
Abstract
Abstract
Phytoremediation may be used for remediation of cocontaminated soil with heavy metals and polycyclic aromatic hydrocarbons (PAHs). However, the low bioavailability of heavy metals and PAHs in soils may have adverse effects on the efficiency of phytoremediation; it is necessary to find an effective method for simultaneously improving bioavailability of PAHs and heavy metals in cocontaminated soils. In this study, a novel aspartic acid-β-cyclodextrin (ACD) was synthesized, the structural characteristics of ACD were confirmed by FT-IR spectroscopy, and the complexation of fluorene (Flu) and cadmium (Cd) with ACD was studied. Pot culture experiments were conducted to investigate effects of ACD on phytoremediation of soil cocontaminated with Flu and Cd. Results showed that ACD could enhance uptake of Flu and Cd in alfalfa (Medicago sativa L.), and removal percentages of Flu and Cd in planted soil with ACD increased from 45.78% and 25.62% to 66.96% and 34.88%, respectively. Biological activity of planted soils with ACD was obviously higher than that of planted soils without ACD, which resulted in improvement of phytoremediation efficiency. As a promising remediation strategy, ACD-assisted phytoremediation may be a feasible technology for simultaneous removal of PAHs and heavy metals in cocontaminated soils.
Introduction
R
Surfactants can be used to enhance bioavailability of PAHs through micellar solubilization (Gao et al., 2008). Additionally, they may also improve heavy metal bioavailability through the formation of complexes (Mulligan et al., 2001). However, surfactants are difficult to remove due to forming high-viscosity emulsions and have some toxicity effects to soil microorganisms and plants (Ying, 2006).
As a kind of cyclic oligosaccharide consisting of seven glucose units attached by α-1, 4-linkages, β-cyclodextrin (β-CD) has been used successfully to improve solubility and bioavailability of PAHs through the formation of inclusion complexes (Molnar et al., 2002, 2005; Badr et al., 2004). In addition, β-CD offers some advantages over surfactants because of their lower toxicity, higher biodegradability, and low affinity of sorption to the solid phase (Brusseau et al., 1997; Martin, 2004; Gould and Scott, 2005). However, there is no evidence that β-CD may improve the bioavailability of heavy metals. Chemical properties of β-CD such as aqueous solubility and metal complexation potential may be altered by substituting functional groups to the outside of cyclodextrin. Modified β-CD containing carboxyl and amino groups may be used to improve bioavailability of heavy metals through the formation of complexes.
In this work, a novel modified β-CD, aspartic acid-β-cyclodextrin (ACD), was synthesized and characterized. Complexation of Flu and Cd with ACD was studied, the effects of ACD on the accumulation and removal of Flu and Cd in contaminated soil were investigated during the phytoremediation process, and the enhanced removal mechanism of Flu and Cd in contaminated soil was also analyzed. The application of ACD-assisted phytoremediation may provide a kind of feasible technology for remediation of cocontaminated soil caused by heavy metals and PAHs.
Materials and Methods
Materials
β-CD was obtained from Aladdin Reagent Inc. (Shanghai, China), with a purity of 99%. Flu was purchased from Sigma-Aldrich Chemical Co. (Shanghai, China), with a purity of 98%. Minimal salts medium (MSM) was prepared as described previously (Nam et al., 2005, 2008) and contained 3.5 g of Na2HPO4 2H2O, 1 g of KH2PO4, 0.5 g of (NH4)2SO4, 0.1 g of MgCl2 6H2O, 50 mg of Ca(NO3)2 4H2O, and trace elements per liter of distilled water. All the other chemicals used were of analytical grade and purchased from Aladdin Reagent Incorporation (Shanghai, China).
Purified water was obtained using the Milli-Q water purification system (Millipore). Alfalfa seeds (Medicago sativa L.) were purchased from Shanghai Yanhua Bio-Tech Co., Ltd. (Shanghai, China); 1.5% ACD was used for the phytoremediation experiments.
Preparation of ACD
β-CD (8.1 g), potassium hydroxide (6.7 g), and aspartic acid (13.3 g) were dissolved successively in 70 mL of deionized water at 50°C, and then epichlorohydrin (10.2 g) was added dropwise to the above solution. The mixture was then reacted at 60°C for 1 h and cooled to room temperature. The pH was adjusted to 5–6 using sulfuric acid. The mixture was dehydrated by using anhydrous ethanol (150 mL) and pumped through a column packed with neutral alumina. Ethanol (60%, v/v) was used as a flushing agent and was repeatedly passed through the column. The effluent was left to stand for 4–5 h and supernatant was moved away. With the addition of enough anhydrous methanols to the residue, white ACD precipitate was obtained and the precipitate was then filtrated, freeze-dried, and stored. The reaction scheme for the synthesis is shown in Fig. 1.

Reaction scheme for synthesis of ACD. ACD, aspartic acid-β-cyclodextrin.
Soil properties and preparation of cocontaminated soil
Soil without detectable Flu and Cd was collected from the top layer (0–20 cm) of an agricultural field in Fuzhou City, China. The fresh soil was brought to the laboratory, some large particles and roots in the soil were removed by hand, and then the soil samples were air-dried, ground, and sieved through a 2-mm mesh screen; the soil properties were determined according to standard methods (Lu, 2000).
The soil was a fine sandy loam containing about 60% sand, 30% silt, and 10% clay. The physical–chemical properties of soil were given as follows: pH 6.92 (1:2.5, soil/water), organic matter 23.40 g/kg, TN 3.40 g/kg, TP 2.18 g/kg, and CEC 24.36 cmol/kg.
Preparation of cocontaminated soils with Cd and Flu was carried out as follows: first, the Cd-contaminated soil was prepared by completely mixing clean soils with cadmium acetate solution, the levels of Cd added into soil were 5, 15, and 30 mg Cd/kg soil, and the soils were air-dried at room temperature for a week. Second, Flu was initially dissolved in hexane, then three levels of Flu (50, 100, and 200 mg/kg) were added to Cd-contaminated soils, respectively, and nine different treatments may be obtained, including Flu50Cd5, Flu50Cd15, Flu50Cd30, Flu100Cd5, Flu100Cd15, Flu100Cd30, Flu200Cd5, Flu200Cd15, and Flu200Cd30. Flu50, Flu100, and Flu200 denote 50, 100, and 200 mg fluorene/kg soil, respectively. Cd5, Cd15, and Cd30 denote 5, 15, and 30 mg cadmium/kg soil, respectively. The Cd-Flu cocontaminated soils were incubated at room temperature for 30 days in the dark.
Experimental design
One kilogram of the treated soil was placed in each plastic pot (15 cm in height and 18 cm in diameter). Precultured seedlings of alfalfa (10 per pot) with uniform sizes were transplanted to each relevant planting pot. The soil surface was covered with a layer of silica sand to minimize Flu volatilization. ACD was added with ACD crystals at a final concentration of 1.5% (w/w, ACD/soil). Each treatment was prepared in triplicate and the position of pots was changed randomly every week. Soil water content was kept at about 50% of the field water-holding capacity by weighing the pots, regularly adding sterile distilled water as necessary, and a Petri dish was placed under each pot to collect potential leachate during the experiment. The plants were harvested after 90 days of growth, washed with deionized water, and separated into roots, stems, and leaves. After drying with filter paper, the samples were freeze-dried (at 530 ± 17 Pa and −50°C ± 2°C) and weighed to determine the biomass. The soil samples were air-dried at room temperature and ground sufficiently to pass through a 2-mesh sieve.
The removal efficiency (R) of Flu or Cd in contaminated soil was calculated as follows:
where C0 and Ct are the initial concentrations and residual concentrations of Flu or Cd in soil, respectively.
Extraction and analysis of Flu in plants and soils
Extraction of Flu in soils and plants was performed according to Fan et al. (2008). Briefly, 2 g of soil sample was mixed with 20 mL of dichloromethane and extracted for 2 h in an ultrasonic instrument, in which the water temperature was kept below 40°C. As for plant tissues analysis, about 0.5 g of dried plant fragment was weighed into a 25-mL centrifuge tube and extracted by ultrasonication for 30 min with a 30 mL solution of 1:1 hexane and acetone (v/v) for three successive extractions. After each extraction, the extract was centrifuged at 4,000 rpm for 15 min to separate the supernatant from the soil or plant. Then, 2 mL of the supernatant was filtered through 2 g of silica gel column and eluted consecutively with 10 mL of the mixture of 1:1 hexane and dichloromethane (v/v). The extract was concentrated to near dryness in a rotary vacuum evaporator at 40°C, afterward 2 mL of chromatography-grade methanol was added in a rotary steam bottle, mixed, and filtered through a 0.22-μm organic phase membrane and samples were sealed in vials for analysis.
Flu concentrations were determined using high-pressure liquid chromatography (HPLC; Agilent 1260LC) with a 4.6 × 150 mm reverse-phase XDB-C18 column and FLD detector. Chromatography was performed at 30°C with acetonitrile/water (80/20, v/v) as the mobile phase at a flow rate of 1 mL/min. The excitation wavelength of Flu was 275 nm, while the emission wavelength was 350 nm for the FLD detector.
Analysis of Cd
Samples of the plant were digested with a solution of 3:1 HNO3:HClO4 (v/v) in closed Teflon vessels at 443 K, and soil samples were digested with a solution of 10:5:5:3 of HCl:HNO3:HF:HClO4 at 453 K. Cd concentrations in digested samples were determined at 229 nm using an AA 240 Duo atomic absorption spectrophotometer.
Speciation of Cd in soils was analyzed by sequential extraction according to the Tessier method (Tessier et al., 1979). It is divided into five fractions: (1) the exchangeable fraction (F1); (2) the fraction bound to carbonate (F2); (3) the fraction bound to Fe-Mn oxides (F3); (4) the fraction bound to organic matter (F4); and (5) the residual fraction (F5).
Isolation and identification of Flu-degrading bacteria
One gram of soil was added to 100 mL of MSM liquid medium supplemented with Flu and incubated aerobically at 30°C on a reciprocal shaker at 150 rpm. Aliquots were transferred weekly to fresh MSM containing corresponding Flu at a concentration of 100 mg/L as the sole carbon and energy source and incubated under the same conditions. This process was repeated four times until the bacterial strains were isolated. Isolation and purification procedures were carried out on MSM agar plates by conventional spread plate techniques. After about 2 weeks, developed colonies on Flu plates were isolated as bacteria with a potential to degrade Flu (Nam et al., 2015).
To identify the bacteria, genomic DNA was extracted and 16S rDNA was amplified by polymerase chain reaction (PCR) using the following primers: F27(5′-CAGCGGTACCAGAGTTT GATCCTGGCTCAG-3′); R1492(5′-CTCTCTGCAGTACGGCTACCTT GTTACGACTT-3′) (Heuer et al., 1997). The amplification was carried out in GeneAMP®PCR system 9700 by using a temperature profile as follows: predenaturation at 95°C for 5 min, denaturation at 94°C for 30 s, annealing at 53°C for 30 s, extension at 72°C for 90 s (35 cycles), and a final extension at 72°C for 10 min. The amount of recovered DNA was estimated by agarose gel electrophoresis. The PCR product was purified with a TIANgel Midi Purification Kit (Tiangen, Beijing, PR China) and sequencing was done by BGI Company (Shenzhen, PR China). The obtained sequences were compared with the GenBank database using the Basic Local Alignment Search Tool (BLAST). The calculation of sequence similarity and phylogenetic tree inference were carried out using the MEGA4 program (Masakorala et al., 2013).
Determination of microbes' growth curve and degradation of Flu by strains
To investigate the growth and biodegrading ability of the strain, the bacterial suspension (cell concentration: ∼107 colony-forming units [CFU]/mL) was inoculated with sterile selective liquid medium (volume percentage is 10%) with an initial concentration of Flu of 40 mg/L and at pH 7.0. All flasks with the medium were incubated on a rotary shaker (35°C, 120 rpm) in the dark. The growth of bacteria was measured every day by determining the optical density (OD) of the culture solution using the visible spectrophotometer at wavelength 600 nm, and the residual concentration of Flu was determined by HPLC.
Determination of root colonization
Root colonization by arbuscular mycorrhizae (AM) was determined according to literatures (Huang et al., 2007; Wu et al., 2008). One gram of fresh roots was randomly taken and cut into ∼1-cm pieces to estimate the proportion of total root length colonized by AM. Root segments were immersed in 10% (v/v) KOH for 10 min at 90°C in a water bath, rinsed with water, and then stained with 0.1% Trypan blue for 3–5 min at 90°C in a water bath. Mycorrhizal colonization was determined by the grid line intersect method (Phillips and Hayman, 1970).
Bacterial counts and polyphenol oxidase activity
Soil Flu-degrading bacteria were enumerated by using the plate spread method over MSM agar amended with 0.05 g/L Flu and 75 mg/L cycloheximide for suppression of fungal growth. A 10-fold serial dilution factor (from 10−1 to 10−8 dilution) was prepared for each retrieved sample. An aliquot of 0.1 mL diluted culture was spread over agar plates, and each dilution had three replicates. The number was counted after 14 days of incubation at 30°C, and results were expressed as CFU/g soil (Chen et al., 2016).
Polyphenol oxidase (PPO) activity in soil was determined as described by Yu et al. (2011). The mixture of 1.0 g of soil and 10 mL of 1% pyrogallic acid was incubated at 30° for 2 h, and 4 mL of citric–phosphoric acid buffer (pH 4.5) was added to the mixture. The purpurogallin (PPG) formed was extracted with chloroform and then measured spectrophotometrically at 430 nm. The results were expressed as μg PPG/g soil.
Quality control
Experiments on Flu recovery were carried out by adding a known concentration of Flu standards (1 mg/kg) to the uncontaminated soil and plant. The recovery efficiency was 95.3% ± 5.4% and 97.6% ± 8.7% for soils and plants, respectively. Similarly for Cd, a certified reference material, bush leaf material (GBW07603, China), was used to verify the accuracy and precision of the digestion procedure and subsequent analysis.
Statistical analysis
In this experiment, all data are represented as mean ± standard deviation of three replicates. Statistical analysis of the results was performed with SPSS 11.0 for Windows (SPSS, Inc.). Means were compared using a t-test. The least significant difference of means at a 5% confidence level (p < 0.05) was applied to assess significant differences.
Results and Discussion
Characterization of ACD
FT-IR spectra of aspartic acid, β-CD, and ACD are shown together in Fig. 2. Compared with the FT-IR of β-CD, the FT-IR spectra are similar. Some characteristic absorption peaks of β-CD at 857, 946, 1,026, and 1,157 cm−1 have been found in FT-IR spectra of ACD, which indicates that the cavity structure of β-CD was kept in ACD molecular. In addition, characteristic absorption peaks at 1,742 and 1,400 cm−1 for ACD could be ascribed to C = O and H–O stretching vibration in carboxyls, and the peaks at 1,504 and 1,342 cm−1 are assigned to N–H deformation vibration and C = N stretching vibration, respectively. These results show that the synthetic compound (ACD) has a cavity structure of CD; in addition, the carboxyl and amino groups occur outside the cavity of CD.
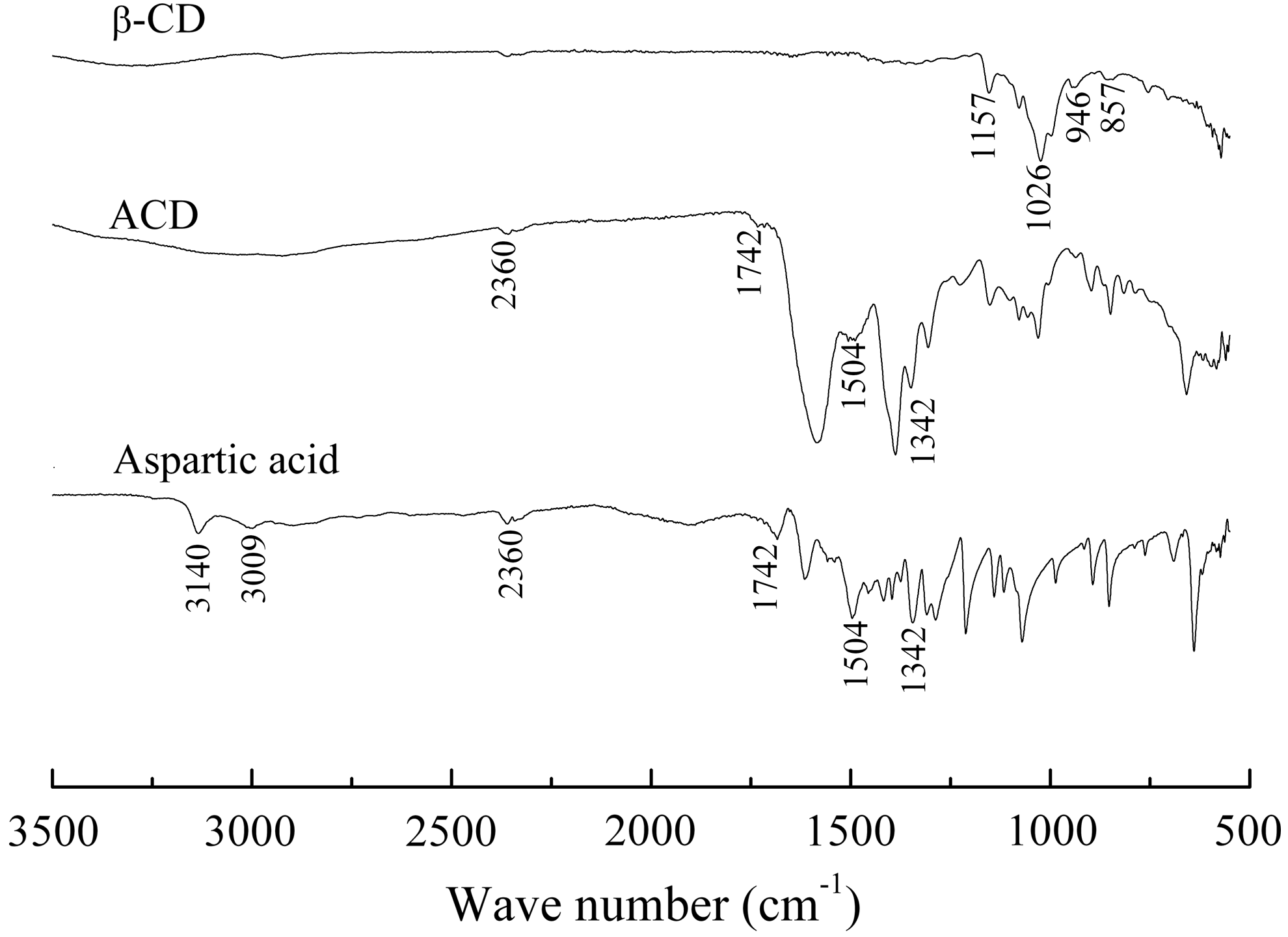
FT-IR spectra of β-CD and ACD. β-CD, β-cyclodextrin.
Characterization of complexation of Flu and Cd with ACD
In this experiment, the inclusion interaction between ACD and Flu was characterized through fluorescence spectra, and results are shown in Fig. 3. The fluorescence intensity of the inclusion complex was enhanced with increasing ACD concentration. This is because as Flu molecules enter the hydrophobic inner cavity of ACD, the revolving freedom of Flu decreases, resulting in an increase in fluorescence quantum yield of Flu molecules (Raymond et al., 1991).

Fluorescence spectra of Flu/ACD inclusion complex. [Flu] = 1.0 mg/L; pH 6.0. Flu, fluorene.
Complexation interaction between ACD and Cd was characterized using XPS spectra. Table 1 shows the XPS data of N1s, O1s, and Cd 3d in ACD and ACD-Cd coordination complex. In comparison with the Cd 3d electron in CdCO3, the binding energy of the Cd 3d electron in ACD-Cd decreased by 0.80 eV. However, binding energies of the O1s and N1s electrons of ACD-Cd increased by 0.88 and 0.60 eV, compared with those of the O1s and N1s electrons of ACD. It is because N and O atoms in ACD are believed to be electron-pair donors, leading to a decrease in electron density of N and O atoms in the ACD-Cd complex, and Cd (II) is an electron-pair acceptor, leading to an increase in the electron density of Cd in the ACD-Cd complex. Therefore, these spectral changes indicate the formation of an ACD-Cd complex. The carboxyl and amino groups of ACD are likely to be responsible for Cd binding and these groups complex with Cd outside the cavity.
ACD, aspartic acid-β-cyclodextrin.
Identification of strain W-1
The 16S rDNA gene sequence of strain W-1 was analyzed and compared with other sequences published in the GenBank database through BLAST on the National Center of Biotechnology Information (NCBI) website. The BLAST result showed that W-1 had high sequence similarity to Alcaligenes faecalis strain cb-4 (100% homology). This sequence was deposited in the NCBI GenBank (accession No. FJ588233).
Growth of strain W-1 and degradation of Flu
Growth of strain W-1 and degradation of Flu are shown in Fig. 4. The results indicated that growth of strain W-1 and degradation of Flu were synchronous and presented a mirror-image relationship. In the beginning of the cultivation, the growth of strain W-1 was in the lag phase, cells grew slowly, and the Flu removal rate was inefficient. After a transitory lag period of about 3.5 days, cells turned into the logarithmic phase. In this stage, the strain used Flu as the sole carbon and energy source for breeding and significant growth. Flu in the medium was metabolized gradually to lead to the drop of Flu concentration. Cells turned into the stationary phase after this stage and the OD at 600 nm of culture reached a maximum of 1.15. At this moment (namely the 11th day), the concentration of Flu in the media decreased from 40 to 5.0 mg/L and the biodegradation rate reached 87.5%. The isolated strains had a high growth and degradation rate in the culture system. This indicated that W-1 was a predominant Flu-degrading strain with high activity.
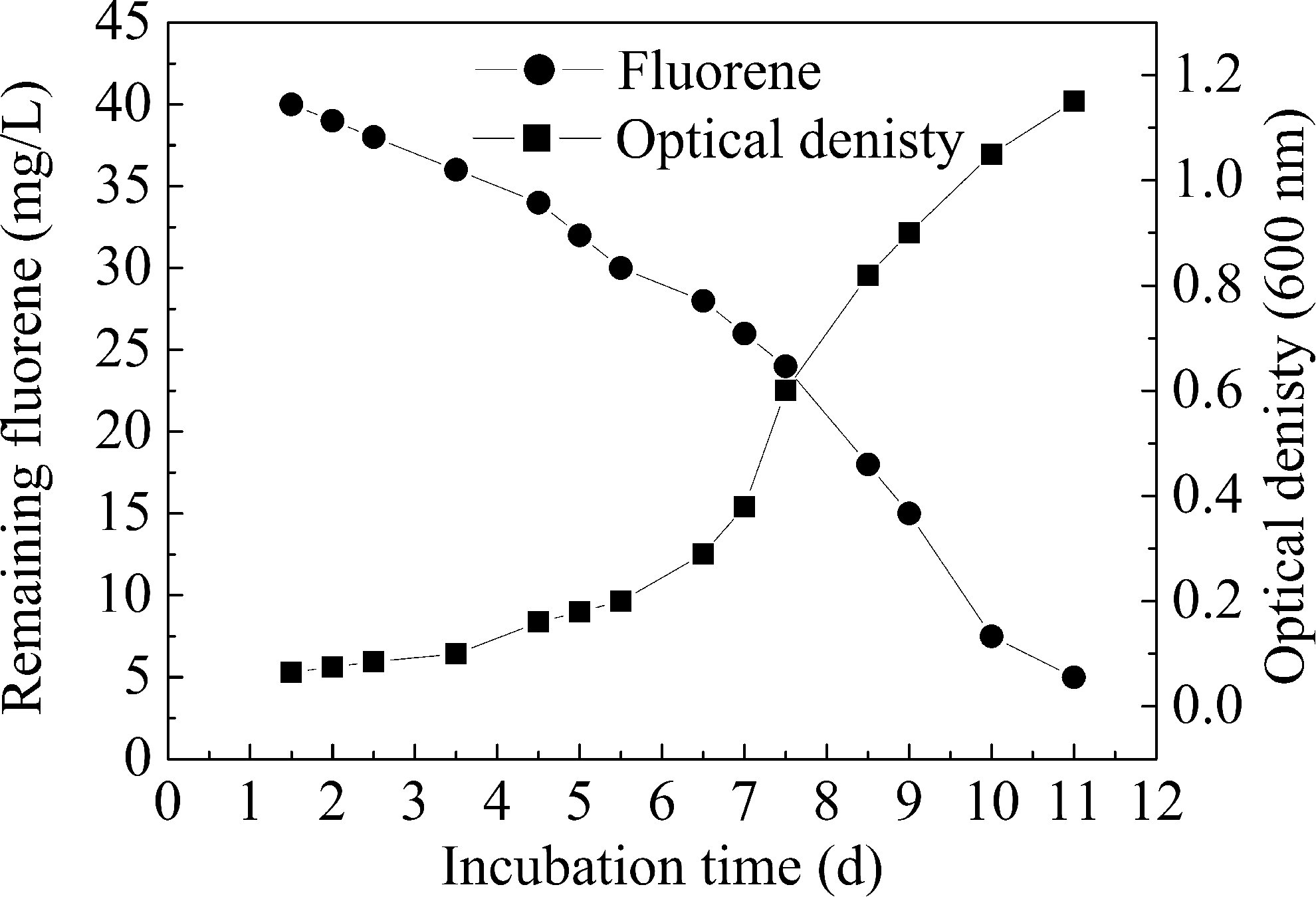
Growth curve of strain W-1 and degradation of Flu.
Symbiotic status of alfalfa with indigenous arbuscular mycorrhizal fungi and rhizobia
To investigate the symbiotic status of alfalfa with indigenous arbuscular mycorrhizal fungi (AMF) and rhizobia in soil, the mycorrhizal colonization rate and root nodule number per plant were determined, respectively; results are shown in Table 2. The results showed that there were significant differences (p < 0.05) in the mycorrhizal colonization rate and root nodule number per plant for different treatments. Alfalfa could form a better dual symbiosis with indigenous rhizobia and AMF and had no restraining effect between nodule and AM formation. The mycorrhizal colonization rate of alfalfa roots for uncontaminated soils is obviously higher than that of alfalfa roots for contaminated soils, and the higher root nodule number per plant is also obtained in uncontaminated soils. In cocontaminated soils, the mycorrhizal colonization rate and root nodule number decrease with increasing concentration of Cd, and Cd has obvious inhibition effects on AMF and rhizobia. However, the mycorrhizal colonization rate and root nodule number in the presence of ACD are obviously higher than those in the absence of ACD. It is because ACD can decrease toxicity of Cd through formation of complexes. Thus, the presence of ACD in soil is beneficial to the growth of indigenous AMF and rhizobia.
Values in each column followed by different capital letters (A and B) indicate significant (p ≤ 0.05) differences between soil amended with and without ACD and values in each row followed by different lowercase letters (a–d) indicate significant (p ≤ 0.05) differences between different treatments. Values represent mean ± SD.
SD, standard deviation.
Effects of ACD on biomass, microbial numbers, and PPO activity
To evaluate the effect of ACD on shoot and root growth of alfalfa (M. sativa L.), different concentrations of ACD were added to uncontaminated soil; effects of ACD on biomass of alfalfa shoots and roots are shown in Fig. 5. The results show that ACD had a stimulatory effect on shoot and root biomass yields, biomass yields of shoots and roots of alfalfa significantly increased with increasing ACD concentration, and higher biomass of alfalfa shoots and roots was obtained in the presence of 1.5% ACD. It is because ACD can improve the permeability of the cell membrane, which results in a more efficient uptake of nutrients in soil (Mohamed et al., 2002). In addition, alfalfa could form a better dual symbiosis with indigenous AMF and rhizobia; AMF can improve plant growth and health by improving mineral and water nutrition and by increasing tolerance to biotic and abiotic stresses, and rhizobia are bacteria that fix nitrogen inside the root and stem nodules that they incite on leguminous plants. Thus, the presence of AMF and rhizobia in soil is beneficial to the host plant's growth (Wang et al., 2011); when ACD is added to the soil, ACD can promote the growth of AMF and rhizobia, which can further improve the nutritional status of host plants, promoting their growth and development (Rajtor and Piotrowska-Seget, 2016). Thus, the presence of ACD is beneficial to the growth of alfalfa during the phytoremediation process.

Effect of ACD on biomass of shoots and roots of alfalfa. Columns denoted by different capital letters indicate a significant difference between the shoot and root. Columns denoted by different lowercase letters indicate a significant difference between different treatments.
When the concentration of Cd and Flu in contaminated soil is 5 and 50 mg/kg, respectively, the effects of ACD and plant on the numbers of Flu-degrading bacteria in soil are recorded and shown in Table 3. It was clear that numbers of Flu-degrading bacteria in soil planted with alfalfa were significantly higher than those in the unplanted soil. The rhizosphere effects could explain differences between planted soils and unplanted soils, and the roots are known to release some organic compounds, such as amino acids, organic acids, sugars, and carbohydrates, providing a carbon source and energy for the growth of microorganisms (Shim et al., 2000; Singer et al., 2003). The results also showed that the application of ACD could significantly increase microbial numbers for both planted soils and unplanted soils, and the numbers of Flu-degrading bacteria in the planted soils with ACD were 1.36 times that of Flu-degrading bacteria in the planted soils without ACD. The bioavailability of Flu in soil could be enhanced in the presence of ACD, and ACD could also provide carbon, nitrogen, and energy sources for microbial populations, which resulted in increase of microbial population numbers. Thus, the presence of ACD is beneficial for biodegradation of Flu in soil.
Values in each row followed by different lowercase letters (a–d) indicate significant (p ≤ 0.05) differences between different treatments. Values represent mean ± SD.
CFU, colony-forming units; PPG, purpurogallin; PPO, phenol oxidase.
As one of the most important oxidoreductases in soil, PPO can be used for conversion of aromatic organic compounds in humus (Chen et al., 2016). The effects of the plant and ACD on PPO activity in soil were also investigated, and results are shown in Table 2. It was shown that PPO activity in the soil planted with alfalfa was much higher than that in unplanted soils, alfalfa growth enhanced PPO activity, and ACD can increase PPO activity in the soil. Furthermore, PPO activity in the planted soil with ACD was 1.30 times higher than that in planted soils without ACD. PPO activity results matched well with results of microbial counts and biomass in the planted soil. It could be explained that the PPO activity was enhanced due to the supply of nutrients from vigorous plant roots and the flourished growth of microbes with the addition of ACD (Shen et al., 2009).
Effects of ACD on accumulation and removal of Flu
To investigate the effects of ACD on the plant uptake of Flu, the concentrations of Flu in alfalfa in the presence and in the absence of ACD were determined, and results are shown in Fig. 6. The uptake of Flu by alfalfa obviously increased with increasing concentrations of Flu in contaminated soil, while it decreased with increasing concentrations of Cd in contaminated soil. The concentrations of Flu in alfalfa with ACD were 1.1–1.3 times those of Flu in alfalfa without ACD, which indicates that the application of ACD can enhance the uptake of Flu from contaminated soil by alfalfa. It is because ACD may improve the bioavailability of Flu through the formation of inclusion complexes.
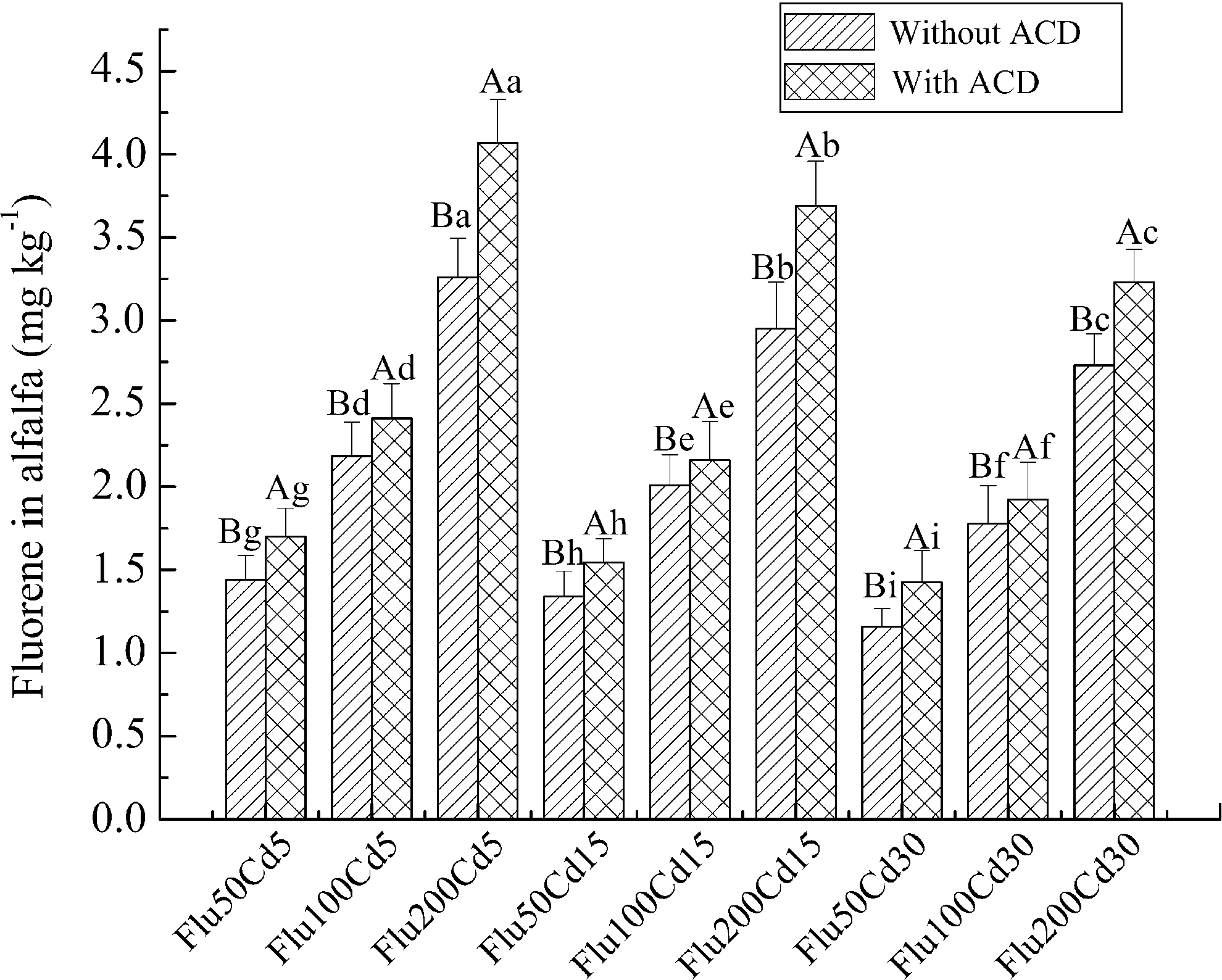
Flu concentration in alfalfa with and without ACD. Columns denoted by different capital letters indicate a significant difference between soil amended with and without ACD. Columns denoted by different lowercase letters indicate a significant difference between different treatments.
When the concentration of Cd and Flu from cocontaminated soil is 5 and 50 mg/kg, respectively, after 90 days of greenhouse pot experiment, the removal percentage of Flu in contaminated soil is recorded and shown in Table 4. The results show that the removal percentage of Flu in the soils planted with alfalfa was significantly higher than that of Flu in the unplanted soils. It is because the rhizosphere effects occurred in the planted soil with alfalfa and the bioavailability of Flu was increased, which results in cometabolic degradation of Flu in contaminated soil (Singer et al., 2003). A more recent study revealed that AMF hyphae could facilitate the absorption of PAHs by the host plant (Yu et al., 2011). Furthermore, the Flu removal percentage was 66.96% in the planted soil with ACD, while it was 45.78% in the planted soil and 17.24% in the unplanted soil for no amendment. The enhanced removal of Flu in the planted soil with ACD mainly results from enhanced accumulation and degradation of Flu in the presence of ACD. Previous results have proven that the presence of ACD has a stimulatory effect on microbial numbers and PPO activity in soil, and the greater activity of microorganisms and PPO results in enhanced degradation of Flu in planted soil with ACD. In addition, ACD may also improve the AMF colonization rate in the planted soil, which is beneficial for the uptake of Flu in the plant.
Values in each row followed by different lowercase letters (a–d) indicate significant (p ≤ 0.05) differences between different treatments. Values represent mean ± SD.
Effects of ACD on accumulation and removal of Cd
To evaluate the effects of ACD on the plant uptake of Cd in contaminated soil, concentrations of Cd in alfalfa in the presence and in the absence of ACD were determined, and results are shown in Fig. 7. The uptake of Cd by alfalfa obviously increased with increasing concentration of Cd in contaminated soil, and the presence of Flu had a negligible effect on the plant uptake of Cd in soil. Results are similar to those of Zhang et al. (2012). The concentrations of Cd in alfalfa with ACD were 1.1–1.2 times those of Cd in alfalfa without ACD, which indicates that the application of ACD can enhance the uptake of Cd from contaminated soil by alfalfa. It is because ACD may improve the bioavailability of Cd in contaminated soil through the formation of coordination complexes.

Cd concentration in alfalfa with and without ACD. Columns denoted by different capital letters indicate a significant difference between soil amended with and without ACD. Columns denoted by different lowercase letters indicate a significant difference between different treatments. Cd, cadmium.
When the concentration of Cd and Flu from cocontaminated soil is 5 and 50 mg/kg, respectively, after 90 days of greenhouse pot experiment, the removal percentage of Cd in soil is recorded and shown in Table 4. The results show that the removal percentage of Cd in soils planted with alfalfa was obviously higher than that in unplanted soils. The removal efficiency of Cd was increased due to rhizosphere effects in the planted soils. Some beneficial bacteria and fungi, acting as plant growth-promoting rhizobacteria (PGPR), may alleviate metal phytotoxicity and stimulate plant growth indirectly, and microorganisms can also change metal bioavailability in soil through various mechanisms such as acidification, chelation, complexation, and redox reactions. In the presence of ACD, PGPR activity in planted soil can be further improved. It was suggested that ACD could increase the permeability of the cell membrane, which could lead to a more efficient uptake of nutrients for rhizobacteria. Furthermore, in the presence of ACD, the removal percentage of Cd in the planted soil increased from 25.62% to 34.88% because of enhanced accumulation effects. Thus, the presence of ACD is also beneficial for the removal of Cd in planted soil.
Enhanced removal mechanism of Flu and Cd in contaminated soil
During the phytoremediation process, the removal of PAHs in contaminated soil was mainly attributed to plant metabolism and accumulation, microbial degradation, and plant–microbial interactions. Plant–microbial interaction is the predominant pathway responsible for the removal of PAHs (Wei and Pan, 2010). Thus, the removal efficiency of PAHs in contaminated soil mainly depends on the bioavailability of PAHs in contaminated soil; however, the bioavailability of PAHs was relative to the solubility of PAHs. In these experiments, the effects of ACD on the solubility of Flu were investigated. The solubilization effects of β-CD and ACD on Flu are plotted in Fig. 8. The results show that the apparent aqueous solubilities of Flu are linearly increased with increasing β-CD and ACD concentrations. This phenomenon is attributed to the formation of 1:l inclusion complexes (Ko et al., 1999). The linear relationship can be expressed as follows:
where St is the aqueous-phase concentration of Flu in the presence of CD, S0 is the concentration of Flu in the absence of CD, C0 represents the initial concentration of CD, and Kf is the stability constant of inclusion complexes for Flu with CD, and it was used to evaluate the solubilization capacity of CD for phenanthrene. As shown in Fig. 8, the stability constant of inclusion complex for Flu with β-CD and ACD is 1.663 and 4.716, respectively, and a higher binding constant was found for Flu with ACD than with β-CD, which indicates that the solubilization capacity of ACD for Flu is higher than that of β-CD for Flu. ACD has obvious solubilization for Flu; the solubility of Flu in 10 g/L of ACD was enhanced about 49-fold. Therefore, the presence of ACD may obviously improve the solubility and bioavailability of Flu in contaminated soil. In addition, the presence of ACD also stimulated biological activities (microbial assays and PPO activities), leading to enhanced Flu degradation. Thus, the presence of ACD is beneficial to the bioremoval of Flu in contaminated soil.
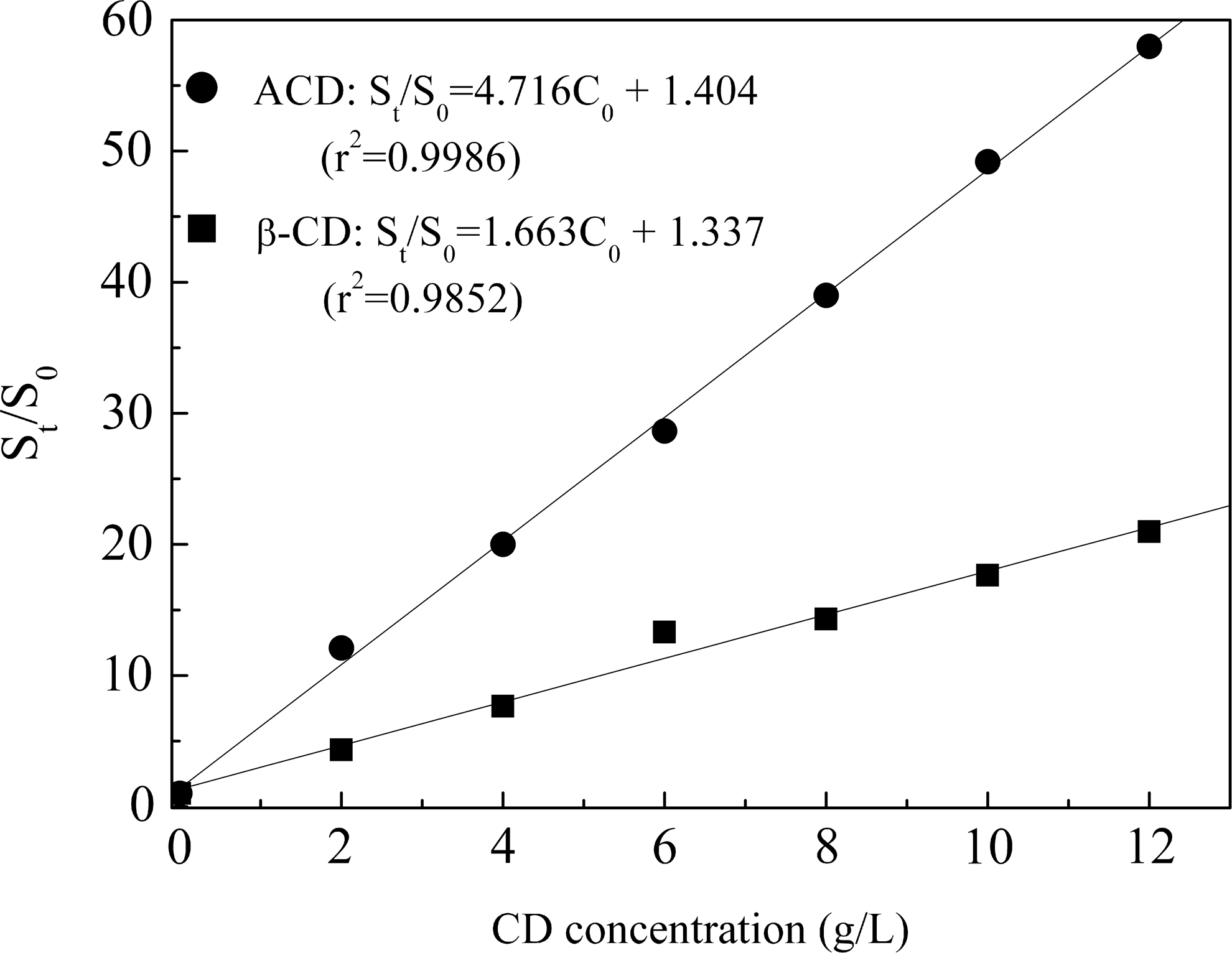
Solubilization curves of Flu by β-CD and ACD.
During the phytoremediation process, the removal of heavy metals in contaminated soil mainly derived from plant accumulation, and heavy metal concentrations in plants largely depend on heavy metal bioavailability in soil (Abou-Shanab et al., 2006). However, the bioavailability of heavy metal was relative to the metal speciation in soil. In general, the solubility and bioavailability of heavy metals decrease in the following order: exchangeable fraction > carbonate-bound fraction > Fe-Mn oxide-bound fraction > organic-bound fraction > residual fraction (Mahanta and Bhattacharyya, 2011). Effects of ACD on the speciation distribution and transformation of Cd in soils are shown in Fig. 9; the addition of ACD altered the speciation distribution of Cd in soils and the exchangeable fraction obviously increased; however, the carbonate-bound fraction, Fe/Mn oxide-bound fraction, and organic fraction obviously decreased. The results suggested that added ACD enhanced the solubility of Cd in soils and resulted in high bioavailability of Cd in soil. Thus, the presence of ACD can also enhance the bioremoval of Cd in contaminated soil.

Effects of ACD on the speciation distribution of Cd in soils. F1, F2, F3, F4, and F5 denote the exchangeable fraction, carbonate fraction, Fe–Mn oxide fraction, organic fraction, and residual fraction, respectively.
Conclusions
Addition of ACD increased the bioavailability of Flu and Cd in soil, and the concentrations of Flu and Cd in alfalfa significantly increased in the presence of ACD; ACD can enhance the accumulation of Flu and Cd in alfalfa. In addition, ACD also stimulated biological activities (microbial assays and PPO activities), leading to enhanced Flu degradation. ACD-assisted phytoremediation technology was an effective and environmentally friendly remediation means for the removal of PAHs and heavy metals from cocontaminated soils.
Footnotes
Acknowledgments
This work was financed by the Natural Science Foundation of China (41761069, 41261078), the Natural Science Foundation of Jiangxi, China (20142BAB213021), and the Science Funds of the Education Office of Jiangxi, China (GJJ12374). The authors thank the anonymous reviewers for their comments.
Author Disclosure Statement
No competing financial interests exist.