
Abstract
Abstract
Two laboratory-scale partial nitritation/anammox (PN/A) bioreactors—suspended growth (SR) and attached growth (AR)—were initiated to investigate microbial population differences under an elevated temperature (35°C) and room temperature (21°C). During more than 300 days of operation, SR was able to remove 0.54 ± 0.093 kg-N/(m3·d) at 35°C and 0.57 ± 0.061 kg-N/(m3·d) at 21°C. Similarly, AR could remove 0.60 ± 0.111 and 0.56 ± 0.103 kg-N/(m3·d1) at 35°C and 21°C, respectively. Similarity in nitrogen removal rates was probably due to presence of similar microbial communities predominated by three phyla: Chloroflexi, Planctomycetes, and Proteobacteria, nearly same numbers of key functional genes and similar nitrogen loadings. Although seeded from different biomass sources, both the SR and AR operated as sequential batch reactors harbored the same anammox bacteria of Brocadia fulgida and ammonium oxidizing bacteria of Nitrosomonas sp., which suggested the existence of competitive advantages over other species. Moreover, this study indicated that increasing temperature during startup could accelerate the startup process for PN/A systems. Ammonia monooxygenase (amoA) functional gene copy numbers were 1.25 ± 0.08 × 109 and 1.85 ± 0.16 × 109 in SR and AR, respectively, at 35°C. The amoA gene copy numbers were 0.78 ± 0.11 × 109 and 1.9 ± 0.09 × 109 in SR and AR, respectively, at 21°C indicating a negligible change in amoA gene copy numbers with temperature decrease. Similar results were obtained with hydrazine synthase (hzsA). Although, this article presents results in the context of sidestream application of PN/A systems, results are equally important for mainstream PN/A applications.
Introduction
A
The PN/A process is mainly driven by two groups of prokaryotes: (1) ammonium oxidizing bacteria (AOB) and (2) anammox bacteria (ANAMMOX). First, AOB oxidizes ammonium (NH4+) to nitrite (NO2−) in the presence of dissolved oxygen (DO). As of the writing of this article, five recognized genera of AOB in two phylogenetically distinct groups have been reported: Nitrosomonas (e.g., N. eutropha, N. europaea), Nitrosospira, Nitrosovibrio, Nitrosolobus, and Nitrosococcus (Junier et al., 2010). ANAMMOX bacteria are able to produce N2 gas from NO2− and NH4+ under anoxic conditions. Similar to AOB, five genera of anammox have been reported: Brocadia, Scalindua, Kuenenia, Jettenia, and Anammoxoglobus (Strous et al., 1999). All five bacteria were found in freshwater systems and wastewater treatment plants except Scalindua, which was enriched from marine sediments (Kartal et al., 2012). As none of these species have been isolated as pure culture, genome information and growth physiology of these enriched cultures are still under way.
PN/A installations have been commissioned globally in excess of 100 units primarily for sidestream treatment of anaerobic digester reject water (Lackner et al., 2014). In recent years, there has been a trend toward moving from two stage (i.e., nitritation and anammox happening in separate bioreactors) to single stage (i.e., nitritation and anammox simultaneously working in one reactor) as the PN/A configurations offer reduced footprints and better operational control. Also, efforts are in place to evaluate the possibility to employ PN/A technology for mainstream nitrogen management. In applying the PN/A process to wastewater treatment plants (WWTPs), the two most commonly installed reactor configurations are suspended growth (e. g. DEMON®) and attached growth (e.g., Anita™Mox). The spatial distributions and the activity of AOB and anammox bacteria in the suspended growth and attached growth have been investigated and reported. AOB are active in the outer oxic region of biofilms or aggregates, while ANAMMOX is protected in the inner anoxic region (Vlaeminck et al., 2010; Gilbert et al., 2013; Persson et al., 2014; Chu et al., 2015). However, a close comparison of reactor performance and microbial community in single stage suspended and attached growth reactors under similar feed composition is missing.
Although so many installations have been implemented, it is still unclear how the reactor configuration and temperature impact the selection of anammox species. Reactor configurations and temperatures may have an impact on the flanking species involved in the nitrogen transformation in PN/A systems. This impact may also be different in suspended and attached growth systems even when those units are operated under similar conditions. Besides AOB and anammox bacteria, other heterotrophs widely found in these systems include Ignavibacteriacea (Gonzalez-Martinez et al., 2014; Zhang et al., 2014), Anaerolinea (Gilbert et al., 2014; Chu et al., 2015), and denitrifiers Rhodocyclaceae, Comamonadaceae (Gilbert et al., 2014), and their role and dynamics in different reactor configurations and under different temperatures have not been reported.
In PN/A systems, other phenotypes such as nitrite oxidizing bacteria (NOB) may exist and compete with ANAMMOX for nitrite. Typical are Nitrobacter and Nitrospira (Persson et al., 2014). To outcompete the NOB in the system, PN/A reactors are generally operated at temperatures higher than 25°C. Higher temperatures also facilitate the growth of slow-growing anammox bacteria. Although PN/A systems offer the advantage of energy saving, operating these systems at mesophilic temperatures requires heat energy to sustain these configurations. Since both suspended and attached growth (i.e., AnitaMox™ type) configurations are being practiced for PN/A systems, it is also important to compare the performances of both systems at normal operating and lower (room) temperatures. Examining the performance of PN/A system at lower temperatures is important, especially given the fact that PN/A systems are now being investigated for mainstream nitrogen management. The overall objective of this research was to evaluate the performance and microbial ecology of attached growth and suspended growth PN/A reactors when operated at room (21°C) and mesophilic (35°C) temperatures.
Materials and Methods
Suspended and attached growth reactor startup and operation
A 9 L suspended growth PN/A rector (hereafter called the SR reactor) was inoculated with biomass collected from a single-stage DEMON at HRSD York River Treatment Plant (Seaford, VA) to maintain the initial mixed liquor suspended solid concentration of 3,000 mg/L. Anaerobic digester filtrate from a local wastewater treatment plant (North Davis Sewer District, Syracuse, UT) was used as the feed solution to the SR reactor. Fresh centrate was collected from North Davis sewer district biweekly and stored in cold until further use. Each batch of fresh centrate was characterized before its use as a feed for PN/A reactors. NH4+ concentration in the filtrate was 406 ± 23 mg-N/L (SD, n = 76). Table 1 shows the feed composition in terms of important parameters. DO concentration was maintained below 0.4 mg/L by intermittent aeration with an aquarium pump. The reactor was operated in sequencing batch mode with four batches per day, yielding an hydraulic retention time (HRT) of 2 days in the beginning and decreasing it to 0.5 days at the steady state. Each batch included 10 min of feeding, 4 h of reaction, 20 min of final aeration as a polishing step to further oxidize remaining ammonia, 20 min of settling, and 10 min of decanting. The 4-h reaction period contained 16 aeration cycles consisting of 10 min of aerobic conditions and 5 min of anoxic conditions. Anoxic conditions were maintained by switching off the aeration. There was nearly a 1 h of waiting time to collect the feed from the nitritation reactor in between each cycle. The SR was maintained at 35°C for 122 days by wrapping the reactor with silicone heating tape (BriskHeat, Columbus, OH). Afterward, the heating tape was removed to let the reactor sit at room temperature of 21°C for 183 more days.
AR, attached growth; SR, suspended growth.
Along with the suspended growth PN/A reactor, an attached growth PN/A reactor (hereafter called the AR reactor) was started and operated with a total volume of 10.5 L, of which 85% was void volume (i.e., effective volume). The AR was filled with round bio-balls (CoraLife Bio Ball Aquarium Filter) as the biofilm growth media, of which roughly 50% of the fresh bio balls were grown with nitrifying population and the rest contained anammox biofilm. The bio-balls with anammox biofilm were obtained from an ongoing attached growth anammox reactor in the laboratory. To form the nitrifying biofilm and accelerate the startup process, fresh bio balls were soaked in suspended nitrifying biomass from a partial nitritation reactor (Kotay et al., 2013) for a week. The bio-balls containing nitrifying and anammox biofilms were randomly filled in the AR reactor and the AR reactor was flushed with real centrate to remove the suspended biomass.
Each bio-balls has a diameter of 2.54 cm and a surface area of 88.9 cm2. In the beginning, 50% diluted filtrate was fed to the AR to thicken the biofilm growing on the bio balls and let the microbial community form biofilm on the freshly attached growth media. The nitrogen loading was slowly increased depending on the reactor performance. Similar to the SR, AR was also operated with four batches everyday. The AR was operated with intermittent aeration by switching the aquarium pump in on and off modes for about half a month. However, it was found that the NO2− concentration after 10 min of aeration was consistently lower than 1 mg-N/L, even when the DO in the AR was maintained at a level similar to the DO in the SR. Following this observation, continuous aeration was applied to the AR PN/A reactor and the settling phase was removed to maximize the reaction time. As a result, each 6-h cycle in the AR consisted of 10 min of feeding at the beginning and 10 min of decanting at the end, with continuous aeration in between. The AR was operated at 35°C for 133 days and then at 21°C for 172 days.
Analysis methods
Influent and effluent samples were collected regularly and filtered with 0.45 μM Millipore membrane filter paper for nitrogen species analysis. NH4+-N in the influent and NH4+-N, NO2−-N, and NO3−-N in the effluent were monitored. NO2−-N and NO3−-N were analyzed using Ion Chromatography, while NH4+-N was measured using High Range Ammonia Nitrogen by the AmVer™ Salicylate Test N Tube™ Method.
Quantitative polymerase chain reaction
Genomic DNA extracted for SR and AR amplicon sequencing were simultaneously used for quantification of all bacterial 16S rRNA, AOB-specific 16S rRNA, amoA, anammox-specific 16S rRNA, and hzsA gene (per mL) using different primer sets, listed in the table. All samples for quantification were analyzed in triplicates along with standards for each run. Standard curves for quantification of all bacterial 16S rRNA, AOB-specific 16S rRNA, amoA, anammox-specific 16S rRNA, and hzsA genes were constructed with plasmids containing inserts, taking into account the plasmid DNA molecular mass, including the insert length, and the concentration. Copy number for functional gene hzsA for anammox and amoA for AOB were normalized based on anammox-specific 16S rRNA and AOB-specific 16S rRNA, respectively. Each reaction (20 μL) contains 1 μL of genomic DNA, 10 μL of SYBR Green master mix (ThermoFisher Scientific, CA), 1 μL of bovine serum albumin, 1 μL of each primer (forward and reverse), and others nuclease-free water. The quantitative polymerase chain reaction (qPCR) conditions for each reaction was applied according to the references listed in Table 2.
DNA extraction and amplicon sequencing
For the SR, mixed liquor samples were harvested and used for DNA extraction. For the AR, 10 bio balls were randomly collected and carefully washed twice with deionized water. Biomass was carefully scraped off using autoclaved pipette tips and then resuspended in deionized water. The biomass was vortexed for at least 5 min until the big flocs were evenly dispersed. DNA was extracted from the biomass using a PowerSoil DNA Isolation Kit (MoBio, Carlsbad, CA) as described in the manufacturer's protocol. The nucleotide concentration was examined on a Nanodrop spectrophotometer (Nanodrop Technologies, Inc., USA). Then, the DNA concentration was normalized to ∼20 ng/μL and was sent to Research and Testing laboratory, TX, for MiSeq Illumina sequencing. The primers used for 16S rRNA V4 region amplification were 515F (5′-GTG CCA GCM GCC GCG GTA A-3′) and 806R (5′-GGA CTA CHV GGG TWT CTA AT-3′).
Raw reads obtained were interleaved using PEAR Illumina paired-end read merger (Zhang et al., 2014). Low-quality sequences with Phred Scores below 25 were removed through an internally developed quality trimming algorithm. After being trimmed for quality, reads were sorted by length from longest to shortest. Prefix dereplication and clustering at 4% divergence were performed using USEARCH algorithm (Edgar, 2013), wherein sequences shorter than 100 bp were ignored, with no minimum cluster size to allow singleton clusters in the output. All clusters containing <2 members were removed from the datasets. The UPARSE OTU selection algorithm (Edgar, 2013) was used to classify a large number of clusters into OTUs. Chimera check was performed on each selected OTU using UCHIME (Edgar 2013). To determine the taxonomic information, the sequences were run through the USEARCH global alignment program (Bokulich et al., 2014), wherein quality filtered reads were mapped to their corresponding nonchimeric clusters. The output generated was then assigned to a taxonomic profile using an internally developed script.
Results and Discussion
Performance of SR and AR at 35°C
Operation of the SR can be divided into two phases based on the temperature maintained in this reactor. As illustrated in Fig. 1a and Table 3, during Phase I, where the temperature was maintained at 35°C, the HRT was decreased from 2 days to 1 day and finally to 0.5 days. This adjustment was done by increasing the influent/effluent volume that was loaded and drawn out of the reactor. At the same time, aeration was gradually increased to improve the AOB activity and eventually to provide more NO2− for anammox. The average DO concentration in the mixed liquor was maintained around 0.4 mg/L. Due to this change, the NO2− in the effluent significantly increased and an NO2− concentration as high as 19.8 mg-N/L on the 110th day was recorded. Such a buildup of NO2− might have occurred due to the high AOB activity and/or low activity of anammox bacteria. Based on the fact that AOB and NOB mainly grow as tiny flocs, while ANAMMOX tends to grow as large granules, resulting in different settling characteristics (Nielsen et al., 2005; Vlaeminck et al., 2010; Hubaux et al., 2015), a shorter settling time of 5 min during each batch around the 27th day was applied to improve the washout of nitrifiers. From the 57th day of operation, the NH4+ concentration in the final effluent began to increase. After checking the pH and alkalinity in the influent and aeration pump, it was determined that the reactor was alkalinity limited. Thereafter, an equivalent amount of CaCO3 (added as NaHCO3) was provided in the influent. Following this change, the NH4+-N removal efficiency increased from 56.5% at day 58 to 82.4% at day 72. The inadequacy of alkalinity occurred during the gradual increase in the removal rate to 0.49 kg-N/(m3·d−1) with a pH of 7.8. It is not yet known if the higher pH was due to a reduced metabolic activity of microbes in the system or because of the limited availability of carbon.
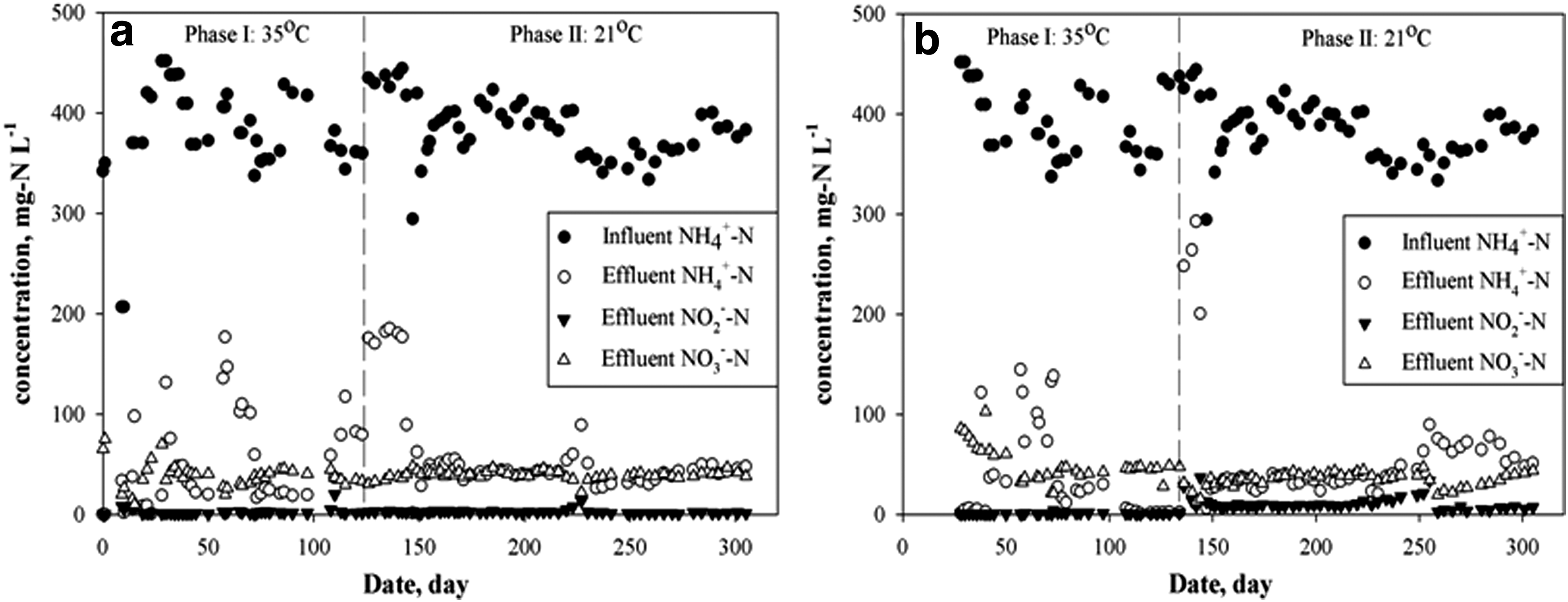
Variation of influent and effluent concentrations of ammonium, nitrite, and nitrate for
HRT, hydraulic retention time.
The operation of the AR can also be divided into two phases based on the temperature gradients provided to this PN/A system. During the initial 27 days, to allow development and growth/maturing of biofilms on the surface of media, the reactor was operated manually and aeration was adjusted frequently (data not shown). From the 28th day of operation, the reactor was fed with the same reject water as in the SR reactor. Similar to the SR PN/A reactor, NH4+ in the effluent increased when the HRT was decreased on days 38 and 57. The details of the ARs performance are listed in Table 2. When the AR was operated at 35°C, NO2− in the effluent never exceeded 5 mg-N/L. The ratio of NO3−-N production to NH4+-N depletion was 0.17, which is higher than the reported ratio of 0.11 in single-stage PN/A systems (Strous et al., 1999). This might be due to the presence of NOB in the AR, where a long solid retention time was maintained, resulting in an inadequate washout of NOB from the system. Moreover, the continuous aeration strategy may also have favored the growth of NOB. Due to the limitation of available alkalinity on the 72nd day, the temperature shock was delayed till a steady state was reached.
Performance of SR and AR at 21°C
At the 123rd day of operation, a temperature shock was provided to the SR reactor by reducing the temperature to 21°C. Following this, the NH4+ in the effluent increased significantly, while NO2− and NO3− remained stable, indicating a significantly decreased AOB activity in the SR PN/A system. To investigate the anammox activity change, ex situ tests were performed in triplicate under anaerobic conditions. The results indicated a decrease in the specific activity of anammox by ∼40% during the temperature shock from 35°C to 21°C (raw data not shown). This could have been due to (1) temperature shock to anammox bacteria and (2) limited availability of NO2− due to suppression of AOB activity. The suppression of anammox activity suggests that NH4+ removal rate could be further improved if the AOB activity is improved. During the next 20 days of the acclimatization stage, the average TIN removal was 0.46 kg-N/(m3·d−1). By the 144th day, the NH4+ concentration in the effluent slowly dropped to 89.2 mg-N/L, yielding a TIN removal of 0.57 kg-N/(m3·d−1). Afterward, the reactor performance was stable with an average TIN removal rate of 0.59 kg-N/(m−3·d−1), indicating a successful adaptation to room temperature by the microbial communities performing PN/A in the SR.
In contrast to the SR, NO2− accumulation and NO3− depletion in the effluent were observed in the AR for over the next 2 days right after the decrease in temperature. The nitrite and ammonium accumulation indicated that both the AOB and anammox activity decreased due to cold temperature shock. The build down of NO3− in the effluent resulted from the decreased activity of anammox, in which case, less NO3− was produced by oxidation of NO2− for biomass synthesis (Kartal et al., 2012). The AR reactor recovered from the cold temperature shock in just 10 days (days 135–144). The TIN removal rate increased from 0.24 kg-N/(m−3·d−1) on day 136 to beyond day 147, finally achieving a constant TIN removal of 0.58 kg-N/(m−3·d−1) with 89% of NH4+ removal efficiency consistently recorded in the AR PN/A reactor. This result indicates that the PN/A system can be operated at room temperature with high removal efficiency.
qPCR results
Ammonia monooxygenase (amoA) is an enzyme, which catalyzes the first step of ammonia oxidation to hydroxylamine by AOB (Hollocher et al., 1981). This functional gene has been used as a biomarker to understand AOB community at a fine scale (Rotthauwe et al., 1995). In anammox metabolism, the hzsA gene cluster is responsible for the synthesis of hydrazine from ammonium and nitric oxide (Strous et al., 2006). This gene cluster is not present in other nitrogen cycling bacteria and, as a result, hzsA is a very good biomarker for anammox bacteria (Harhangi et al., 2012). Biomarkers targeting amoA and hzsA were used to quantify AOB and anammox bacteria in both reactors under different temperatures.
The ammonia monooxygenase (amoA) functional gene copy numbers were 1.25 ± 0.08 × 109 and 1.85 ± 0.16 × 109 in SR and AR, respectively, at 35°C. The amoA gene copy numbers were 0.78 ± 0.11 × 109 and 1.9 ± 0.09 × 109 in SR and AR, respectively, at 21°C, indicating a negligible change in amoA gene copy numbers with temperature decrease. Likewise, the hzsA gene copy numbers were 5.36 ± 0.58 × 108 and 1.65 ± 0.70 × 108 in SR and AR, respectively, at 35°C and, 9.72 ± 2.02 × 108 and 8.62 ± 0.040108 in SR and AR, respectively, at 21°C. It is clear that the total number of amoA and hzsA did not change significantly and these findings align well with reactor operation details, in which case, the total inorganic nitrogen removal efficiencies also did not change appreciably. In addition, no appreciable changes were recorded in functional gene copy numbers, which could be due to slow growth of anammox and AOB and, short durations of temperature changes.
Microbial communities analyzed by amplicon sequencing
The primer set 515F/806R targeting the V4 hypervariable region of the 16S rRNA gene yields accurate phylogenetic information and has few biases against any bacterial taxa (Bergmann et al., 2011). So far, these primers have been used to reveal considerably accurate microbial community structure in PN/A reactors (Gilbert et al., 2014). The Illumina sequencing (Illumina, CA) yielded a total of 16,463 to 19,798 short reads for four samples (Table 4). To compare the species abundance between both samples, OTUs were picked to reflect the bacterial species richness. The species richness ranged from 344 to 398 (Table 4). Shannon, chao1, and Simpson indices were calculated to evaluate the diversity of the species at different temperatures in both reactors. An increase in bacterial diversity in both reactors occurred with decreasing temperature (Table 4). To evaluate whether the produced data were enough to cover all species as well as to represent species abundance, rarefaction curves were plotted (Fig. 2). The nearly asymptotic nature of the curves as the number of reads on the x-axis increased demonstrate that the number of reads obtained for all samples from sequencing provided sufficient coverage of the overall microbial communities.
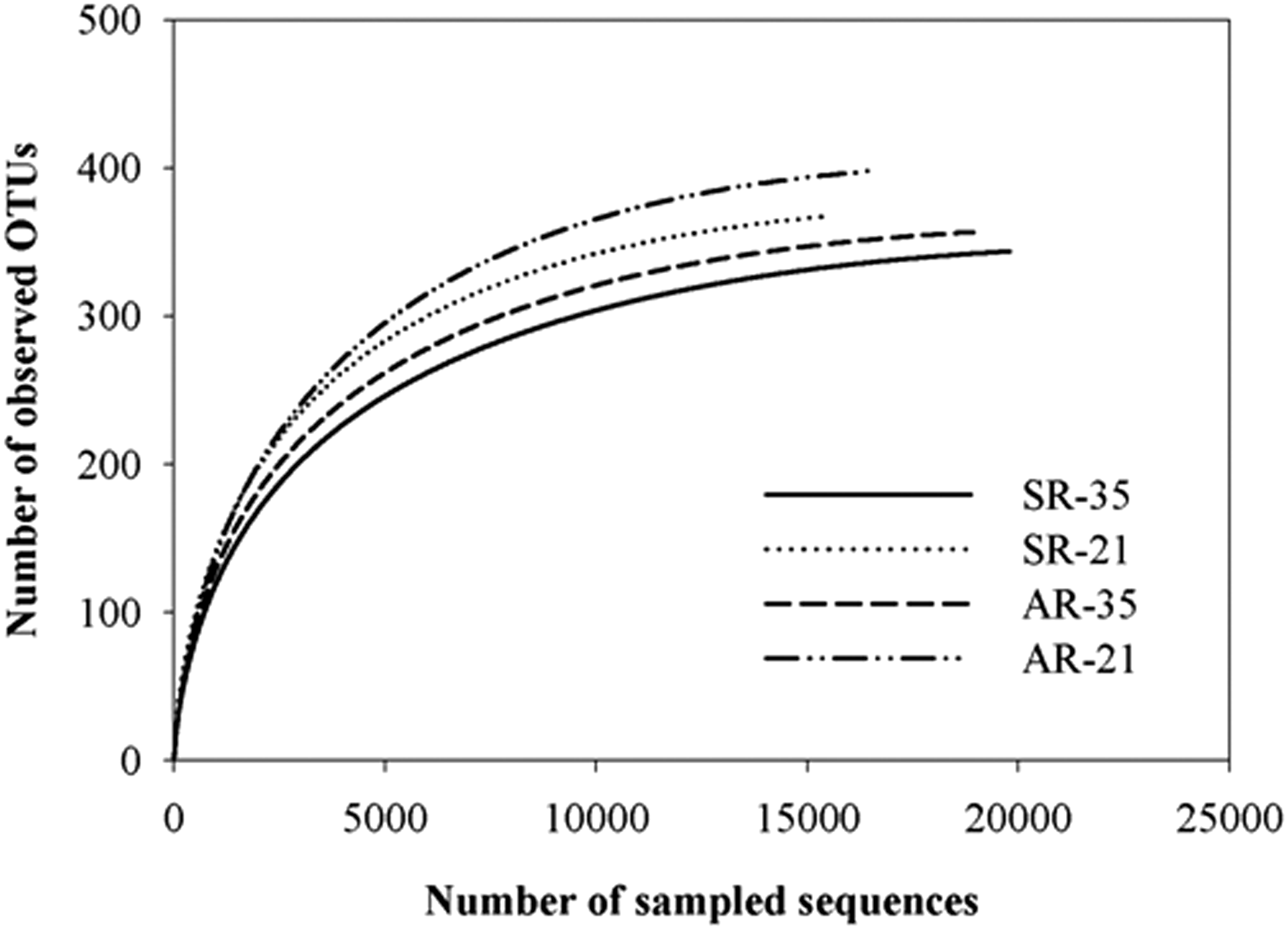
Rarefaction curve of sequencing results from two reactors (SR and AR) under 35°C and 21°C.
Microbial community results are illustrated in Fig. 3 for both reactors at different temperatures. The dominant phyla observed in all four samples were Chloroflexi, Planctomycetes, Bacteroidetes, Verrucomicrobia, Acidobacteria, and Proteobacteria. In general, the percentage of Chloroflexi decreased in both reactors when the temperature decreased from 35°C and 21°C. For the SR, the relative abundance of Chloroflexi and Planctomycetes decreased from 31.4% to 25.0% and from 22.2% to 15.1%, respectively. On the other hand, in the SR reactor, the relative abundance of Proteobacteria increased from 22.9% to 29.3%, Bacteroidetes from 3.0% to 9.1%, and Verrucomicrobia from 0.19% to 1.1%. In the case of AR, when it was operating at 35°C, the most abundant species was Chloroflexi, which accounted for 54.1% of the total reads. Surprisingly, the abundance of Planctomycetes was only 16.4% in the AR reactor at 35°C.
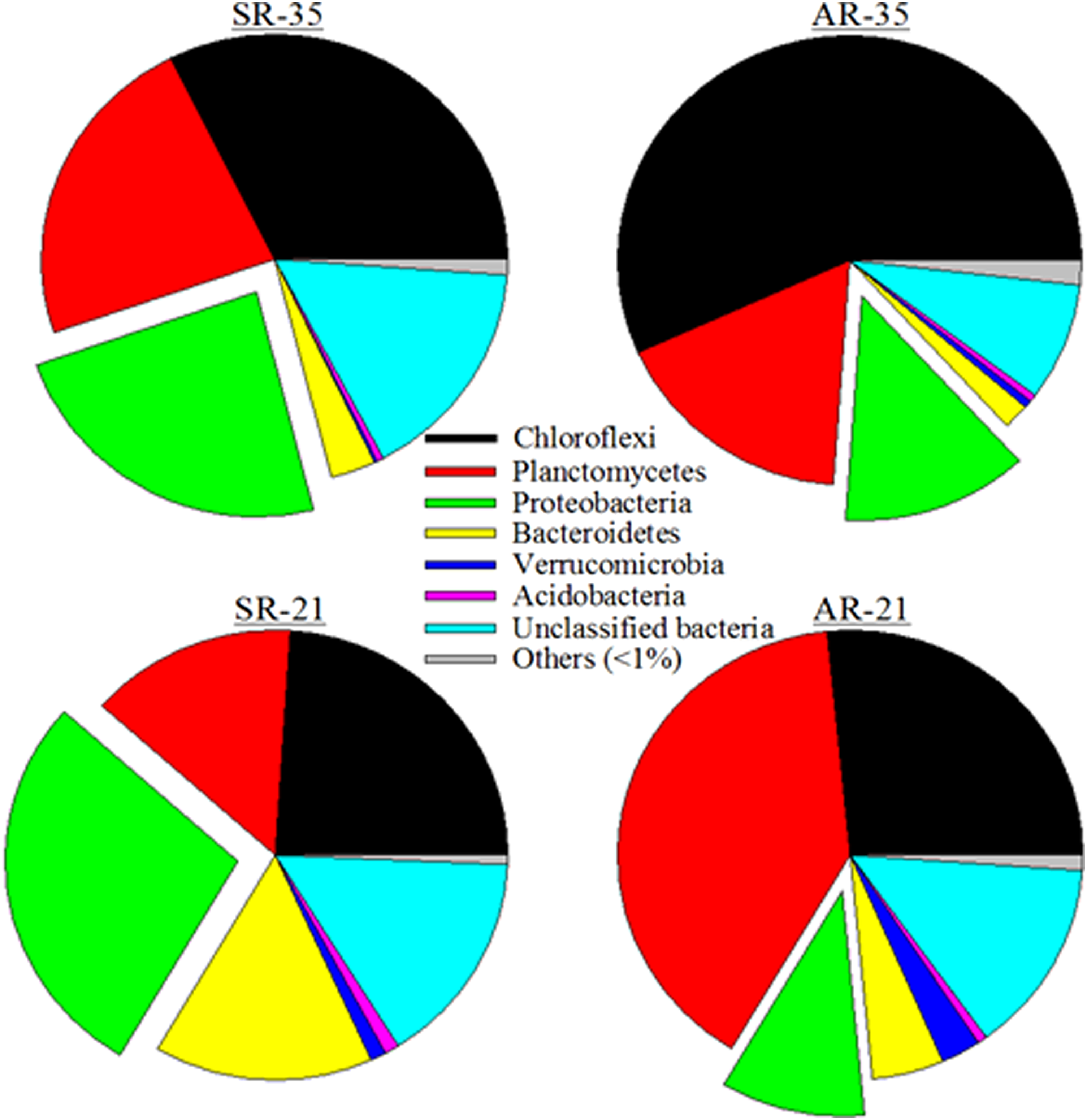
Relative abundance of different phyla in the two reactors (SR and AR) operated under 35°C and 21°C.
During ∼300 days of operation, the SR and AR were able to remove (1) 0.54 ± 0.093 and 0.60 ± 0.111 kg-N/(m−3·d−1) at 35°C and (2) 0.57 ± 0.061 and 0.56 ± 0.103 kg-N/(m−3·d−1) at 21°C, respectively. The similar performance of SR and AR systems might be a result of the similar microbial communities, represented in both reactors by three phyla: Planctomycetes, Chloroflexi, and Proteobacteria. In addition, the morphology of suspended growth granules/flocs and attached growth biofilms in PN/A system has been reported to be similar with the internal anammox zone shielded by an outer AOB rim (Vlaeminck et al., 2010; Gilbert et al., 2013; Persson et al., 2014; Chu et al., 2015). This morphological structure ensures minimum exposure to DO for anammox, enabling them to be active even under aerobic conditions. In this research, the AR was operated under continuously aerated conditions, indicating that the “protection” of anammox is more effective in attached growth PN/A configurations.
Effect of temperature on key individual phylum
In this study, a single species of anammox, Brocadia fulgida, accounted for more than 99% of all species under Planctomycetes for all four samples. The dominant species in the original seed to the AR reactor had Kuenenia stuttgartiensis as the dominant anammox species. However, B. fulgida was the dominant anammox bacteria in the AR, indicating the community selection in the biofilm reactor. The relative abundance of Planctomycetes could be used as the indicator of B. fulgida. As shown in Fig. 3, with the decrease in temperature, the amount of Brocadia in the SR decreased from 18.7% to 10.8%, in contrast to the AR wherein Brocadia grew from 13.2% to 33.7%. It is worth mentioning that the two reactors were seeded with different sources of biomass, while the dominant anammox species for both reactor configurations were “Candidatus B. fulgida.” This suggested that “Ca. B. fulgida” might be selective over other anammox species in single-stage systems fed with the same feed. Corroborating our finding, similar results have been also reported by other researchers who concluded that Brocadia sp. adapts better in engineered ecosystems because of their lower affinity for ammonium nitrite and higher tolerance to DO with higher growth rate (Hendrickx et al., 2014). Moreover, Brocadia sp. is able to thrive at elevated COD concentrations and can coexist with heterotrophic bacteria (Kartal et al., 2008).
Various orders of species under Proteobacteria were detected as Burkholderiales, Nitrosomonadales, Rhodocyclales, and a group of uncultured gamma-Proteobacteria. Figure 4 shows how different Proteobacteria species occupied the phylum in both the reactors. When operated at 35°C, N. europaea, classified under Nitrosomonadales, was found to be the responsible AOB in both the SR (3.15% of total reads) and the AR (3.41% of total reads). However, when the temperature was decreased to 21°C, another Nitrosomonas sp. was detected together with N. europaea for aerobic ammonium oxidation. This might indicate that the emerging Nitrosomonas sp. is more tolerant to lower temperature, although more evidence is needed for a definitive conclusion. Bukholderiales and Rhodocyclales were regarded as heterotrophs able to denitrify nitrate to nitrogen gas under an anoxic condition with an organic carbon source. The presence of denitrifying bacteria in both the SR and AR might be due to the organic carbon present in the influent and the organics produced from the decay of cells.
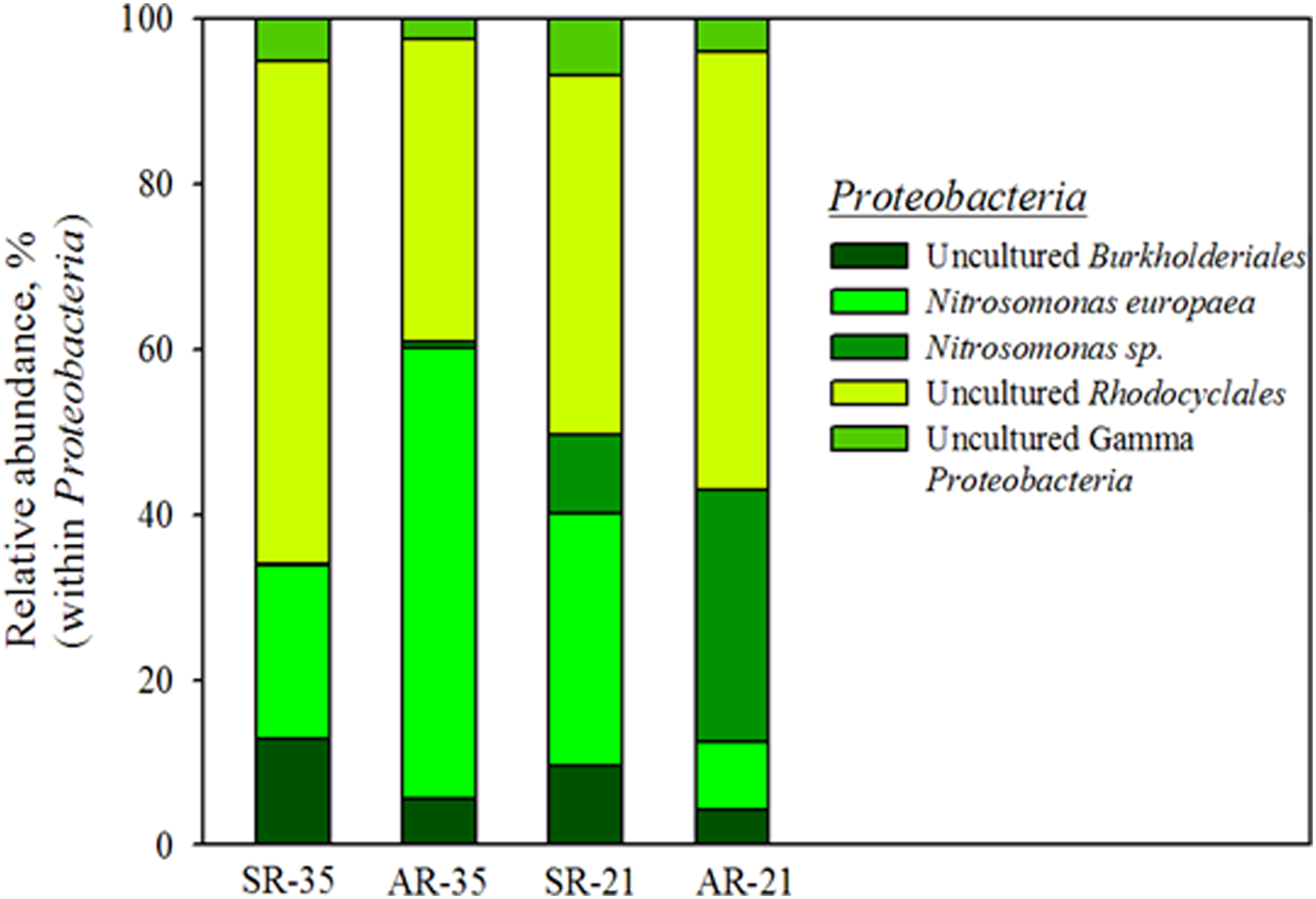
Relative abundance of the different species within Proteobacteria.
In the biofilm system, Nitrosomonas, which has low-affinity constant for NH3, might have a growth advantage over other AOB (Karkman et al., 2011). Moreover, previous studies showed an increase in nitrifying biofilm thicknesses with exposure to cold temperatures in a nitritation system (Bjornberg et al., 2009; Delatolla et al., 2012). In this case, the total Nitrosomonas population did not increase significantly in the AR over the 300 days of operation. It can be concluded that in PN/A systems, where ANAMMOX grow inside the floc or the granule/biofilm and AOB grow outside, the thickness of the outer layer of AOB was constant due to the shear force of air flow. In SR, the doubling of Nitrosomonas population and the decrease of Brocadia possibly hint that washing away the excess AOB from the system by shortening the settling time may not be efficient. However, in full-scale applications, which generally operate over multiple years, a more efficient sludge separation method (i.e., hydrocyclone) is necessary. DO can be another factor controlling the process as, by limiting the ammonium oxidation rate, it ensures a balance in the activity of AOB with ANAMMOX.
With the decrease in temperature, an increase was observed for a different Nitrosomonas sp. along with the earlier N. europaea. Similar results have been reported by Gilbert et al. (2014) highlighting the coexistence of Nitrosomonas sp. JL21 with N. europaea when the temperature of a PN/A reactor gradually decreased from 20°C to 10°C. This indicated that N. europaea might be less tolerant to cold condition. The emerging Nitrosomonas sp. is in the same branch as N. nitrosa, which was reported to have an advantage in environments with fluctuating oxygen levels or during the absence of oxygen (Limpiyakorn et al., 2005).
Under the phylum Chloroflexi a highly dominant species of uncultured Anaerolineales was detected ranging from 83.1% to 89.6% in both reactors with the remainder being unknown Chloroflexi species. The uncultured Anaerolineales found in this study was closely related (100% homology) to an uncultured Chloroflexi, which was isolated from an anammox biofilm reactor reported by Kindaichi et al. (2012). So far, Chloroflexi has been reported in many PN/A bioreactors, including suspended growth systems (Gonzalez-Martinez et al., 2014; Chu et al., 2015) and attached growth systems (Park et al., 2010; Gilbert et al., 2014). The species classified under the phyla Chloroflexi belonged to the order Anaerolineales. These heterotrophic species are known to be thermophilic with a relatively slow growth rate and a need to associate with other microbes for efficient growth (Yamada and Sekiguchi, 2009). Based on the fact that in any bioreactor, including PN/A systems, soluble microbial products, and extracellular polymeric substances are inevitable (Laspidou and Rittmann, 2002), the presence of Anaerolineales is not a surprise. The carbon source for species belonging to phyla Chloroflexi was probably from the decay of bacterial cell materials in the system (Kindaichi et al., 2012; Ni et al., 2012) and the COD present in the influent. A decline in abundance of Anaerolineales species for both the reactors was observed as the result of the low temperature. In the SR, Anaerolineales decreased from 27.25% to 22.35% and in the AR, Anaerolineales declined from 49.97% to 22.17%, suggesting that uncultured Anaerolineales are sensitive to lower temperature.
Compared to suspended growth, the attached growth system maintains an extremely long solid retention time, thus, the higher decay rate of microorganisms accelerates the growth of Chloroflexi. This may explain why the highest abundance of Chloroflexi was found in the AR when operated at 35°C. At room temperature, the growth of the thermophilic bacteria was limited by the lower temperature, which resulted in the decrease of the relative abundance of Chloroflexi in both the SR and AR at low temperatures. Furthermore, Chloroflexi has been reported to be the backbone for the three-dimensional microbial aggregates called flocs and to reinforce the granule structure in suspended growth systems (Cho et al., 2010). Chloroflexi is also involved in the spatial organization in the biofilm of attached growth systems (Botchkova et al., 2014). Therefore, it could be inferred that at the beginning of the operation, the relatively high temperature favored the growth of Chloroflexi, which in turn involved in the growth of biofilm in the AR. Overall, irrespective of whether AR or SR, Chloroflexi was the species responsible for creating the structure in the PN/A system. However, the role of Chloroflexi on the ammonium removal rate of the reactors remains to be explored.
Besides Chloroflexi, other microbes present in the PN/A reactors also compete for soluble microbial products and extracellular polymeric substances. These microbes include denitratisoma and Ignavibacterium. Denitratisoma are denitrifiers able to use complex organics as their carbon source to reduce nitrate to nitrogen gas. However, the ratio of produced NO3− to removed NH4+ in both reactors did not show any remarkable sign of heterotrophic denitrification. The reported value of 0.11 for the molar ratio of NO3− production to NH4+ depletion was recorded and no NOBs were detected in either system. Ignavibacterium album has been proposed as a facultative anaerobe, moderately thermophilic, and a heterotrophic fermenter (Liu et al., 2012). Moreover, this bacterium has been reported carrying the gene NifAH, which encodes for the key enzyme responsible for dissimilatory nitrate reduction to ammonia (DNRA) (NCBI 2014 gene IDs for A. thermophile nrfA, nrfH: 10172349, 10171709; I. album strain JCM 16511 NC_017464 nrfA: NC_017464 region: 957373-958887) (Liu et al., 2012). This finding suggested that this bacterium has the potential to perform DNRA when an appropriate condition is provided (i.e., the absence of oxygen, and available nitrate and organic substrates (Sgouridis et al., 2011)). Further research is needed to confirm the metabolic pathway of Ignavibacterium.
In summary, we suspended growth (SR) and attached growth (AR) systems had similar microbial communities, which enabled both reactors to perform with a similar high nitrogen removal rate during the 300 days of operation at 35°C and 21°C. In the long run, the nitrogen removal rate in the PN/A systems did not deteriorate when the operation temperature decreased to 21°C. In fact, both the reactors adapted to the lower temperature within 2 weeks, showing that in full-scale application, it is feasible to bring down the bioreactor temperature to reduce the operation cost.
Conclusions
This article demonstrated the performances of attached and suspended growth single stage PN/A reactors at two different temperatures. The microbial ecology in both reactors was also investigated using high-throughput sequencing. The microbial community analysis not only focused on key players, that is, ammonia oxidizers and anammox bacteria but also included other flanking community generally present in PN/A systems. The results convincingly showed that the performances of both reactors in terms of total nitrogen removal and microbial ecology were nearly identical. The removal efficiencies decreased at a lower temperature. Although the ecology of flanking community was established in this research, their ecophysiology in PN/A reactors remains a subject of further research. Figure 5 shows the overall scheme of the concept tested.

Performance and community comparisons of suspended and attached growth PN/A systems.
Footnotes
Acknowledgments
In advance, we appreciate the editor and the reviewers for their time and useful comments. This research was partially funded by North Davis Sewer District in Utah. The views and opinions expressed in this article are those of authors and do not necessarily relate to the funding agency.
Author Disclosure Statement
No competing financial interests exist.