Analysis of total and biologically active organic carbon throughout biofilm growth in channel 1 (fed with river water) (
Regarding the pigments, Chl a increased throughout the experiment up to day 14 in sections 1 and 2, reaching concentrations of 2.97 and 2.38 μg/g, respectively, and then it decreased slightly (Fig. 3a). In the darkest section 3, Chl a concentrations showed little variation or even decreased at intermediate times, not surpassing the initial concentrations of about 1.71 μg/g. Algal cells (together with the EPS) comprise the majority of the biofilm biomass in freshwater biofilms under light conditions (Romaní, 2010). Thus, the increase in Chl a, which is present in all algal groups, can be interpreted as an indicator of biofilm development, demonstrating the positive influence of light availability on the biofilm. The results of this study are also consistent with those obtained by Romaní and Sabater (1999), and Sekar et al. (2002), in stream and lentic freshwaters, respectively, who observed that the concentration of Chl a was significantly higher in light-grown biofilms in comparison with dark-grown biofilms. Chl a concentrations in the experimental channels were in the range of those found by Devesa-Rey et al. (2009) and Sanmartín et al. (2011) for the riverbed sediments of the Anllóns River, which ranged between 3.4 and 83.8 μg/g, and 2.2 and 60.1 μg/g, respectively, but slightly lower than those obtained by Gerbersdorf et al. (2007) in the sediments of the Neckar River (Germany), where Chl a varied between 35 and 197 μg/g.
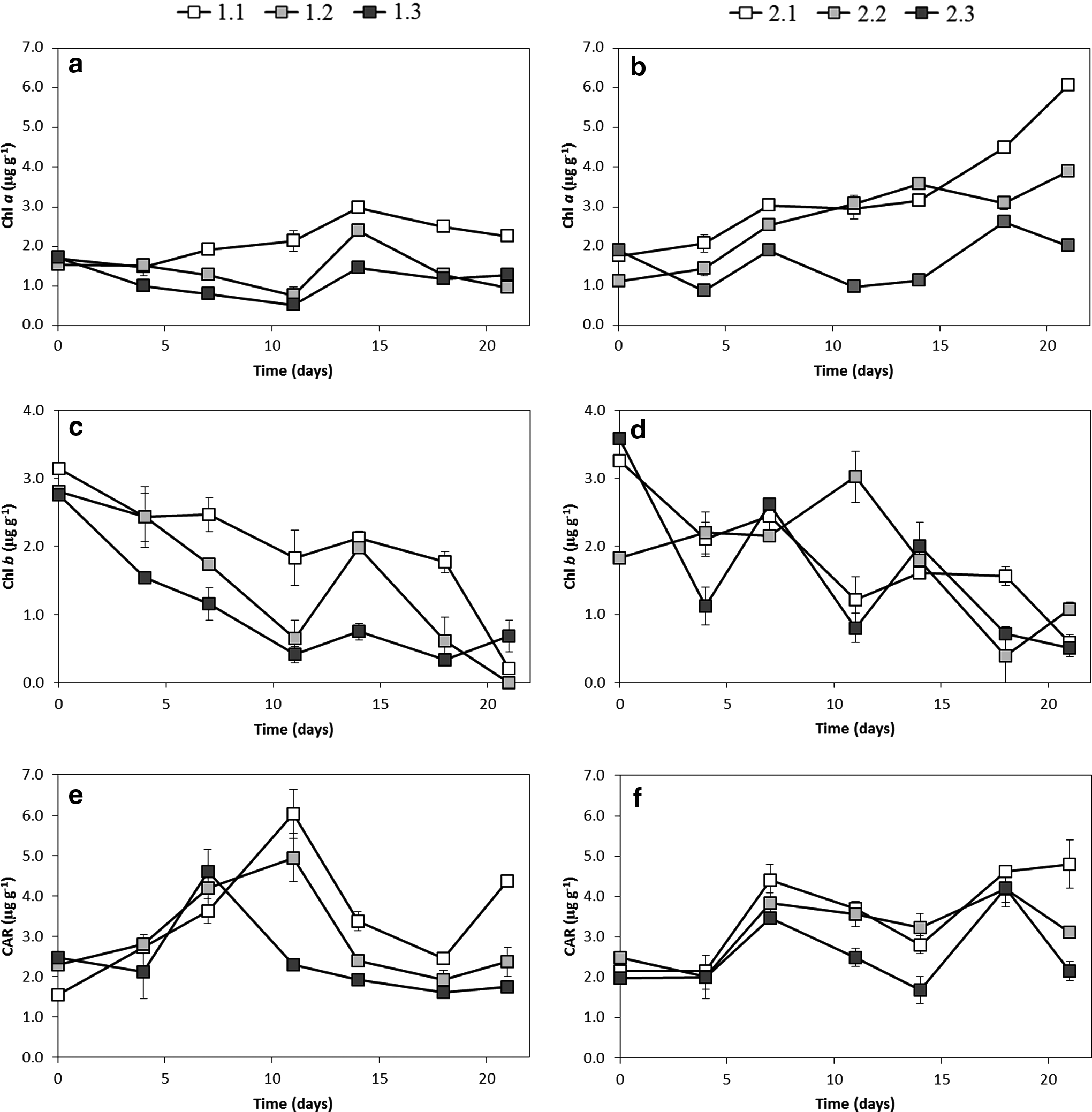
Analysis of phytopigments (chlorophyll a and b and total carotenoids) throughout biofilm growth in channel 1 (fed with river water) (
Chl b concentrations decreased with time in all three sections, the highest values corresponding to the most illuminated section 1, while CAR experienced increase after day 7 (maximum values reached between 7 and 11) with a remarkable decline after reaching the maximun more pronunced for the least illuminated sections (Fig. 3c, e, respectively). The results may suggest a more cushioned decrease of Chl b for the most illuminated sections. Chl b and CAR concentrations are in the same range as those obtained by Devesa-Rey et al. (2009) and by Sanmartín et al. (2011) in the Anllóns riverbed sediments, with Chl b values of 0–31.0 and 2.7–55.6 μg/g, and CAR values of 0–44.5 and 1.0–24.2 μg/g, respectively.
Chl b and CAR act as light-harvesting pigments covering a region of the visible spectrum not accessible to Chl a (Colyer et al., 2005). Among the freshwater microalgae groups, Chl b is present in the Chlorophyta and Euglenophyta divisions, while carotenoids are ubiquitous, each algal group presenting its own particular carotenoid compounds (Leavitt and Hodgson, 2001).
Previous studies carried out in the Anllóns River (Penalta-Rodríguez et al., 2008) showed that diatoms, belonging to the Heterokontophyta division, represent 90–98% of the autotrophic population of the epipsammic biofilm. Diatoms present, as major photosynthetic pigments, chlorophyll a and c, and fucoxanthin (carotenoid) as an accessory pigment (Gómez et al., 2009). The possible predominance of diatoms in the biofilm growth in the experimental channel may explain the increase in Chl a throughout the experiment, without a concomitant increase in Chl b.
As mentioned above, SC are considered a measure of the EPS (Sutherland et al., 1998) and thus they should increase with the development of the biofilm. SC increased up to day 11, 14, and 18, by about 250%, 100%, and 350% of the initial values, for sections 1, 2, and 3, respectively (Fig. 4a), reaching maximum values between 0.32 and 0.50 mg glucose equivalents g−1. SC values are in the range of the data reported by Underwood et al. (1995) for colloidal carbohydrates of four microbial assemblages in freshwater biofilms over sediments, with values between 1.57 and 5.57 mg glucose equivalents g−1. Total C in the form of SC represented between <1% and 33% of the BAOC. The influence of light availability was manifested in a delay of the SC maximum and in a more pronounced decline at the end of the experiment, for the darkest sections.
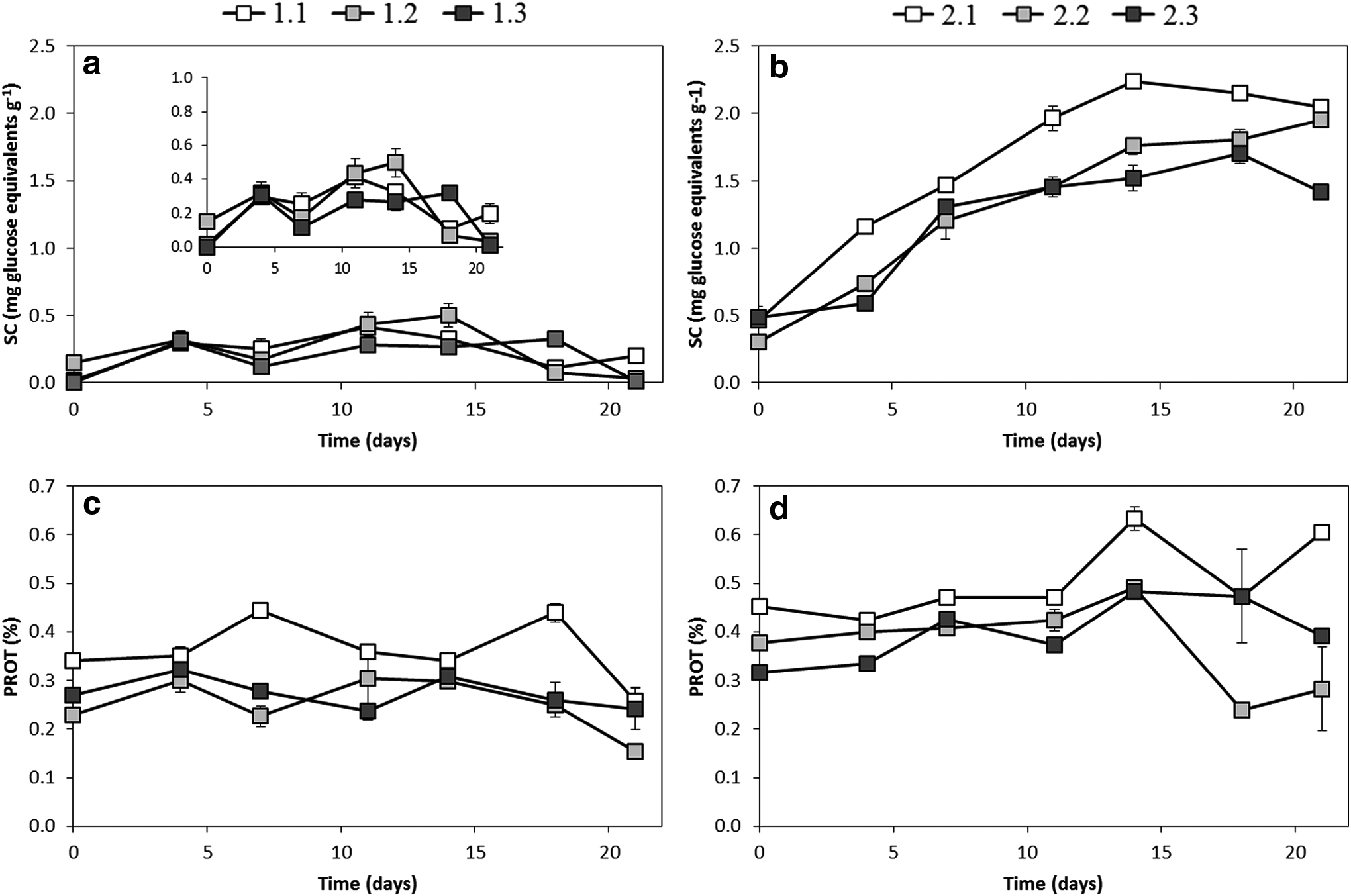
Analysis of soluble carbohydrates and crude proteins throughout biofilm growth in channel 1 (fed with river water) (
Crude protein (PROT) content did not exhibit a clear trend, but points to a decrease after day 18 (Fig. 4c). The influence of light is revealed by the highest PROT concentrations corresponding to the brightest section.
APHOS activity was variable, although the highest values were found for the two most illuminated sections at the end of the experiment (Fig. 5a). It has been observed that the synthesis of extracellular phosphatases by microorganisms is enhanced in a phosphate-depleted medium (Chróst and Overbeck, 1987; Siuda and Chróst, 1987; Romaní et al., 2004; Sabater et al., 2005; Proia et al., 2012). In fact, BioP inversely evolves in relation to APHOS activity (Fig. 5c), decreasing throughout the experiment as a consequence of this nutrient intake by the biofilm.

Analysis of acid phosphatase activity and bioavailable phosphorous throughout biofilm growth in channel 1 (fed with river water) (
In summary, a clearly positive effect of light availability could only be observed for OC, BAOC, Chl a, and CAR. The evolution in time of these parameters showed maximum Chl a and SC concentrations at days 14 and 11–18, respectively. Also, OC, CAR, PROT, and APHOS reached maximum values at intermediate times (days 7–18), whereas Chl b showed a clear decrease over time. These results can be interpreted as light favoring the development of phototrophic organisms in the biofilm, namely Heterokontophyta algae, which have Chl a but not Chl b.
According to Chl a and SC concentrations, an incubation period of 11–18 days is suitable for the development of biofilms, in the experimental conditions of this study. Biofilm development is possible in channel flumes fed with river water, which is of interest for further environmental and biotechnological studies.
Influence of nutrient input on biofilm growth
The second objective of this study was to test whether nutrient inputs modify biofilm growth. The results for OC and BAOC for channel 2, fed with a nutrient-enriched solution, are shown in Figure 2 (b and d, respectively). OC concentration increased over time to reach a maximum value at day 7–14 for the brightest section, and between days 14 and 18 for the other two, and then decreased toward the end of the experiment, most markedly for the darkest section. The highest OC values corresponded to the brightest section 1, but no clear difference attributable to nutrient input was observed in comparison with channel 1. BAOC, which represented between 4.2% and 8.2% of OC, showed similar profiles in both channels, the highest concentration corresponding to the brightest sections.
Chl a profiles showed clear differences between the two channels. Chl a steadily increased over time in the nutrient-enriched channel 2, reaching levels four times higher than in channel 1 (Fig. 3b). The effect of light can be observed in the highest Chl a concentrations found in the brightest sections at days 18 and 21. The importance of nutrients (nitrate and phosphate) as key factors for algal growth was previously reported by Perrin et al. (1987), Borchardt (1996), Tank and Dodds (2003), and Dodds (2006). The combined effects of light and nutrients were put forward by Mosisch et al. (2001) who studied algal production in subtropical streams and revealed that nitrogen stimulated the production of periphyton in streams with sufficient light. Also Taulbee et al. (2005) observed an increase in Chl a concentrations with increasing light levels, for algae growing on substrata to which nitrogen was added. Our maximum values of Chl a were reached in the same period as those previously obtained by Proia et al. (2012), who studied the effect of light and nutrients on biofilms colonizing artificial glass substrata in a Mediterranean stream, and showed that the highest increase of Chl a was reached between days 16 and 22 in all conditions; they also observed a different Chl a temporal pattern under diverse light and nutrient conditions but without significant effects.
With regard to the other two analyzed pigments, Chl b showed similar profile in the two channels, with an almost constant decline, whereas CAR exhibited once again a remarkable increase from day 7 and the maximum values reached at days 18 and 21 (Fig. 3). The effect of light can be observed in the highest CAR concentrations found in the brightest sections at days 7, 18, and 21.
SC were notably higher for channel 2 than for channel 1 (Fig. 4), reaching differences of up to 90%, pointing to a clear influence of nutrient input on this parameter, which is related to EPS and thus to biofilm development. Total carbon in SC represented between 4% and 32% of OC and between 23% and 146% of BAOC. In the brightest section, SC increased up to day 14 and then leveled off. Similar time trends, but with lower SC concentrations, were found for the darkest sections.
PROT analysis revealed a noteworthy effect of nutrient input, as demonstrated by the higher concentrations in channel 2 (Fig. 4). Once again, for channel 2, the brightest section exhibited the highest PROT concentrations, with a maximum value at day 14.
APHOS activity was significantly affected by nutrient availability and was lower in channel 2. Moreover, it tended to decrease toward the end of the experiment, as opposed to the trend observed in channel 1 (Fig. 5). As mentioned above, the synthesis of extracellular phosphatases is enhanced in a phosphate-depleted medium. Therefore, this behavior can be inversely related to sediment BioP concentrations, which were initially higher in channel 2 than in channel 1, and again between days 7 and 14. In the interim, BioP decreased to similar concentrations in both channels, despite the addition of phosphate to channel 2 every 2 days to maintain P concentrations close to 20 mM. This fact suggests that organisms are taking available P from the overlying water and sediment in a very active growth phase. The subsequent increment in BioP may be explained by the repeated P input and the lower uptake by the organisms at later stages of growth.
General overview
This study attempts to assess the influence of light and nutrient inputs on biofilm development in mesocosm conditions. Nutrients may change in river waters in response to soil leaching and runoff and to inputs of cattle slurries and manure and wastewater to the river course. Light availability is mainly affected by the riparian cover and the turbidity. The results herein highlight the influence of nutrient input on biofilm growth, mostly under the nonlight limiting conditions in which the highest algal growth occurred. Light availability also had direct effects on parameters indicative of biofilm development. Chl a, which can be considered a proxy of photosynthetic microorganisms, appears to be the most informative parameter related to biofilm growth under different light availability and trophic state. SC, which can be considered a proxy of the EPS embedding cells in the biofilm, is also a sensitive parameter that widely responds to the combination of brightness and high nutrient conditions.
Overall, the results confirmed that (a) light intensity and trophic state affects biofilm growth in experimental channels. Light is the key factor as limiting light reduces biofilm growth even in the presence of nutritive media; (b) the designed experimental fluvial channels can be employed to obtain epipsammic biofilm for use in environmental and biotechnological experiments; and (c) epipsammic biofilm can grow in channels fed only with river water, thus avoiding the affectation of competitor aninons or high ionic strength due to the presence of supplementary nutrients.
Conclusions
This study demonstrates that biofilm growth is affected by the different conditions of light intensity and water composition set for the experimental fluvial channels. Increasing light availability favors the growth of the autotrophic component of the biofilm, and this effect is more noticeable under the highest nutrient conditions. The most sensitive parameters are chlorophyll a, total carotenoids, SC, and crude proteins, whereas phosphatase activity is conditioned by bioavailable P concentrations. Experimental indoor fluvial channels can be successfully used for the development of biofilm over riverbed sediments, which could be used in further environmental research and biotechnological applications.
Footnotes
Acknowledgments
The authors wish to thank the Spanish Ministry of Economy and Competitiveness for (MINECO-FEDER) for financial support (Projects Ref. CGL2010-22059 and CGL2013-46003P). Diego Martiñá Prieto wishes to acknowledge the support of the Spanish Ministry of Economy and Competitiveness for his personal funding (FPI Fellowship Ref. BES-2011–044514). David Rubinos is also grateful for personal funding of the Xunta de Galicia (Plan Galego de Investigación, Innovación e Crecemento—I2C, Consellería de Educación e Ordenación Universitaria) and the European Social Fund. Additionally, the authors also wish to thank lab technician Montserrat Recarey Pérez for her support.
Author Disclosure Statement
No competing financial interests exist.