
Abstract
Abstract
Our previous work (published in Letters in Applied Microbiology 2013) showed that Burkholderia phytofirmans candidate strain ST01hv can degrade thiocyanate (SCN−), a common contaminant from gold ore processing. However, the impacts of major environmental parameters on the biodegradation activity of this strain must be characterized to implement it in bioremediation efforts. In this study, several environmental factors, including pH, the concentrations of SCN−, phosphate, and ammonium, and the type and concentration of organic carbon, were tested for effects on the efficiency of SCN− biodegradation by the bacterium. Results showed that SCN− biodegradation occurred within a range of pH values (6.0 ± 0.2 to 8.0 ± 0.2), with an optimum between 7.0 and 7.5. SCN− breakdown depended on initial ammonium and SCN− concentrations, as well as the availability of phosphate. At lower ammonium and SCN− levels (<60 and <220 mg/L, respectively), the extent of biodegradation was greater, while at higher concentrations (≥60 and 350–1,500 mg/L), these substances acted as inhibitors. Different carbon substrates showed different degrees of influence, with glucose (4.0 g/L, the highest amount of available C at 133 mM C/L medium) providing the best results (i.e., complete SCN− breakdown after 4 days). The tested environmental parameters are relevant to industrial environments such as mining- and coking-contaminated soils, and the results from this work will inform strategies for SCN− bioremediation.
Introduction
T
Biological treatment has been proven to be the most cost-effective and environmentally sustainable strategy for dealing with SCN− contamination (Dzombak et al., 2006; Dash et al., 2009; Gould et al., 2012). A range of microorganisms can use SCN− as their carbon, energy, nitrogen, and sulfur source (Gould et al., 2012). The end products of SCN− bioremediation are ammonium (NH4+), carbon dioxide, and sulfate (SO42−). Two pathways for SCN− degradation have been described, with the intermediate compounds carbonyl sulfide and cyanate, respectively (Sorokin et al., 2001; Ebbs, 2004; Gould et al., 2012). The ability to biodegrade SCN− has mainly been studied with Thiobacillus or halophilic/alkaliphilic genera (Ebbs, 2004; Gould et al., 2012); however, one study used Burkholderia under alkaline pH conditions (Adjei and Ohta, 1999).
In a previous study, we showed that Burkholderia phytofirmans, candidate str. ST01hv, isolated from SCN−-contaminated soil from an Australian gold mine, can aerobically and heterotrophically degrade SCN− at neutral pH (Vu et al., 2013). The bacterium also appears to be able to reduce ambient sulfate levels when SCN− has been fully consumed, possibly via assimilation. This latter capability is of interest as sulfate is typically an end product of SCN− degradation that can lower pH in mine tailings or groundwaters. Optimization of bioremediation efforts involving Burkholderia requires detailed assessments of the effects of important environmental parameters on metabolic activity. In this study, we experimentally quantified the effects of pH, SCN−, ammonium, and phosphate concentrations, and organic carbon substrate type on the growth, metabolism, and SCN− degradation capacity of B. phytofirmans candidate str. ST01hv.
Experimental Protocols
All experiments (unless otherwise stated) were conducted using a broth medium for cultivation (Kwon et al., 2002; Vu et al., 2013), with a 50% reduction in the previously published phosphate concentration. Specifically, the medium contained (per liter) 200 ± 10 mg SCN− (prepared from NaSCN), 1.0 g acetate (prepared from CH3COONa), 3.0 g Na2HPO4, 1.5 g KH2PO4, 1.0 g NaCl, 0.01 g CaCl2, 0.5 g MgSO4.7H2O, 0.04 g FeSO4.7H2O, and 0.0016 g MnSO4.H2O. SCN− bioremediation experiments were conducted using previously described protocols (Vu et al., 2013). SCN− biodegradation was carried out at 28°C in 250-mL conical flasks with the total volume of 50 mL. The flasks were shaken at 150 RPM (rotations per minute). The degradation experiments were conducted in three replicates, with cultures initially grown in a neutral medium containing acetate (1.0 g/L) as the sole C source. Each culture (1.0 mL) was washed three times with growth medium before transfer into experimental media. The following parameters were investigated: pH from 5.0 to 10.0; SCN− concentration from 50 to 1,500 mg/L; PO43− (prepared from Na2HPO4) concentration from 0.375 to 3.0 g/L; different carbon sources, including glucose (C6H12O6), pyruvic acid (C3H4O3), yeast extract, citrate (C6H5Na3O7*2H2O), and acetate (CH3COONa) at 1.0 g/L each (except for glucose, for which up to 4.0 g/L was tested); and ammonium from 10 to 100 mg/L. SCN− and ammonium concentrations were determined by spectrophotometric (Standard Methods 4500-CN) and turbidimetric (Method 10031) methods, respectively (Eaton and Franson, 2005; Hach, 2014).
Results
Biodegradation of SCN− occurred only between pH 6.0 ± 0.2 and 8.0 ± 0.2; no SCN− degradation was observed at either lower or higher pH values (Fig. 1). The reported SCN− fractions, hereafter, are the ratios of measured SCN− to the initial SCN− concentrations. The optimum pH condition was found to be between 7.0 and 7.5, at which up to 44% of SCN− was degraded after ∼3 days.
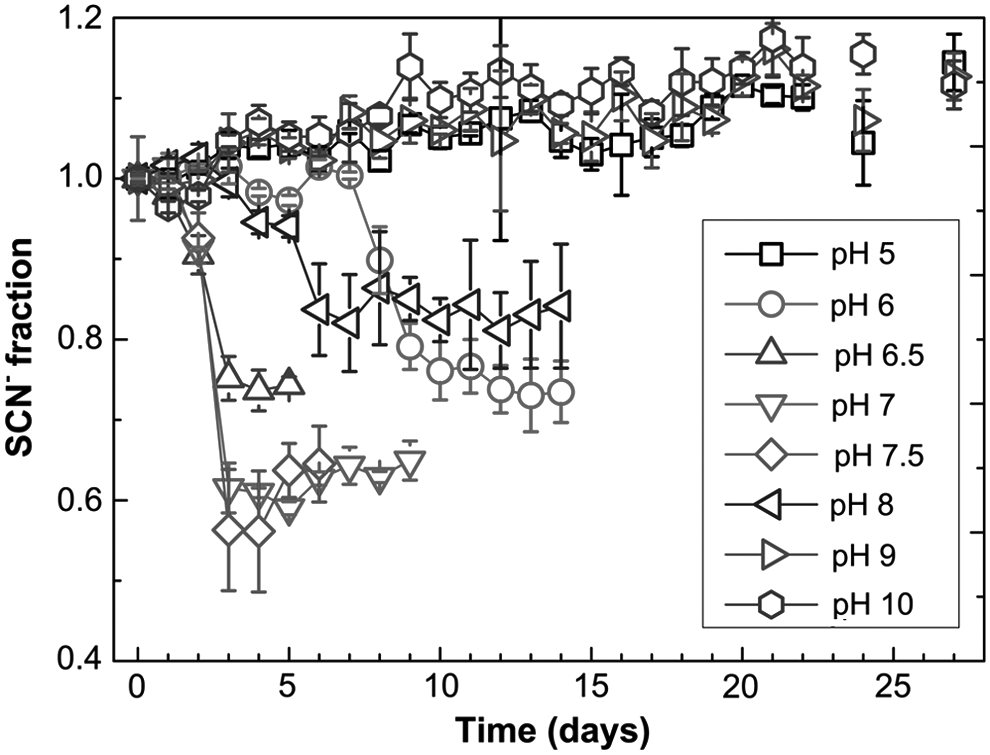
Influence of pH on the SCN− biodegradation by Burkholderia phytofirmans ST01hv.
The dependence of biodegradation efficiency on the initial concentration of SCN− is presented in Fig. 2. The highest proportion of SCN− degradation (up to 70% SCN− degraded) was observed at the lowest initial concentration of SCN− (50 mg/L), while at higher SCN− concentrations (>350 mg/L), the fraction of degraded SCN− decreased significantly (≤10% of SCN− degraded). At SCN− concentrations >350 mg/L, the ability of the bacterium to utilize SCN− as a sole N source seemed to be negligible (Fig. 2).
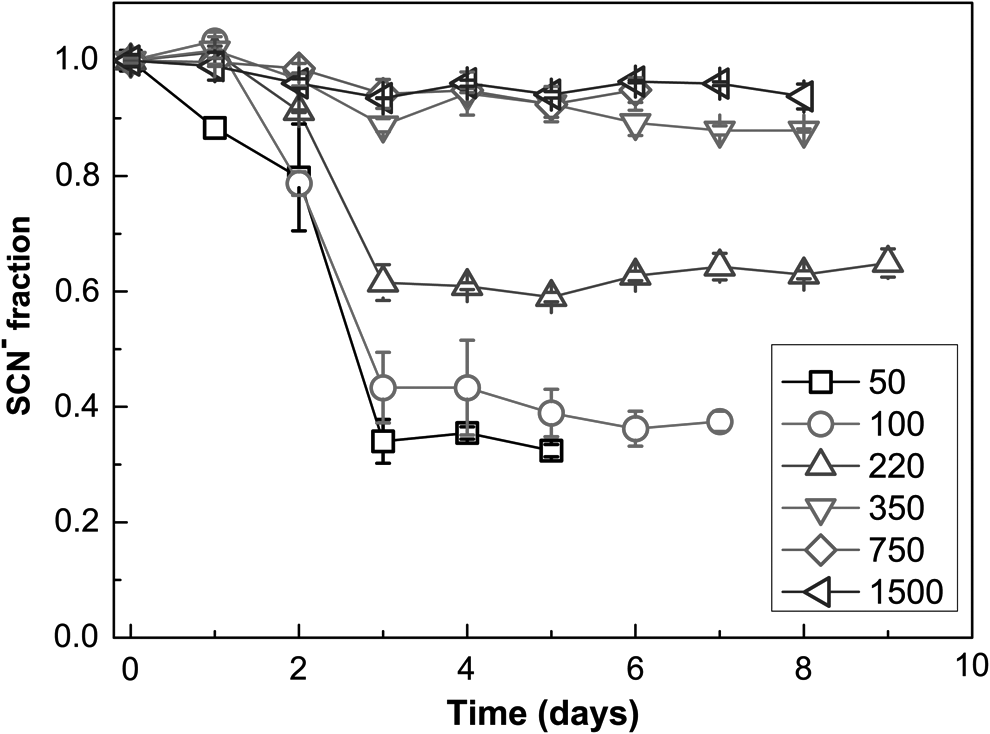
Impact of initial concentration of SCN− on the biodegradation of SCN− by Burkholderia phytofirmans ST01hv. The numbers in the legend represent initial SCN− concentrations in mg/L.
SCN− degradation efficiency also depended on the level of phosphate in the medium (Fig. 3). Biodegradation of SCN− progressed more quickly and efficiently at higher concentrations of phosphate (e.g., at 3.0 g PO43−/L, Fig. 3).
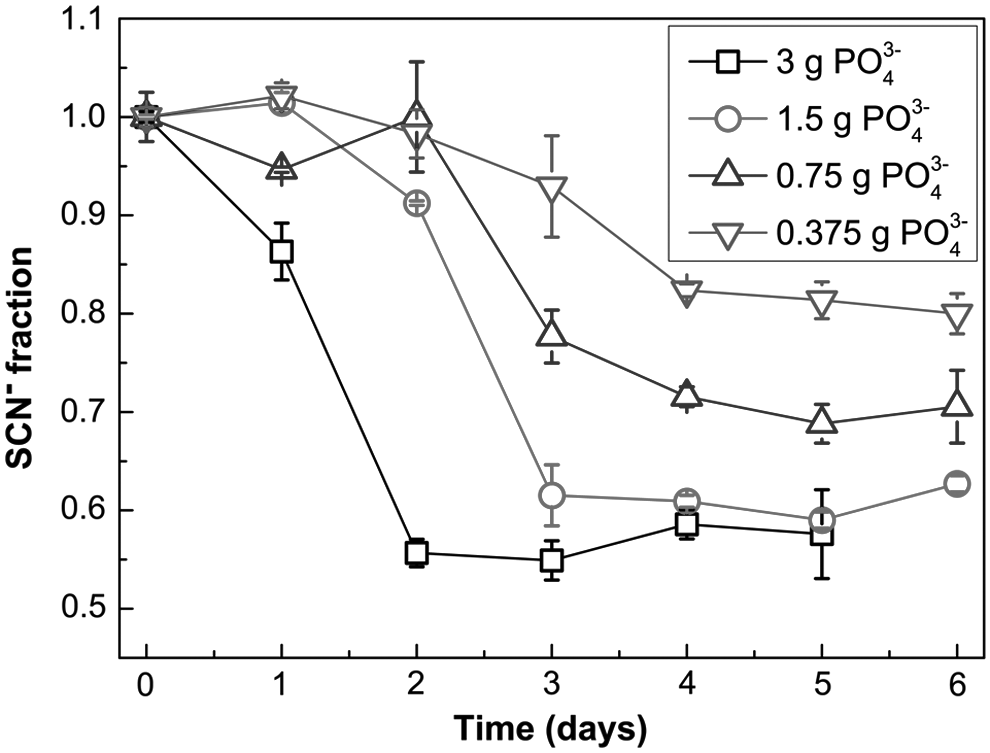
Dependence of SCN− biodegradation by Burkholderia phytofirmans ST01hv on the concentration of phosphate.
Different organic carbon substrates (at 1.0 g/L) exhibited different effects on SCN− biodegradation (Fig. 4, with respective moles of C per substrate also shown). No SCN− degradation was detected in experiments for which yeast extract was used as the sole carbon source. Among the carbon substrates tested, glucose provided the optimal substrate and promoted the greatest extent of SCN− biodegradation (up to 70% SCN− degraded). Degradation stopped after 3 days in experiments using glucose as the sole carbon source (i.e., at 1.0 g glucose/L, Fig. 4). Acetate and pyruvic acid also supported SCN− biodegradation, with the breakdown of SCN− stopped after 3 to 4 days (up to 50% SCN− degraded). The bacterium also utilized citrate as a sole carbon source, but SCN− breakdown required more time (up to 40% SCN− degraded after 10 days) when compared to other experimental carbon substrates (Fig. 4). In the case of glucose, a complete breakdown of SCN− occurred when supplied with 4.0 g glucose/L or 133.3 mM C/L.

Effects of different C sources on SCN− biodegradation by Burkholderia phytofirmans ST01hv.
Ammonium at higher concentrations (>60 mg/L) inhibited SCN− degradation; but at lower concentrations, its presence had less impact on SCN− breakdown (Fig. 5; Supplementary Fig. S1). At lower ammonium concentrations (e.g., a 20 mg/L), the bacterium could utilize both SCN− and ammonium as N sources; however, SCN− degradation either required more time to initiate or the degraded faction was smaller compared to ammonium-free experiments (Figs. 4 and 5; Supplementary Fig. S1).

Effects of ammonium concentrations on SCN− biodegradation by Burkholderia phytofirmans ST01hv.
Discussion
The results showed that the best conditions for bioremediation of SCN− by B. phytofirmans candidate str. ST01 were as follows: (1) a narrow range of pH, consistent with the range over which environmental Burkholderia spp. can degrade cyanide (Adjei and Ohta, 2000; Chen et al., 2003); (2) a low concentration of SCN− and ammonium; and (3) a sufficient amount of phosphate and a suitable carbon source. We observed potential lag periods for cultures under pH conditions outside the range of the growth (inoculating) medium (pH 7.0–7.5; Vu et al., 2013), indicating a possible adaptation requirement before SCN− biodegradation could proceed. We also noted that less SCN− degradation was observed outside this pH range, indicating a real pH dependence for metabolic performance. The inhibition of SCN− biodegradation by ammonium (NH4+) at high concentration reflects the possibility that ammonium was more easily utilized as a nitrogen source, when compared to SCN− (the latter contains a triple bond between C and N). As a result, in experiments with a concentration of ammonium higher than 60 mg/L, SCN− breakdown was not detected. Additionally, the inhibitive effect of SCN− at higher concentrations has been reported in a previous study, which investigated SCN− biodegradation by a coculture (Boucabeille et al., 1994). The requirement of phosphate was expected, as it provides an essential nutrient for microorganisms, and acts as a growth-limiting factor (Willey et al., 2008).
Among selected carbon sources, yeast extract can provide both carbon and nitrogen (Willey et al., 2008). Possibly the bacterium preferentially obtained nitrogen from yeast extract (calculated as up to 8.9 mM N/L) or C from yeast extract might have acted as inhibitor for the SCN− degradation. Thus, no SCN− degradation was observed in the presence of that amendment (Fig. 4), consistent with previous observations (Stafford and Callely, 1969; Mirizadeh et al., 2014; Combarros et al., 2015). Other studies have noted that SCN- was not degraded until ammonium was fully consumed (Stafford and Callely, 1969), or that the bacteria (P. thiocyanatus and C2 strains) could grow, but no SCN− degradation was observed (Mirizadeh et al., 2014; Combarros et al., 2015). We, therefore, speculate that once N from the yeast extract was completely consumed in our experiment, SCN− degradation could proceed. Glucose is considered one of the richest and most preferred carbon sources (Michels, 2002; Morales-Sanchez et al., 2013); thus, with glucose as the sole C source (1.0 g/L or 33.3 mM C/L), the yielded degraded SCN− fraction (up to 70%) was larger compared to ∼50% of degraded SCN− when 1 g/L of pyruvic acid (34.1 mM C/L) was used. In addition, with 4.0 g/L of glucose (133.3 mM C/L), our results showed that complete degradation of SCN− was achieved. This observation is in agreement with the interpretation that the highest growth rate was sustained by the substrate with the largest C yield, leading to the greatest amount of SCN− degradation (Fig. 4). The net SCN− degradation capacities of B. phytofirmans cultures in the presence of pyruvic acid (34.1 mM C/L), acetate (24.4 mM C/L), and citrate (20.4 mM C/L) were similar, but SCN− degradation required a longer time when citrate was used as the sole C source. Interestingly, similar rates of SCN− degradation were observed when acetate and pyruvate were used, despite pyruvate (34.1 mM C/L) having roughly 1.4x the amount of available C as acetate (24.4 mM C/L), indicating a possible metabolic preference for acetate by the Burkholderia strain. Reanalysis of our previous work (Fig. 2 in Vu et al., 2013) with the same strain consuming acetate (1 g/L or 24.4 mM C/L) shows that 1.88–2.16 millimoles of SCN− per 1011 cells/day were degraded.
Our previous work showed that B. phytofirmans ST01hv can heterotrophically degrade SCN− and potentially assimilate sulfate (Vu et al., 2013). In this study, the optimization of SCN− biodegradation to various environmentally relevant parameters (Stott et al., 2001; Dzombak et al., 2006; Vu et al., 2013) allows for better estimation and potential implementation of this microorganism to real settings. For example, careful monitoring of pH range and upper limits to noninhibitory ammonium concentrations will be required to ensure the efficiency of using an aerobic bacterium such as B. phytofirmans.
The outputs of potential SCN− biodegradation systems in situ, sulfate and ultimately nitrate (via ammonia oxidation by other microorganisms), will also need to be considered for the potential to acidify or eutrophy, respectively, downstream ecosystems and applications of mine wastewaters. The application of heterotrophic sulfate-assimilating species, such as B. phytofirmans, in sequential bioreactors, for example, could potentially reduce effluent sulfate levels, and also lead to the downstream presence of organic carbon substrates at levels that may influence or alter the activity or composition of, for example, nitrogen-cycling microbial consortia; such factors must be considered to optimize the design of an SCN− bioremediation system.
Furthermore, some subsets of the organic acids tested in this experiment, such as citric, acetic, and pyruvic, may be bioavailable in SCN− impacted sediments or soils, such that efficient SCN− bioremediation may require primarily just the addition of phosphate. However, phosphate amendments in situ are likely to stimulate a wide range of microorganisms with which species such as B. phytofirmans would have to compete. Ex situ or in situ SCN− biodegradation experiments using native microbial consortia and simple phosphate-only amendments should be the focus of future research in this area.
We note that the microorganism used here was relatively easily isolated from SCN−-contaminated soils at an operational gold mine tailings site. Therefore, we speculate that similar SCN−-degrading species can be recovered locally from most gold mine sites under the optimal pH and ammonium conditions, without the necessity for importing commercially available microbial consortia. However, we note that we did not experimentally test the activity of B. phytofirmans for SCN− biodegradation in the copresence of different potential contaminants (e.g., metals) that might be present in SCN−-contaminated tailings and difficult to predict (as they are dependent partially on the local geological “feedstock” for gold ore processing and extraction); such work might also be a useful focus for further experiments.
Summary
In this study, we tested environmental parameters that are relevant to conditions in soils, sediments, and groundwaters contaminated by SCN− from gold ore processing or coal-coking operations. We quantified the relationships between optimal SCN− biodegradation rates by B. phytofirmans candidate strain ST01hv and pH, phosphate and ammonium concentrations, and the type and concentration of organic carbon substrate, looking for potential metabolic inhibitions or preferences. Results from this study will help to refine bioremediation strategies for treating SCN− contamination in industrial sites.
Footnotes
Acknowledgments
H.V. was supported by a McKenzie Fellowship and Early Career Researcher Grant (University of Melbourne). Dr. Mathew Watts is acknowledged for his helpful comments/suggestions. We thank Mr. David Coe (Kirkland Lake Gold) for access and sampling support.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.