
Abstract
Abstract
Anaerobic digestion (AD) is the most feasible technology to harness the energy of dairy manure as biogas. However, long retention time (21 days or longer) is needed because of recalcitrance of dairy manure fiber. In this study, we evaluated a thermophilic one-stage AD (T-1stg-AD), a thermophilic-mesophilic two-stage AD (referred to as temperature-phased AD, TPAD), and a mesophilic two-stage AD (M-2stg-AD) system, all at a 15-day retention time, for performance in digesting dairy manure and for variations of microbial populations. The T-1stg-AD system achieved the best performance, followed by the TPAD and the M-2stg-AD systems. For the TPAD system, its first-stage thermophilic digester outperformed its second-stage mesophilic digester, whereas the two digesters of the M-2stg-AD system had similar performance. Each digester harbored distinctive microbial populations, several of which were significantly correlated with system performance. Microbiomes of the two digesters of the TPAD system differed greater than the microbiomes of the two digesters of the M-2stg-AD system. Methanosarcina and Methanobacterium were the most predominant methanogen genera in the first-stage digesters of TPAD and M-2stg-AD and in the T-1stg-AD system, whereas Methanosaeta was the most predominant genus in the second-stage digesters of TPAD and M-2stg-AD. Population size of these genera appeared to be adversely correlated to VFA concentrations. The T-1stg-AD and the TPAD system can be better suited to digest dairy manure, with an additional benefit to kill off pathogens. Results may help further understand the microbiological underpinning of these digester systems and future implementation of AD systems on dairy farms.
Introduction
A
Most of the anaerobic digesters currently in full-scale operation are mesophilic one-stage digesters. Because they are operated at mesophilic temperature and in single stage, such digesters can suffer from poor system performance (Angelidaki et al., 2003) and susceptibility to high organic loading rates (OLR) (Speece et al., 2006). Thus, different AD processes have been developed to improve process stability and to increase biogas yield (Mata-Alvarez et al., 2000; Ye and Chang, 2008; Lv et al., 2010). Thermophilic one-stage AD (T-1stg-AD) systems, mesophilic two-stage AD (M-2stg-AD) systems, and temperature-phased AD (TPAD) systems are considered some of the most promising alternatives. Compared with mesophilic one-stage digesters, T-1stg-AD systems improve solid removal and pathogen inactivation (Song et al., 2004; Yilmaz et al., 2008), whereas M-2stg-AD systems improve performances at elevated OLR (Demirer and Chen, 2005), and TPAD systems improve pathogen inactivation, solid removal, and methane production (Santha et al., 2006; Demirer and Othman, 2008; Riau et al., 2010). Previous studies also suggested that TPAD systems could achieve superior performances over both T-1stg-AD systems (Kaiser et al., 1995; Riau et al., 2010) and M-2stg-AD systems (Dugba and Zhang, 1999; Kim et al., 2004). In recent studies, we showed that TPAD is a potential technology that can efficiently convert dairy manure to biogas with a shorter (10–15 days) retention time than that typically used for CSTR or MPFLR digesters (Lv et al., 2013a, 2013b; 2016). However, the aforementioned types of digester systems have not been systematically evaluated when fed dairy manure and operated with a similar retention time. In the present study, we comparatively evaluated TPAD against T-1stg-AD and M-2stg-AD in digesting dairy manure with respect to digester performance. The microbiome composition in these types of digesters and the association between individual populations and digester performance, both of which remain poorly understood (Hofman-Bang et al., 2003; O'Flaherty et al., 2006), were also investigated.
Materials and Methods
Digester setup
Digesters used in the present study were the same as those used in previous studies (Lv et al., 2013b, 2016). Briefly, the bench-scale digesters were made by using Nalgene polypropylene wide-mouth bottles of 4.3-L capacity (Fisher Scientific). Each digester had a feeding port, a sampling port, and a gas outlet. The feeding port and the sampling port were sealed by rubber stoppers to keep the digesters airtight except during feeding and sampling. Biogas was collected and recorded daily by water displacement by using an inverted graduated cylinder filled with tap water and placed in a water bath maintained at room temperature. All digesters were placed in water baths to maintain their respective temperatures. The operational parameters of each AD system were summarized in Table 1.
TPAD, temperature-phased anaerobic digestion; M-2stg-AD, mesophilic two-stage anaerobic digestion; T-1stg-AD, thermophilic single-stage anaerobic digestion; N/A, not applicable.
Seed sludge
A TPAD system, with the thermophilic digester operated at 50°C and the mesophilic digester operated at 35°C, had been run consecutively for 144 days digesting dairy cattle manure (Lv et al., 2016). The contents from these two digesters were used as seed sludge to inoculate the digesters of the TPAD system in this study. The thermophilic and the mesophilic seed sludge contained 11.5% and 9.3% total solid (TS), and 9.7% and 7.4% volatile solid (VS), respectively. The first-stage thermophilic digester of the TPAD system used in the present study was filled to its working volume (1 L) with a mixture of the seed sludge from the thermophilic and the mesophilic digesters (1:1 ratio), whereas the second-stage mesophilic digester was inoculated with the mesophilic seed sludge (1.5 L) only. During the startup, the content of the mesophilic digester was not discharged until it reached the working volume (2 L).
After the operation of the TPAD system was completed, the content (1 L) of the first-stage thermophilic digester was used as the seed sludge for the T-1stg-AD, whereas the content (2 L) of the second-stage mesophilic digester was used to inoculate the M-2stg-AD systems, with each stage receiving 1-L inoculum. The content of the T-1stg-AD digester and the second-stage digester of the M-2stg-AD was not discharged until the working volume (1.5 L for the former and 2 L for the later) was reached. The thermophilic and the mesophilic seed sludge contained 10.1% and 8.9% TS, and 8.6% and 7.3% VS, respectively.
Feedstock
Dairy cattle manure (composed of feces and urine) was collected on a daily basis from the Waterman Dairy Center, The Ohio State University. Based on dry matter, the total mixed ration fed to the cattle was primarily composed of corn silage (50%), alfalfa hay (4.5%), co-product of corn wet milling (21%), ground corn (9.05%), soybean meal (4.64%), Aminoplus® (1.3%), soy hulls (1.3%), fat (0.38%), and vitamin and minerals (2.01%). The TS and the VS content of the collected manure averaged 14.61% (w/v) and 12.81% (w/v), respectively. Before use, manure was diluted to the desired TS and VS content by using tap water and was mixed thoroughly into slurry. This was done to mimic the manure slurry fed to large-scale manure digesters, to reduce potential clogging in digesters (Sung and Santha, 2003), and to improve the substrate accessibility to the microbial community (Angelidaki and Ahring, 2000).
Startup, operation, and sampling
Each AD system was fed with a manure slurry of 13.6% TS and 11.9% VS and operated in a fed-batch mode on a daily basis. The two-staged systems (TPAD and M-2stg-AD) were operated essentially as previously described (Lv et al., 2013b). Briefly, the effluent from the second-stage digester was discharged; then, the same volume of the content from the first-stage digester was transferred to the second-stage digester; and finally, the same volume of manure slurry was fed to the first-stage digester. In the T-1stg-AD system, the effluent was discharged from the digester before the same volume of manure slurry was fed into it. The content of each digester was manually mixed both before and after feeding. Biogas (equilibrated to room temperature and one atmospheric pressure) production and effluent pH from each digester were recorded daily before feeding. During the startup of each two-stage system, the effluent from the second-state digester was recycled back to the first-stage digester once daily to expedite the startup until the first-stage digester reached stable pH (Lv et al., 2016).
As suggested by Sung and Santha (2003), the TPAD system was operated at an overall hydraulic retention time (HRT, also solid retention time, SRT) of 15 days. For comparison, all the three AD systems had an HRT/SRT of 15 days. Five days was allowed for each of the first-stage digesters of both the TPAD and the M-2stg-AD systems. The three AD systems were operated until they reached their steady states, when the variation of daily biogas production from each digester was less than 10% for 5 consecutive days without any upward or downward trend (Wen et al., 2007). Multiple biogas and sludge samples from each digester were collected during its steady state. The sludge samples were aliquoted and stored at −80°C until they were analyzed.
System performance analysis
Contents of CH4 and CO2 in biogas samples were analyzed by using gas chromatography (GC) as previously described (Patra and Yu, 2014), and methane yields were calculated from corresponding biogas yield and methane content. Sludge samples were centrifuged to collect their supernatants, and the VFA concentrations were analyzed by using GC (Lv et al., 2013a). Content of TS and VS of each sludge sample was determined by following the standard methods (American Public Health Association et al., 2005), and solid removal was calculated for each digester.
Microbial community analysis
Metagenomic DNA was extracted from each sludge sample by using the repeated bead-beating plus column purification (RBB+C) method (Yu and Morrison, 2004b). The DNA quality was evaluated by using agarose gel (0.8%) electrophoresis. Each DNA sample was quantified by using a NanoDrop spectrophotometer (Thermo Scientific) and then diluted to a final concentration of about 50 ng/μL with Tris-EDTA buffer.
Archaeal and bacterial communities in each DNA sample were profiled by using PCR-denaturing gradient gel electrophoresis (DGGE) as previously described (Yu and Morrison, 2004a; Yu et al., 2008). Briefly, the V3 hypervariable region of the 16S rRNA genes was amplified from the DNA samples by using domain-specific primers for both archaea and bacteria (Table 2), and the amplicons were resolved on acrylamide (8%) gel containing a linear 40–60% denaturing gradient. DGGE gel images were analyzed by using BioNumerics version 5.1 (Applied Maths, Inc.) to determine the band profiles of each DNA sample as previously described (Cressman et al., 2010). Dendrograms based on the similarity matrix of DGGE profiles were constructed by using the Jaccard similarity coefficient and the UPGMA (Unweighted Pair Group Method with Arithmetic Mean) clustering method. The determined DGGE profiles were also subjected to principal component analysis (PCA) as previously described (Cressman et al., 2010).
Both aGC-Arc344F and GC-Eub357F had a 40nt GC-clamp attached at their 5′ ends.
DGGE, denaturing gradient gel electrophoresis.
Four methanogen genera, including Methanobacterium, Methanoculleus, Methanosaeta, and Methanosarcina, the WSA2/ArcI group, and total archaea in each DNA sample were quantified by using respective primer/TaqMan probe sets (Table 2). Briefly, a sample-derived qPCR standard was prepared for each target genus or group from a pooled DNA sample representing all the DNA samples to be analyzed as previously described (Yu et al., 2005). Each of the qPCR standards was subsequently quantified and serially diluted with Tris-EDTA buffer to achieve an eight-log range of concentrations (i.e., 100–108 amplicon copies/μL). Each targeted group was quantified in three technical replicates against its corresponding qPCR standard on the same qPCR plate. After an initial denaturation at 95°C for 5 min, all qPCR assays were carried out in 40 PCR cycles of 95°C for 15 s and 60°C for 1 min, with fluorescent signals being acquired at the end of the 60°C step. Abundance was calculated as copies of 16S rRNA genes/ml of sludge sample.
Statistical analysis
T-test was used to make pairwise comparisons among means of system performances (i.e., biogas/methane yield and VS removal), VFA concentrations, and abundances of methanogen genera/total archaea. In all three cases, p ≤ 0.05 was declared statistically significant. Standard deviations (SD) were also calculated for all the mean data. Pearson correlation coefficients were calculated to assess the correlation between population abundances of methanogen genera/total archaea and methane productions/VFA concentrations. Significant correlation was claimed at p ≤ 0.05.
Results and Discussion
System performance
A previous study showed that 50°C in the thermophilic digester produced as much biogas as 55°C and 60°C (Lv et al., 2013a). Thus, the first-stage digesters of the TPAD system and the T-1stg-AD were maintained at 50°C in this study. Without external adjustment for pH, all the digesters maintained a stable pH of about 7.2 during stable operation, except for the first-stage digester of the M-2stg-AD system that had a pH of about 6.5. The stable pH in each digester indicated that a dynamic balance was achieved between hydrolysis/acidogenesis and methanogenesis. The VFA concentration in the first-stage digester of the M-2stg-AD system was significantly (p < 0.05) higher than that in all the other digesters (Fig. 1), and the high VFA concentration in this digester was adversely correlated to the low pH therein. The low pH and the high VFA concentration corresponded to low methane production from the first-stage digester of the M-2stg-AD system and suggested that the first-stage digester of the M-2stg-AD system might be susceptible to elevated OLR. It is not known why the concentrations of VFA, particularly acetic and propionic acids, were lower in the first-stage digester of the TPAD system than in the counterpart of the M-2stg-AD system. Because more biogas was produced in the former than in the latter (Table 3), one plausible explanation is the higher rate of methane production through the hydrogenotrophic methanogenesis pathway coupled with syntrophic acetate oxidation, which is favored in thermophilic digesters (Ho et al., 2013, 2014). This explanation is consistent with a shift from acetoclastic methanogenesis to hydrogenotrophic methanogenesis when dilution rate was high (corresponding to short HRT/SRT) (Shigematsu et al., 2004). Future research to determine the population of syntrophic acetate-oxidizing bacteria and their activity can help confirm the earlier premise.
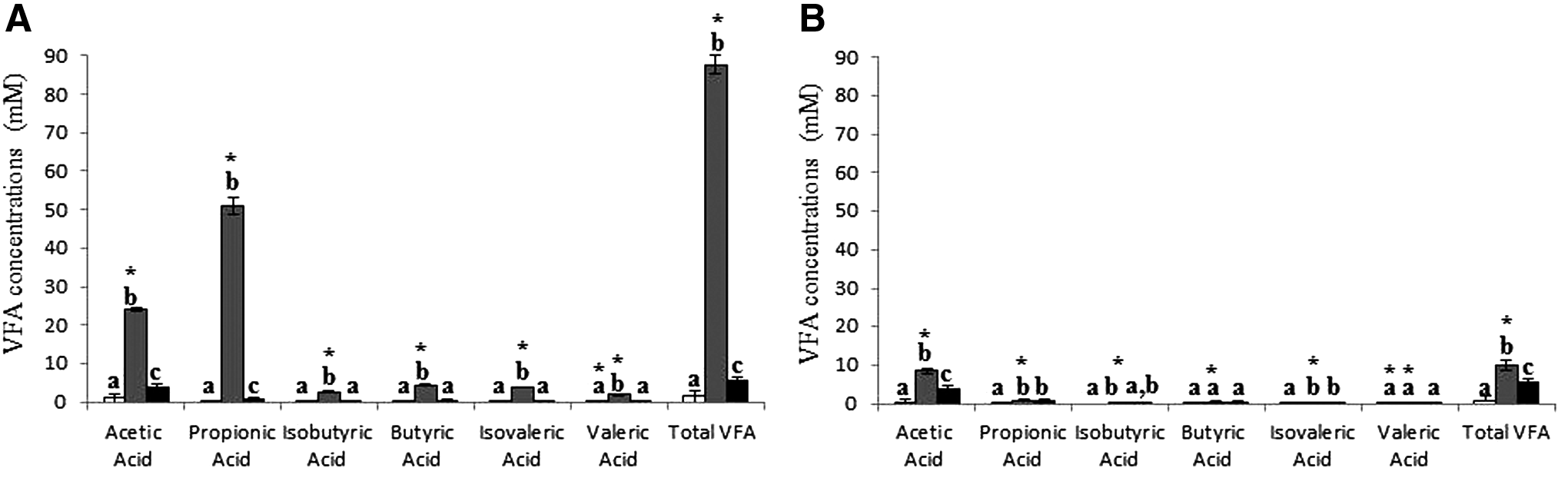
VFA concentrations in the first-
L/L working volume/day. Data were presented as the means ± SD (TPAD, n = 6; M-2stg-AD, n = 3; T-1stg-AD, n = 3). Values in each column with different superscripts were significantly different (p < 0.05).
The first-stage digester of the TPAD system achieved significantly (p < 0.05) greater VS removal and methane production than the first-stage digester of the M-2stg-AD system, whereas the opposite was true for the second-stage digesters of these two two-stage systems (Tables 3 and 4). This better performance of the first-stage digesters of the TPAD system than of the M-2stg-AD system is expected, and it might be attributable to differences in microbiome and metabolic activities that resulted from the different temperatures. The second-stage digesters of these two two-stage systems also had different microbial communities that might have led to different performances. This is conceivable given that the influxes from the first-stage digesters contained significantly (p < 0.05) different VFA concentrations and microbial populations between these two two-stage systems (Fig. 1). The M-2stg-AD system had the lowest overall VS removal and methane production among the three AD systems (Tables 3 and 4). The TPAD and the T-1stg-AD systems had similar (p > 0.05) overall VS removals and biogas production, but the latter produced significantly (p < 0.05) more methane than the former. The T-1stg-AD system achieved the highest system performance among the three AD systems. The elevated temperature might have enhanced balanced hydrolysis/acidogenesis and methanogenesis (as reflected by the stable nearly neutral pH) both in the thermophilic digester of the TPAD system and in the T-1stg-AD system, which led to the superior performances of these two digesters. However, the T-1stg-AD system also needed the highest energy input for heating because its entire content was maintained at 50°C, whereas only the content of the thermophilic digester of TPAD, one third of the entire TPAD content, was maintained at 50°C. It should be pointed out that antimicrobial use in the production of food-producing animals, including dairy cattle, will be restricted starting in 2016 in the United States to combat widespread antimicrobial resistance. Because thermophilic digestion of animal manure can decrease antibiotic resistance by up to three logs (Diehl and LaPara, 2010; Ma et al., 2011) and pathogen load by six logs (Smith et al., 2005; Rounsefell et al., 2013), the TPAD- and T-1stg-AD systems have an additional advantage by decreasing antibiotic resistance and pathogen loads present in dairy manure. This is particularly important when the solid in the digested dairy manure is recovered and used as bedding materials. These results may help dairy farmers to determine the type of digester system to implement on their farms.
g VS/L working volume/day. Data were presented as the means ± SD (TPAD, n = 6; M-2stg-AD, n = 3; T-1stg-AD, n = 3). Values in each column with different superscripts were significantly different (p < 0.05).
VS, volatile solid.
In the TPAD system, the first-stage digester accounted for 78.0% of total VS removal and 75.4% of total methane production, whereas the second-stage digester only accounted for 22.0% VS removal and 24.6% methane production. In the M-2stg-AD system, the first-stage digester accounted for 45.4% of total VS removal and 41.7% of total methane production, whereas the second-stage digester accounted for 54.6% VS removal and 58.3% methane production. This suggests that the two digesters of the TPAD system had different roles, with the first-stage digester making the primary contribution to the overall system performance, and the second-stage digester only scavenging residual hydrolysis and acidogenesis products received from the first-stage digester. In contrast, the two digesters of the M-2stg-AD system made comparable contributions to the overall system performance. These results also indicate that the operation temperature of the first-stage digester not only affected the performance thereof but also had extended impact on the performance of the downstream second-stage digester, possibly by changing metabolites and microbial populations fluxed from the first-stage digester to the second-stage digester.
Microbiome profiles and methanogen populations
Clustering analysis of the DGGE profiles showed that both the archaeal and the bacterial communities present in both the first- and the second-stage digesters of the two-staged systems generally differed between the first- and the second-stage digesters in each two-stage system and between the corresponding digesters (Fig. 2). The T-1stg-AD system developed unique archaeal and bacterial communities, which differed from those of the two-stage systems. The PCA analysis results were consistent with those of the clustering analysis of DGGE profiles (Fig. 3). The difference in both bacterial and archaeal communities between the two digesters of the M-2stg-AD system was smaller than that between the two digesters of the TPAD system (Fig. 3E, F). As expected, the T-1stg-AD system had different archaeal and bacterial communities than the two-stage systems. As reflected by the digester performance, the two digesters of the TPAD system had distinctive microbial communities that played different roles, whereas the two digesters of the M-2stg-AD system had relatively similar microbial communities and might as well function similarly. These results again indicate that the operation temperature of the first-stage digester can have both local and extended impact on the microbial communities in a two-stage AD system. The relatively similar microbial communities between the two digesters of the M-2stg-AD system might be attributable to the recalcitrant nature of the substrates (dairy manure fiber) present in the fed dairy manure so that the two stages had relatively similar feedstock. The same operation temperature of the two stages of the M-2stg-AD system is another factor that is attributable to the relatively similar bacterial and archaeal communities therein. These results were consistent with the findings of recent studies (Kundu et al., 2012, 2013) in that both different operation temperatures and HRT/SRT can lead to development of different microbial communities in an AD system.
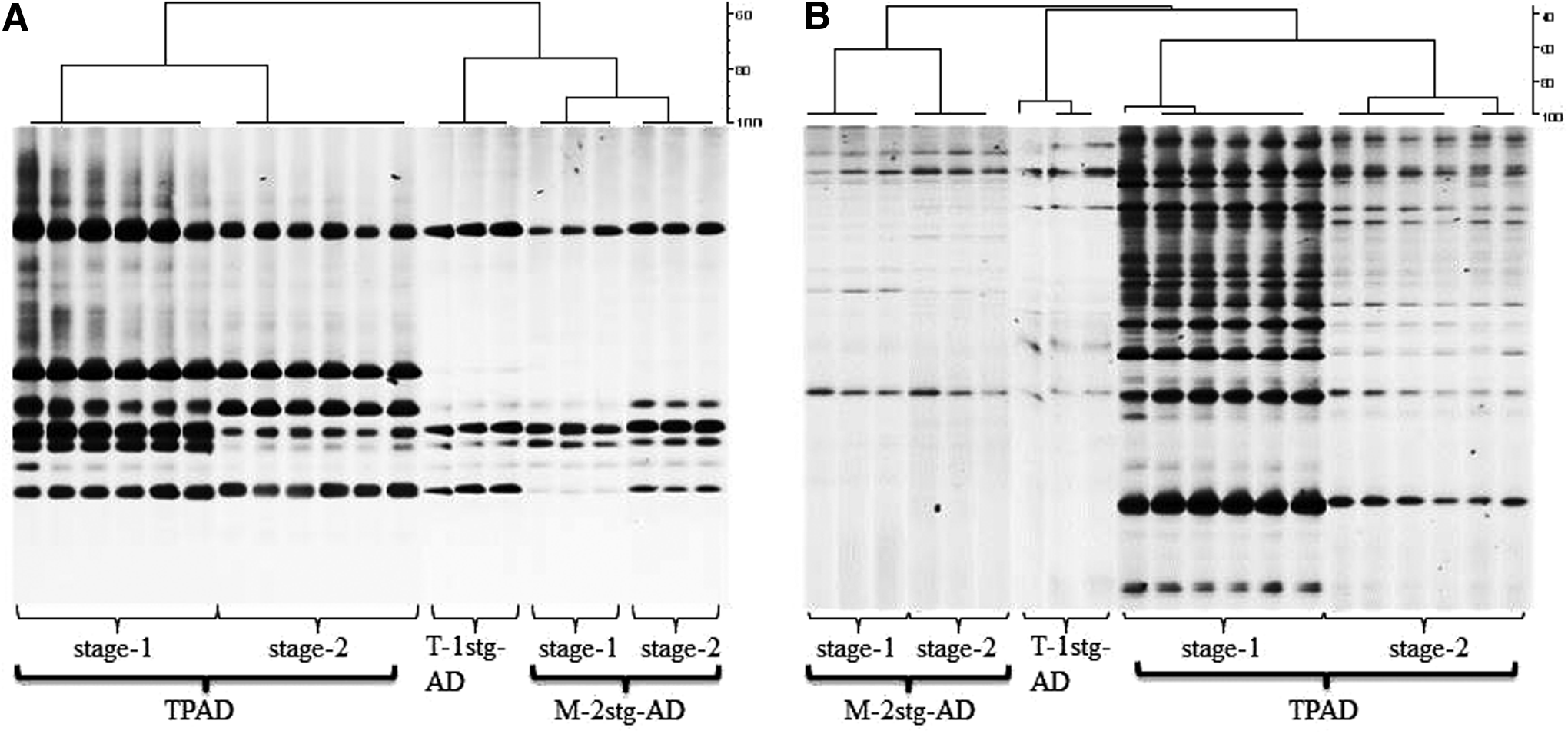
Archaeal
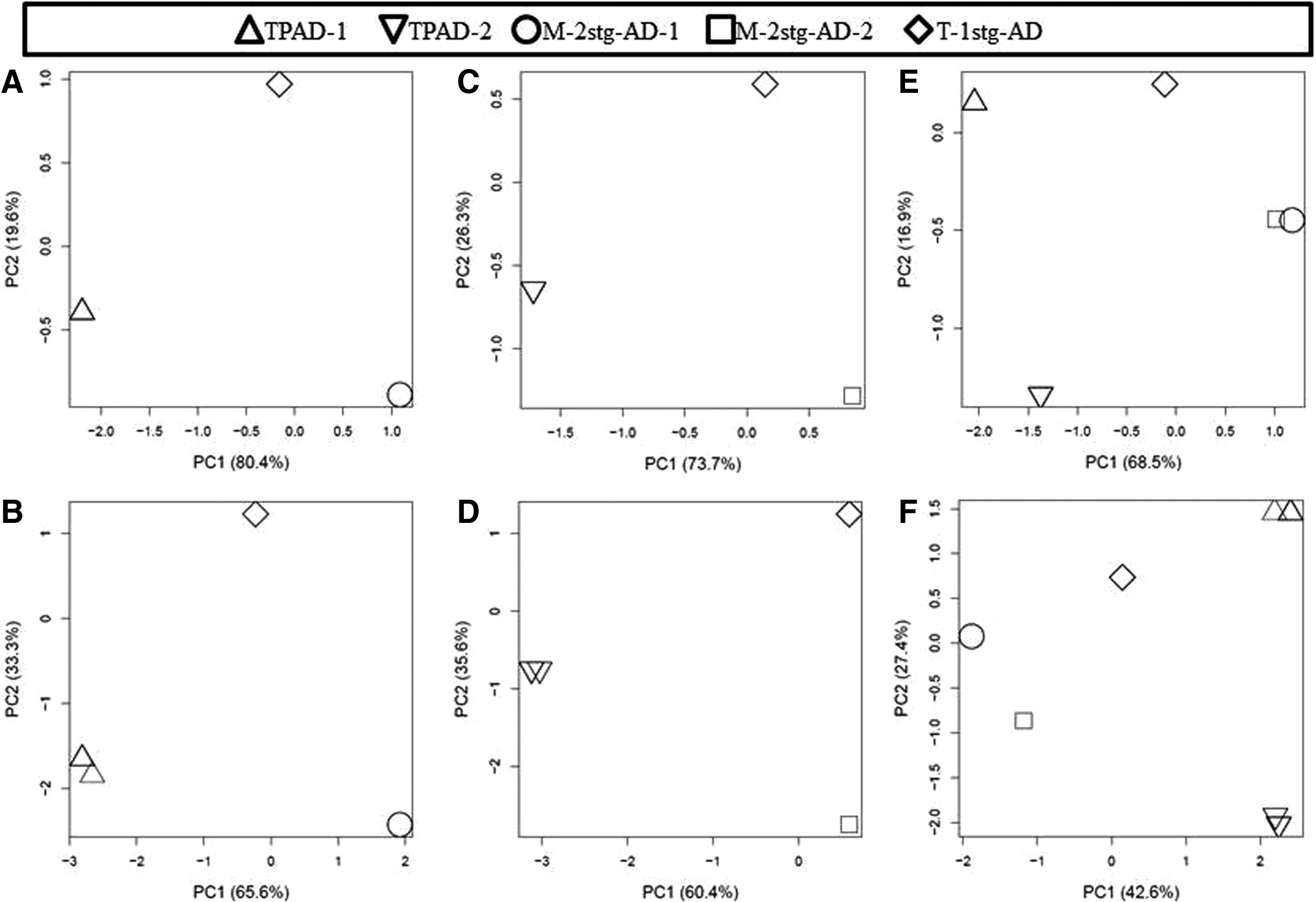
PCA plots of archaeal
In the first-stage digesters of both the TPAD and the M-2stg-AD systems, Methanosarcina and Methanobacterium were the two most predominant methanogen genera (Fig. 4A), which suggests that these two genera were primarily responsible for the methane production in these two digesters. The first-stage digester of the TPAD system had a larger population of all the three detected methanogen genera (Methanobacterium, Methanoculleus, and Methanosarcina) than the first-stage digester of the M-2stg-AD system. Because the latter had a higher VFA concentration than the former, the populations of these methanogens are adversely correlated to the concentrations of VFA (primarily propionic acid). The other acetoclastic genus, Methanosaeta, was not detected in the first-stage digesters of the TPAD or the M-2stg-AD systems. Most members of the genus Methanosaeta grow slowly and have a long generation time of up to 9 days (Yu et al., 2010). Apparently, the short HRT/SRT (i.e., 5 days) of these two digesters did not provide a long enough time for Methanosaeta to be established therein. This genus of acetoclastic methanogens was also not detected in the T-1stg-AD system, which had an HRT/SRT of 15 days. The lack of detection of Methanosaeta in this system is consistent with its dearth in thermophilic digesters reported in other studies (Demirel and Scherer, 2008; Lv et al., 2016). The harsh conditions created by the thermophilic temperature might be responsible for the lack of this genus of fastidious methanogens. The second-stage digester of the M-2stg-AD system contained a large population of Methanosaeta, reflecting the permissive conditions created by its low VFA concentration and long HRT/SRT.
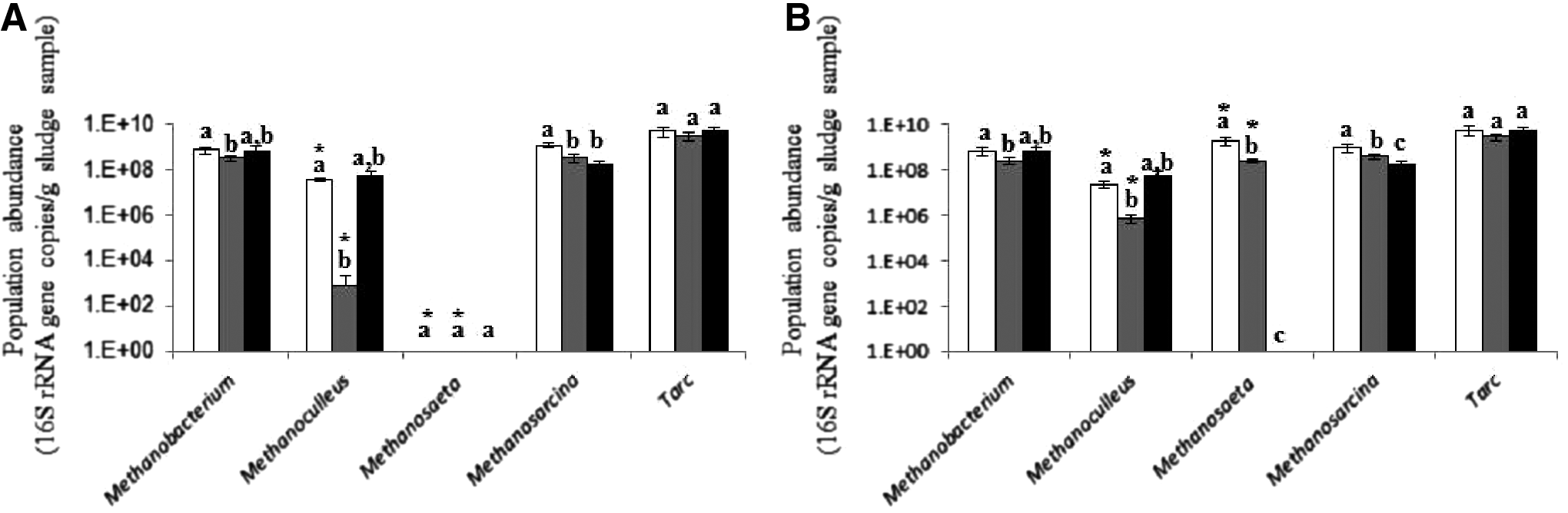
Population abundance of four methanogen genera and total archaea (Tarc) in the first-
In the second-stage digester of the TPAD system, the most predominant methanogen genus was Methanosaeta, with its population significantly (p < 0.05) larger than that of Methanosarcina (Fig. 4B). This suggests that methane might have been primarily produced in this digester by Methanosaeta through the acetoclastic methanogenesis pathway. Methanosaeta spp. generally have lower Ks values and higher affinities for substrates (i.e., acetate) than Methanosarcina spp. (Yu et al., 2010), which make Methanosaeta more competitive than Methanosarcina in the second-stage digester of the TPAD system where the influx from the first-stage digester contained low concentrations of VFA (primarily acetic acid, Fig. 1). In the second-stage digester of the M-2stg-AD system, Methanosarcina, Methanobacterium, and Methanosaeta had similar population sizes, indicating that both the acetoclastic and the hydrogenotrophic methanogenesis pathways probably contributed to the methane production in this digester. Methanosarcina spp. generally have higher growth rates and tolerances against high VFA concentrations than Methanosaeta spp. (Conklin et al., 2006; Yu et al., 2010). Considering the high concentrations of VFA (primarily propionic acid) in the influx from the first-stage digester and the hydrogen production during the syntrophic acetogenesis, Methanosarcina and Methanobacterium might be as important as Methanosaeta in the second-stage digester of the M-2stg-AD system.
In the T-1stg-AD system, Methanosarcina and Methanobacterium were the two most predominant methanogen genera (Fig. 4), which agrees with the predominance of these two genera in the first-stage digesters of the two two-stage systems. The predominance of these two genera suggests their importance in methane production from the digesters of different AD systems where intense hydrolysis/acidogenesis and syntrophic acetogenesis/methanogenesis can occur. Methanobacterium was the most abundant hydrogenotrophic methanogenic genus in all the thermophilic digesters, which is consistent with the results of a previous study (Krakat et al., 2010). On the other hand, Methanosaeta only established itself in the mesophilic digesters where sufficient retention time was allowed and VFA concentrations were low (Guo et al., 2015). The WSA2/ArcI group, first discovered in several AD systems treating municipal sewage and proposed to have a hydrogenotrophic metabolism (Chouari et al., 2005; Riviere et al., 2009), was not detected from the pooled community DNA sample that was used to prepare the sample-derived qPCR standards, so this group of methanogens was not analyzed. It is possible that the WSA2/ArcI group is not essential for methanogenesis from dairy cattle manure.
Correlation between system performance and methanogen populations
In the first-stage digesters of the two two-stage systems and in the T-1stg-AD system, the abundance of Methanosarcina and the methane production were significantly (p < 0.05) positively correlated. The abundance of Methanosarcina was significantly (p < 0.05) greater in the first-stage digester of the TPAD system than in the other digesters (Fig. 4), which mirrors the significantly (p < 0.05) higher methane production from the first-stage digester of the TPAD system than from the other digesters. As one of the most predominant methanogen genera in all the digesters, the population of Methanosarcina, which has high growth rate and high Ks (Yu et al., 2010), probably made important contributions to methane conversions in all the three AD systems, especially when intense hydrolysis/acidogenesis and syntrophic acetogenesis/methanogenesis occurred in the thermophilic digester of the TPAD system.
Populations of both Methanosarcina and Methanosaeta were significantly (p < 0.05) greater in the second-stage digester of the TPAD system than in the counterpart of the M-2stg-AD and in the T-1stg-AD, whereas the methane production from the second-stage digester of the TPAD system was the lowest among these three digesters. Given the fact that VFA concentrations in the first-stage digesters were significantly different between the two two-stage systems (Fig. 1), and Methanosaeta was not detected in any of the first-stage digesters or in the T-1stg-AD system, it was hypothesized that the population of Methanosaeta established better in the mesophilic digesters with a lower concentration of VFA. The highest abundance of Methanosarcina in the second-stage digester of the TPAD system might be attributed to carryover, at least partially, from the first-stage digester of this system where the population abundance of Methanosarcina was significantly (p < 0.05) higher than that of the first-stage digester of the M-2stg-AD system and of the T-1stg-AD system.
Except for Methanosarcina, the abundance of the other three methanogen genera was significantly (p < 0.05) negatively correlated to the total VFA concentration. Population abundance of these groups was significantly (p < 0.05) higher in the digesters with significantly (p < 0.05) lower concentrations of total VFA (primarily propionic acid). This confirmed that high VFA concentrations were adversely correlated to population abundances of methanogens (Chen et al., 2008). Methanosarcina might be resistant, to some extent, against the inhibition of propionic acid.
Conclusion
The T-1stg-AD system achieved the greatest methane yield and VS removal, followed by the TPAD and the M-2stg-AD systems. Balanced hydrolysis/acidogenesis and methanogenesis can be achieved at thermophilic temperature when digesting dairy manure, leading to better system performances. Both performance and microbial communities had larger differences between the two digesters in the TPAD system than between the two digesters in the M-2stg-AD system. Each digester harbored distinctive microbial populations. Methanosarcina and Methanobacterium were the most dominant methanogenic genera in the first-stage digesters and the T-1stg-AD system, whereas Methanosaeta was the most dominant methanogen genus in the second-stage digesters. Their populations were adversely correlated to high VFA concentrations. Either TPAD or T-1stg-AD may be better suited for converting dairy manure to biogas. In addition to enhance digestion, these two systems can help decrease antibiotic resistance and pathogen loads present in dairy manure.
Footnotes
Acknowledgments
This work was partially supported by a Department of Energy grant (award number: DE-FG36-05GO85010) and a North East SUN grant (award number: 52110–8512).
Author Disclosure Statement
No competing financial interests exist.